Immune Evasion Strategies of Ranaviruses and Innate Immune Responses to These Emerging Pathogens


{kind=link}
{kind=link}
{kind=link}
Abstract
:Abbreviations
| [ATV] | Ambystoma tiginum virus |
| [BIV] | Bohle Iridovirus |
| [CARD] | caspase activation and recruitment domain |
| [CCV] | channel catfish herpes virus |
| [DE] | delayed early genes |
| [EHNV] | epizootic haematopoietic necrosis virus |
| [eIF2α] | eukaryotic translation initiation factor 2 alpha |
| [FV3] | frog virus 3 |
| [HIV] | human immunodeficiency virus |
| [IE] | immediate early genes |
| [IFNγ] | interferon gamma |
| [IL-1β] | interleukin-1 beta |
| [IRF] | interferon regulatory factor |
| [L] | late genes |
| [MAPK] | mitogen activated protein kinase |
| [MX1] | Myxovirus-resistance1 |
| [ORF] | open reading frame |
| [PKR] | RNA-dependent protein kinase |
| [PL] | peritoneal leukocyte |
| [RCV-Z] | Rana (Lithobates) catesbeiana Virus Z |
| [RV] | ranavirus |
| [SGIV] | Singapore Grouper Iridovirus |
| [TGIV] | Taiwan Grouper Iridovirus |
| [TNFα] | tumor necrosis factor alpha |
| [vIFα] | viral translation initiation factor-alpha homolog |
1. Introduction
2. The Role of Myeloid Cells in Ranaviral Infections
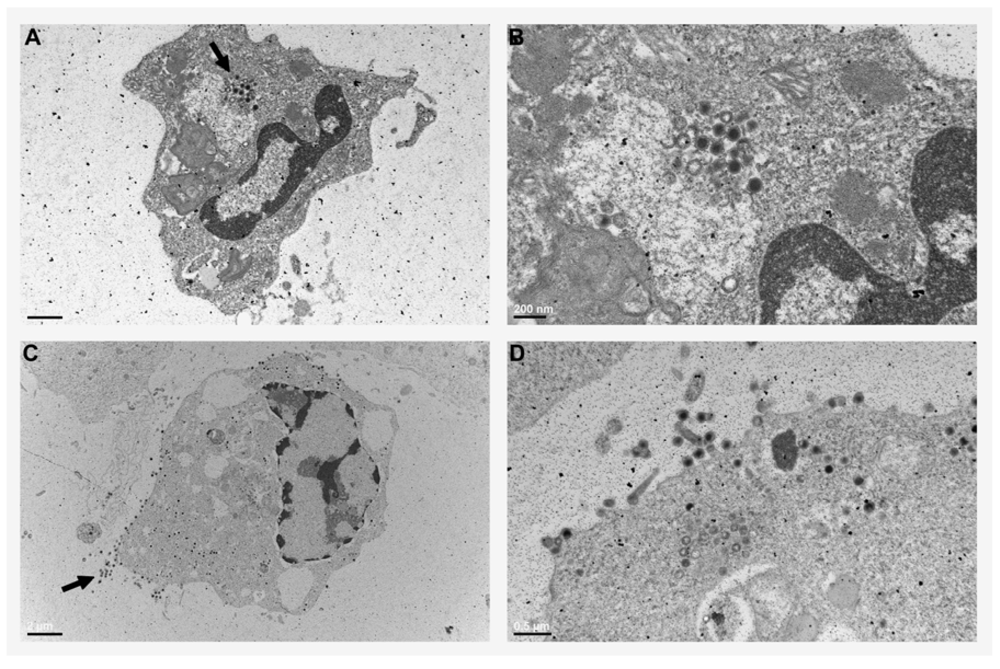
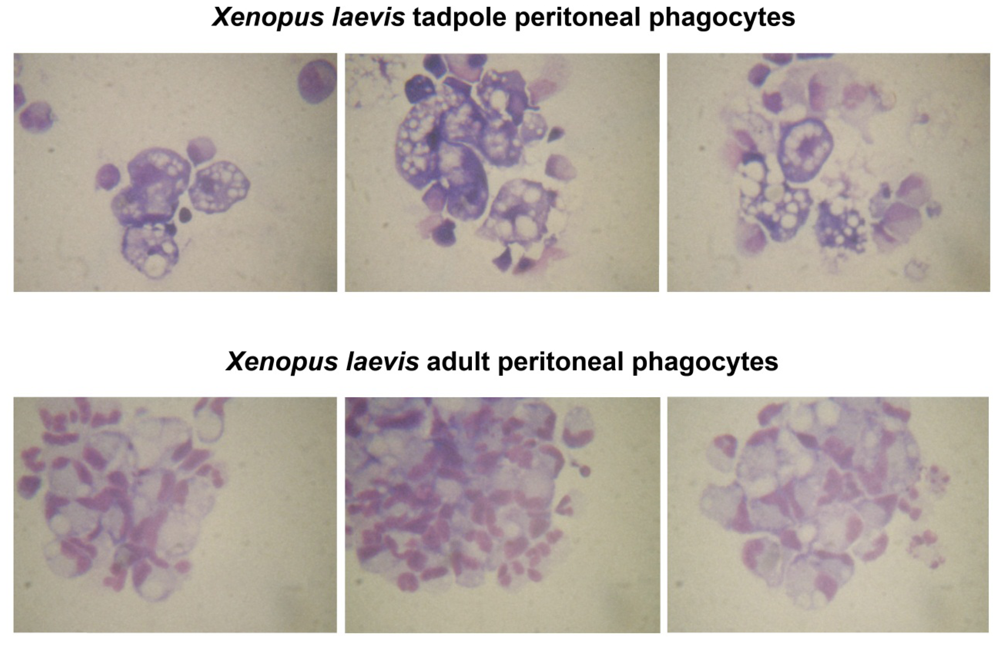
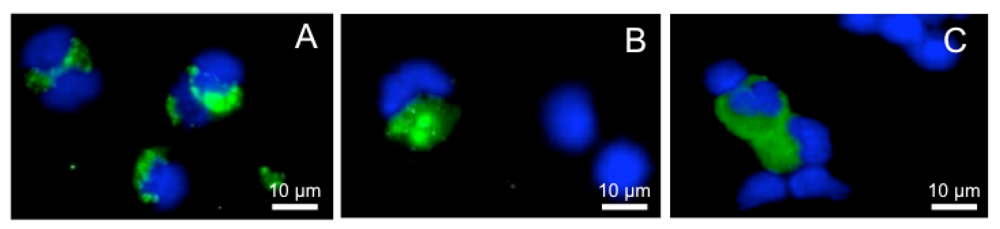
2.2. Xenopus Macrophage Model of Ranaviral Infections

2.3. Macrophage Involvement in Iridovirid Infections of Other Poikilotherms
3. Innate Immune Responses to Ranaviral Infections
3.2. Pro-inflammatory Responses to Iridovirid Infections
3.2.1. Cell Signaling Pathways Induced by Iridovirids
3.2.2. Xenopus Tadpole and Adult Inflammatory Responses to Ranavirus Infections


5. Concluding Remarks
Conflict of Interest
Acknowledgements
References
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar]
- Collins, J.P. Amphibian decline and extinction: What we know and what we need to learn. Dis. Aquat. Organ. 2010, 92, 93–99. [Google Scholar] [CrossRef]
- Daszak, P.; Berger, L.; Cunningham, A.A.; Hyatt, A.D.; Green, D.E.; Speare, R. Emerging infectious diseases and amphibian population declines. Emerg. Infect. Dis. 1999, 5, 735–748. [Google Scholar] [CrossRef]
- Green, D.E.; Converse, K.A.; Schrader, A.K. Epizootiology of sixty-four amphibian morbidity and mortality events in the USA, 1996–2001. Ann. N. Y. Acad. Sci. 2002, 969, 323–339. [Google Scholar] [CrossRef]
- Gray, M.J.; Miller, D.L.; Hoverman, J.T. Ecology and pathology of amphibian ranaviruses. Dis. Aquat. Organ. 2009, 87, 243–266. [Google Scholar] [CrossRef]
- Jancovich, J.K.; Jacobs, B.L. Innate immune evasion mediated by the Ambystoma tigrinum virus eukaryotic translation initiation factor 2alpha homologue. J. Virol. 2011, 85, 5061–5069. [Google Scholar] [CrossRef]
