
Evolution Characterization and Pathogenicity of an NADC34-like PRRSV Isolated from Inner Mongolia, China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Virus Isolation
2.2. Electron Microscopic Observation
2.3. Immunofluorescence Identification
2.4. Primers’ Design and Amplification of Genomic Full Length
2.5. Sequencing Analysis
2.6. Animals and Experimental Design
2.7. Determination of Viral Load in Serum, Tissue, Nasal and Fecal Swabs
2.8. Antibody Detection Assay
2.9. Histopathology and Immunohistochemical Staining
3. Results
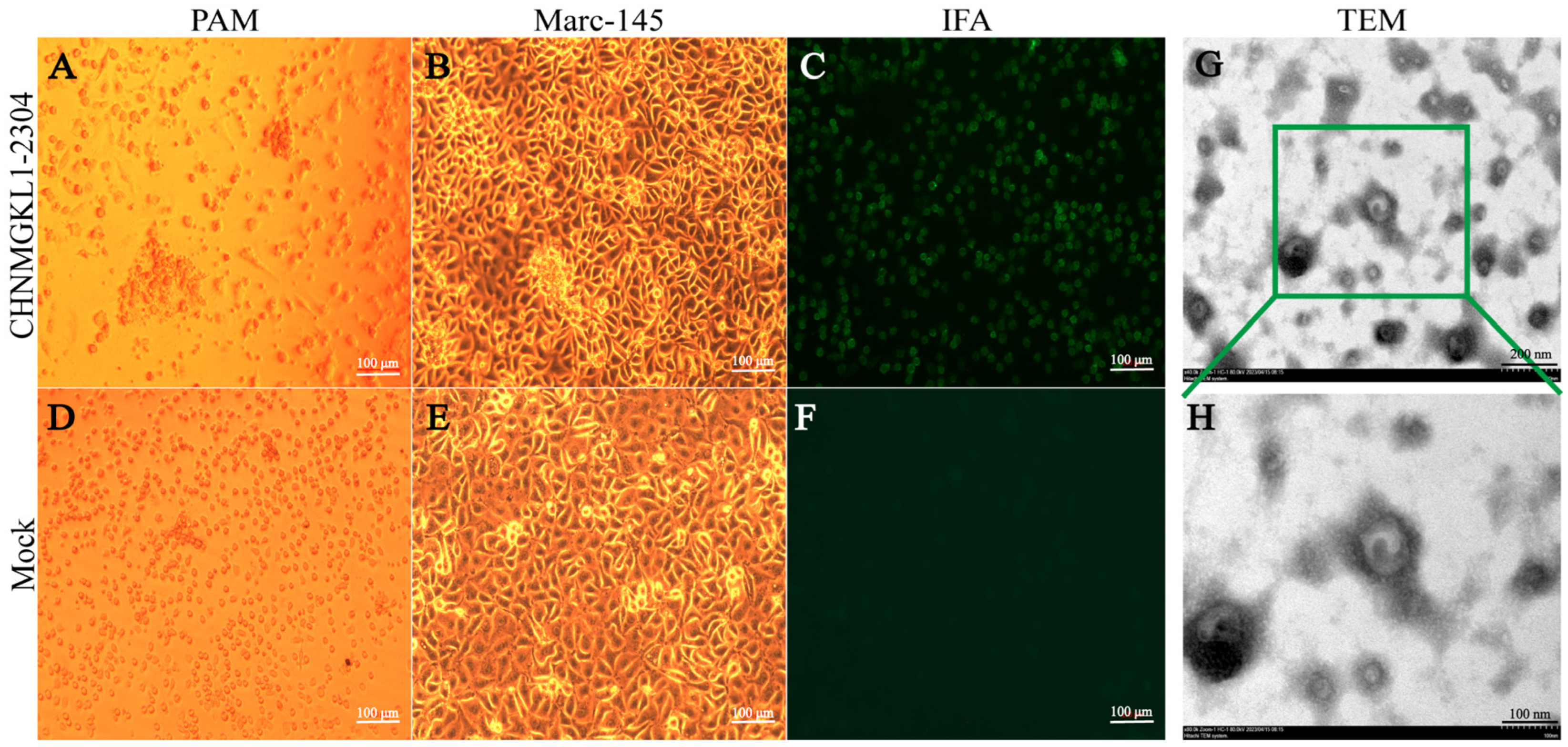
3.1. Results of Virus Isolation and Identification
3.2. Amplification and Sequencing of Full Genome
3.3. Comparison Results of Homologous Sequences
3.4. Results of Reorganization Analysis
3.5. Phylogenetic Analysis Based on ORF5 Gene and Full-Length Genome
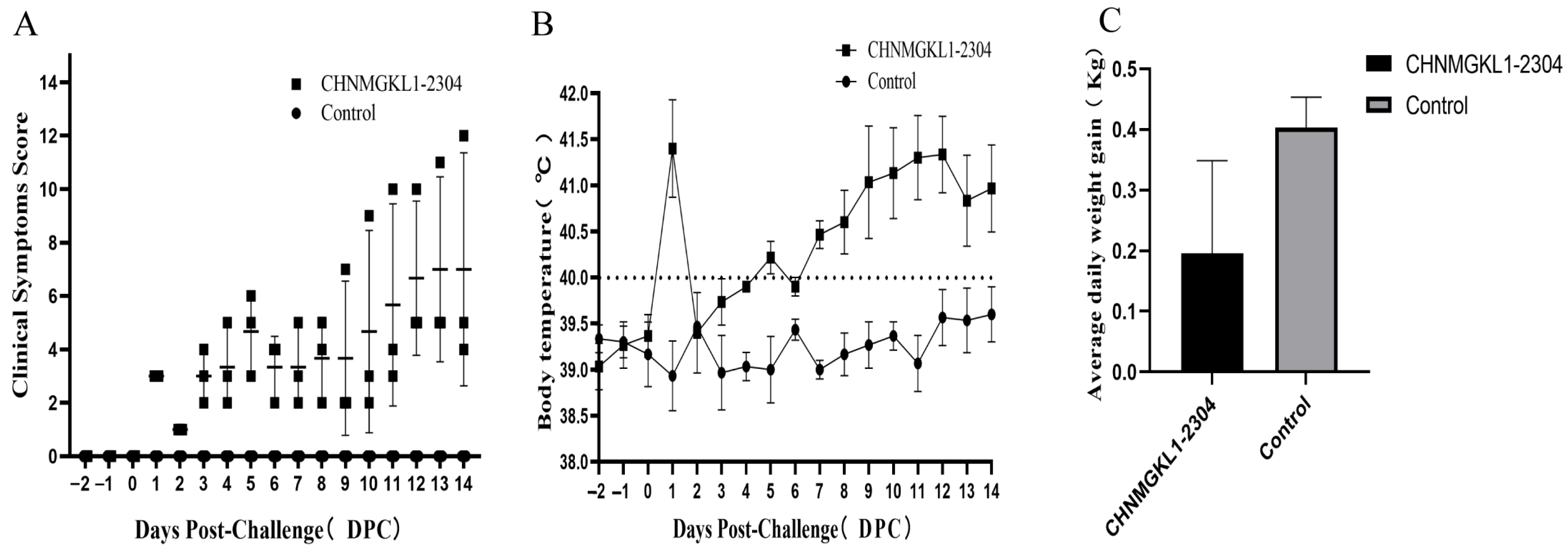
3.6. Clinical Symptom of Piglets Infected with CHNMGKL1-2304
3.7. Viral Load and Antibody Level
3.8. Histopathology