- Chinchar, V.G.; Hyatt, A.; Miyazaki, T.; Williams, T. Family iridoviridae: Poor viral relations no longer. Curr. Top. Microbiol. Immunol. 2009, 328, 123–170. [Google Scholar]
- Robert, J.; Ohta, Y. Comparative and developmental study of the immune system in Xenopus. Dev. Dyn. 2009, 238, 1249–1270. [Google Scholar] [CrossRef]
- Gordon, S.; Martinez, F.O. Alternative activation of macrophages: Mechanism and functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef]
- Coiras, M.; Lopez-Huertas, M.R.; Perez-Olmeda, M.; Alcami, J. Understanding HIV-1 latency provides clues for the eradication of long-term reservoirs. Nat. Rev. Microbiol. 2009, 7, 798–812. [Google Scholar] [CrossRef]
- Gousset, K.; Ablan, S.D.; Coren, L.V.; Ono, A.; Soheilian, F.; Nagashima, K.; Ott, D.E.; Freed, E.O. Real-time visualization of HIV-1 GAG trafficking in infected macrophages. PLoS Pathog. 2008, 4. [Google Scholar]
- Groot, F.; Welsch, S.; Sattentau, Q.J. Efficient HIV-1 transmission from macrophages to T cells across transient virological synapses. Blood 2008, 111, 4660–4663. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Antiviral signaling through pattern recognition receptors. J. Biochem. 2007, 141, 137–145. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. SnapShot: Pattern-recognition receptors. Cell 2007, 129. [Google Scholar]
- Thompson, A.J.; Locarnini, S.A. Toll-like receptors, RIG-I-like RNA helicases and the antiviral innate immune response. Immunol. Cell. Biol. 2007, 85, 435–445. [Google Scholar] [CrossRef]
- Gut, J.P.; Anton, M.; Bingen, A.; Vetter, J.M.; Kirn, A. Frog virus 3 induces a fatal hepatitis in rats. Lab. Invest. 1981, 45, 218–228. [Google Scholar]
- Kirn, A.; Bingen, A.; Steffan, A.M.; Wild, M.T.; Keller, F.; Cinqualbre, J. Endocytic capacities of Kupffer cells isolated from the human adult liver. Hepatology 1982, 2, 216–222. [Google Scholar]
- Kirn, A.; Steffan, A.M.; Bingen, A. Inhibition of erythrophagocytosis by cultured rat Kupffer cells infected with frog virus 3. J. Reticuloendothel. Soc. 1980, 28, 381–388. [Google Scholar]
- Hagmann, W.; Steffan, A.M.; Kirn, A.; Keppler, D. Leukotrienes as mediators in frog virus 3-induced hepatitis in rats. Hepatology 1987, 7, 732–736. [Google Scholar] [CrossRef]
- Aubertin, A.M.; Hirth, C.; Travo, C.; Nonnenmacher, H.; Kirn, A. Preparation and properties of an inhibitory extract from frog virus 3 particles. J. Virol. 1973, 11, 694–701. [Google Scholar]
- Gendrault, J.L.; Steffan, A.M.; Bingen, A.; Kirn, A. Penetration and uncoating of frog virus 3 (FV3) in cultured rat Kupffer cells. Virology 1981, 112, 375–384. [Google Scholar] [CrossRef]
- Elharrar, M.; Hirth, C.; Blanc, J.; Kirn, A. Pathogenesis of the toxic hepatitis of mice provoked by FV3 (frog virus 3): Inhibition of the liver macromolecular synthesis. Biochem. Biophys. Acta. 1973, 319, 91–102. [Google Scholar]
- Kirn, A.; Gut, J.P.; Elharrar, M. FV3 (Frog Virus 3) toxicity for the mouse. Nouv. Presse. Med. 1972, 1, 19–43. [Google Scholar]
- Robert, J.; Abramowitz, L.; Gantress, J.; Morales, H.D. Xenopus laevis: A possible vector of Ranavirus infection? J. Wildl. Dis. 2007, 43, 645–652. [Google Scholar]
- Morales, H.D.; Abramowitz, L.; Gertz, J.; Sowa, J.; Vogel, A.; Robert, J. Innate immune responses and permissiveness to ranavirus infection of peritoneal leukocytes in the frog Xenopus laevis. J. Virol. 2010, 84, 4912–4922. [Google Scholar] [CrossRef]
- Siwicki, A.K.; Pozet, F.; Morand, M.; Volatier, C.; Terech-Majewska, E. Effects of iridovirus-like agent on the cell-mediated immunity in sheatfish (Silurus glanis)—An in vitro study. Virus. Res. 1999, 63, 115–119. [Google Scholar] [CrossRef]
- Chao, C.B.; Chen, C.Y.; Lai, Y.Y.; Lin, C.S.; Huang, H.T. Histological, ultrastructural, and in situ hybridization study on enlarged cells in grouper Epinephelus hybrids infected by grouper iridovirus in Taiwan (TGIV). Dis. Aquat. Organ. 2004, 58, 127–142. [Google Scholar] [CrossRef]
- Chinchar, V.G.; Wang, J.; Murti, G.; Carey, C.; Rollins-Smith, L. Inactivation of frog virus 3 and channel catfish virus by esculentin-2P and ranatuerin-2P, two antimicrobial peptides isolated from frog skin. Virology 2001, 288, 351–357. [Google Scholar] [CrossRef]
- Huang, X.; Huang, Y.; Ouyang, Z.; Cai, J.; Yan, Y.; Qin, Q. Roles of stress-activated protein kinases in the replication of Singapore grouper iridovirus and regulation of the inflammatory responses in grouper cells. J. Gen. Virol. 2010, 92, 1292–1301. [Google Scholar]
- Maniero, G.D.; Morales, H.; Gantress, J.; Robert, J. Generation of a long-lasting, protective, and neutralizing antibody response to the ranavirus FV3 by the frog Xenopus. Dev. Comp. Immunol. 2006, 30, 649–657. [Google Scholar] [CrossRef]
- Gantress, J.; Maniero, G.D.; Cohen, N.; Robert, J. Development and characterization of a model system to study amphibian immune responses to iridoviruses. Virology 2003, 311, 254–262. [Google Scholar] [CrossRef]
- Morales, H.D.; Robert, J. Characterization of primary and memory CD8 T-cell responses against ranavirus (FV3) in Xenopus laevis. J. Virol. 2007, 81, 2240–2248. [Google Scholar] [CrossRef]
- Cuthbertson, R.A.; Lang, R.A.; Coghlan, J.P. Macrophage products IL-1 alpha, TNF alpha and bFGF may mediate multiple cytopathic effects in the developing eyes of GM-CSF transgenic mice. Exp. Eye Res. 1990, 51, 335–344. [Google Scholar] [CrossRef]
- Ferreri, N.R.; Millet, I.; Paliwal, V.; Herzog, W.; Solomon, D.; Ramabhadran, R.; Askenase, P.W. Induction of macrophage TNF alpha, IL-1, IL-6, and PGE2 production by DTH-initiating factors. Cell. Immunol. 1991, 137, 389–405. [Google Scholar] [CrossRef]
- Itoh, A.; Iizuka, K.; Natori, S. Induction of TNF-like factor by murine macrophage-like cell line J774.1 on treatment with Sarcophaga lectin. FEBS Lett. 1984, 175, 59–62. [Google Scholar] [CrossRef]
- Liew, F.Y.; Li, Y.; Millott, S. Tumour necrosis factor (TNF-alpha) in leishmaniasis. II. TNF-alpha-induced macrophage leishmanicidal activity is mediated by nitric oxide from L-arginine. Immunology 1990, 71, 556–559. [Google Scholar]
- McMasters, K.M.; Cheadle, W.G. Regulation of macrophage TNF alpha, IL-1 beta, and Ia (I-A alpha) mRNA expression during peritonitis is site dependent. J. Surg. Res. 1993, 54, 426–430. [Google Scholar] [CrossRef]
- Myers, M.J.; Pullen, J.K.; Ghildyal, N.; Eustis-Turf, E.; Schook, L.B. Regulation of IL-1 and TNF-alpha expression during the differentiation of bone marrow derived macrophage. J. Immunol. 1989, 142, 153–160. [Google Scholar]
- Shimoda, O.; Takeda, Y.; Woo, H.J.; Shimada, S.; Higuchi, M.; Osawa, T. A human macrophage hybridoma producing a cytotoxic factor distinct from TNF, LT, and IL-1. Cancer Immunol. Immunother. 1988, 26, 101–108. [Google Scholar]
- Cassatella, M.A.; Bazzoni, F.; Flynn, R.M.; Dusi, S.; Trinchieri, G.; Rossi, F. Molecular basis of interferon-gamma and lipopolysaccharide enhancement of phagocyte respiratory burst capability. Studies on the gene expression of several NADPH oxidase components. J. Biol. Chem. 1990, 265, 20241–20246. [Google Scholar]
- Cassatella, M.A.; Cappelli, R.; Della Bianca, V.; Grzeskowiak, M.; Dusi, S.; Berton, G. Interferon-gamma activates human neutrophil oxygen metabolism and exocytosis. Immunology 1988, 63, 499–506. [Google Scholar]
- Corradin, S.B.; Buchmuller-Rouiller, Y.; Mauel, J. Phagocytosis enhances murine macrophage activation by interferon-gamma and tumor necrosis factor-alpha. Eur. J. Immunol. 1991, 21, 2553–2558. [Google Scholar] [CrossRef]
- Fremond, C.M.; Togbe, D.; Doz, E.; Rose, S.; Vasseur, V.; Maillet, I.; Jacobs, M.; Ryffel, B.; Quesniaux, V.F. IL-1 receptor-mediated signal is an essential component of MyD88-dependent innate response to Mycobacterium tuberculosis infection. J. Immunol. 2007, 179, 1178–1189. [Google Scholar]
- Grayfer, L.; Belosevic, M. Molecular characterization, expression and functional analysis of goldfish (Carassius auratus L.) interferon gamma. Dev. Comp. Immunol. 2009, 33, 235–246. [Google Scholar] [CrossRef]
- Grayfer, L.; Garcia, E.G.; Belosevic, M. Comparison of macrophage antimicrobial responses induced by type II interferons of the goldfish (Carassius auratus L.). J. Biol. Chem. 2010, 285, 23537–23547. [Google Scholar]
- Grayfer, L.; Walsh, J.G.; Belosevic, M. Characterization and functional analysis of goldfish (Carassius auratus L.) tumor necrosis factor-alpha. Dev. Comp. Immunol. 2008, 32, 532–543. [Google Scholar] [CrossRef]
- Ishibe, K.; Yamanishi, T.; Wang, Y.; Osatomi, K.; Hara, K.; Kanai, K.; Yamaguchi, K.; Oda, T. Comparative analysis of the production of nitric oxide (NO) and tumor necrosis factor-alpha (TNF-alpha) from macrophages exposed to high virulent and low virulent strains of Edwardsiella tarda. Fish. Shellfish. Immunol. 2009, 27, 386–389. [Google Scholar] [CrossRef]
- Nathan, C.F.; Murray, H.W.; Wiebe, M.E.; Rubin, B.Y. Identification of interferon-gamma as the lymphokine that activates human macrophage oxidative metabolism and antimicrobial activity. J. Exp. Med. 1983, 158, 670–689. [Google Scholar] [CrossRef]
- Ordas, M.C.; Costa, M.M.; Roca, F.J.; Lopez-Castejon, G.; Mulero, V.; Meseguer, J.; Figueras, A.; Novoa, B. Turbot TNFalpha gene: Molecular characterization and biological activity of the recombinant protein. Mol. Immunol. 2007, 44, 389–400. [Google Scholar]
- Purcell, M.K.; Kurath, G.; Garver, K.A.; Herwig, R.P.; Winton, J.R. Quantitative expression profiling of immune response genes in rainbow trout following infectious haematopoietic necrosis virus (IHNV) infection or DNA vaccination. Fish Shellfish Immunol. 2004, 17, 447–462. [Google Scholar] [CrossRef]
- Purcell, M.K.; Nichols, K.M.; Winton, J.R.; Kurath, G.; Thorgaard, G.H.; Wheeler, P.; Hansen, J.D.; Herwig, R.P.; Park, L.K. Comprehensive gene expression profiling following DNA vaccination of rainbow trout against infectious hematopoietic necrosis virus. Mol. Immunol. 2006, 43, 2089–2106. [Google Scholar] [CrossRef]
- Xiao, J.; Zhou, Z.C.; Chen, C.; Huo, W.L.; Yin, Z.X.; Weng, S.P.; Chan, S.M.; Yu, X.Q.; He, J.G. Tumor necrosis factor-alpha gene from mandarin fish, Siniperca chuatsi: Molecular cloning, cytotoxicity analysis and expression profile. Mol. Immunol. 2007, 44, 3615–3622. [Google Scholar]
- Bird, S.; Wang, T.; Zou, J.; Cunningham, C.; Secombes, C.J. The first cytokine sequence within cartilaginous fish: IL-1 beta in the small spotted catshark (Scyliorhinus canicula). J. Immunol. 2002, 168, 3329–3340. [Google Scholar]
- Hirono, I.; Nam, B.H.; Kurobe, T.; Aoki, T. Molecular cloning, characterization, and expression of TNF cDNA and gene from Japanese flounder Paralychthys olivaceus. J. Immunol. 2000, 165, 4423–4427. [Google Scholar]
- Igawa, D.; Sakai, M.; Savan, R. An unexpected discovery of two interferon gamma-like genes along with interleukin (IL)-22 and -26 from teleost: IL-22 and -26 genes have been described for the first time outside mammals. Mol. Immunol. 2006, 43, 999–1009. [Google Scholar] [CrossRef]
- Nishikawa, A.; Murata, E.; Akita, M.; Kaneko, K.; Moriya, O.; Tomita, M.; Hayashi, H. Roles of macrophages in programmed cell death and remodeling of tail and body muscle of Xenopus laevis during metamorphosis. Histochem. Cell Biol. 1998, 109, 11–17. [Google Scholar]
- Auffray, C.; Fogg, D.; Garfa, M.; Elain, G.; Join-Lambert, O.; Kayal, S.; Sarnacki, S.; Cumano, A.; Lauvau, G.; Geissmann, F. Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science 2007, 317, 666–670. [Google Scholar]
- Nahrendorf, M.; Swirski, F.K.; Aikawa, E.; Stangenberg, L.; Wurdinger, T.; Figueiredo, J.L.; Libby, P.; Weissleder, R.; Pittet, M.J. The healing myocardium sequentially mobilizes two monocyte subsets with divergent and complementary functions. J. Exp. Med. 2007, 204, 3037–3047. [Google Scholar]
- Zhao, C.; Zhang, H.; Wong, W.C.; Sem, X.; Han, H.; Ong, S.M.; Tan, Y.C.; Yeap, W.H.; Gan, C.S.; Ng, K.Q.; et al. Identification of novel functional differences in monocyte subsets using proteomic and transcriptomic methods. J. Proteome Res. 2009, 8, 4028–4038. [Google Scholar]
- Ziegler-Heitbrock, L. The CD14+ CD16+ blood monocytes: Their role in infection and inflammation. J. Leukoc. Biol. 2007, 81, 584–592. [Google Scholar] [CrossRef]
- Flajnik, M.F.; Kaufman, J.F.; Hsu, E.; Manes, M.; Parisot, R.; Du Pasquier, L. Major histocompatibility complex-encoded class I molecules are absent in immunologically competent Xenopus before metamorphosis. J. Immunol. 1986, 137, 3891–3899. [Google Scholar]
- Kerr, I.M.; Brown, R.E.; Hovanessian, A.G. Nature of inhibitor of cell-free protein synthesis formed in response to interferon and double-stranded RNA. Nature 1977, 268, 540–542. [Google Scholar]
- Meurs, E.; Chong, K.; Galabru, J.; Thomas, N.S.; Kerr, I.M.; Williams, B.R.; Hovanessian, A.G. Molecular cloning and characterization of the human double-stranded RNA-activated protein kinase induced by interferon. Cell 1990, 62, 379–390. [Google Scholar] [CrossRef]
- Roberts, W.K.; Hovanessian, A.; Brown, R.E.; Clemens, M.J.; Kerr, I.M. Interferon-mediated protein kinase and low-molecular-weight inhibitor of protein synthesis. Nature 1976, 264, 477–480. [Google Scholar]
- George, C.X.; Thomis, D.C.; McCormack, S.J.; Svahn, C.M.; Samuel, C.E. Characterization of the heparin-mediated activation of PKR, the interferon-inducible RNA-dependent protein kinase. Virology 1996, 221, 180–188. [Google Scholar] [CrossRef]
- Ruvolo, P.P.; Gao, F.; Blalock, W.L.; Deng, X.; May, W.S. Ceramide regulates protein synthesis by a novel mechanism involving the cellular PKR activator RAX. J. Biol. Chem. 2001, 276, 11754–11758. [Google Scholar]
- Gil, J.; Alcami, J.; Esteban, M. Induction of apoptosis by double-stranded-RNA-dependent protein kinase (PKR) involves the alpha subunit of eukaryotic translation initiation factor 2 and NF-kappaB. Mol. Cell. Biol. 1999, 19, 4653–4663. [Google Scholar]
- Langland, J.O.; Cameron, J.M.; Heck, M.C.; Jancovich, J.K.; Jacobs, B.L. Inhibition of PKR by RNA and DNA viruses. Virus. Res. 2006, 119, 100–110. [Google Scholar] [CrossRef]
- Essbauer, S.; Bremont, M.; Ahne, W. Comparison of the eIF-2alpha homologous proteins of seven ranaviruses (Iridoviridae). Virus Genes 2001, 23, 347–359. [Google Scholar] [CrossRef]
- Chen, G.; Ward, B.M.; Yu, K.H.; Chinchar, V.G.; Robert, J. Improved knockout methodology reveals that frog virus 3 mutants lacking either the 18K immediate-early gene or the truncated vIF-2alpha gene are defective for replication and growth in vivo. J. Virol. 2011, 85, 11131–11138. [Google Scholar] [CrossRef]
- Rothenburg, S.; Chinchar, V.G.; Dever, T.E. Characterization of a ranavirus inhibitor of the antiviral protein kinase PKR. BMC Microbiol. 2011, 11. [Google Scholar]
- Majji, S.; Thodima, V.; Sample, R.; Whitley, D.; Deng, Y.; Mao, J.; Chinchar, V.G. Transcriptome analysis of Frog virus 3, the type species of the genus Ranavirus, family Iridoviridae. Virology 2009, 391, 293–303. [Google Scholar] [CrossRef]
- Whitley, D.S.; Sample, R.C.; Sinning, A.R.; Henegar, J.; Chinchar, V.G. Antisense approaches for elucidating ranavirus gene function in an infected fish cell line. Dev. Comp. Immunol. 2011, 35, 937–948. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Grayfer, L.; Andino, F.D.J.; Chen, G.; Chinchar, G.V.; Robert, J. Immune Evasion Strategies of Ranaviruses and Innate Immune Responses to These Emerging Pathogens. Viruses 2012, 4, 1075-1092. https://doi.org/10.3390/v4071075
Grayfer L, Andino FDJ, Chen G, Chinchar GV, Robert J. Immune Evasion Strategies of Ranaviruses and Innate Immune Responses to These Emerging Pathogens. Viruses. 2012; 4(7):1075-1092. https://doi.org/10.3390/v4071075
Chicago/Turabian StyleGrayfer, Leon, Francisco De Jesús Andino, Guangchun Chen, Gregory V. Chinchar, and Jacques Robert. 2012. "Immune Evasion Strategies of Ranaviruses and Innate Immune Responses to These Emerging Pathogens" Viruses 4, no. 7: 1075-1092. https://doi.org/10.3390/v4071075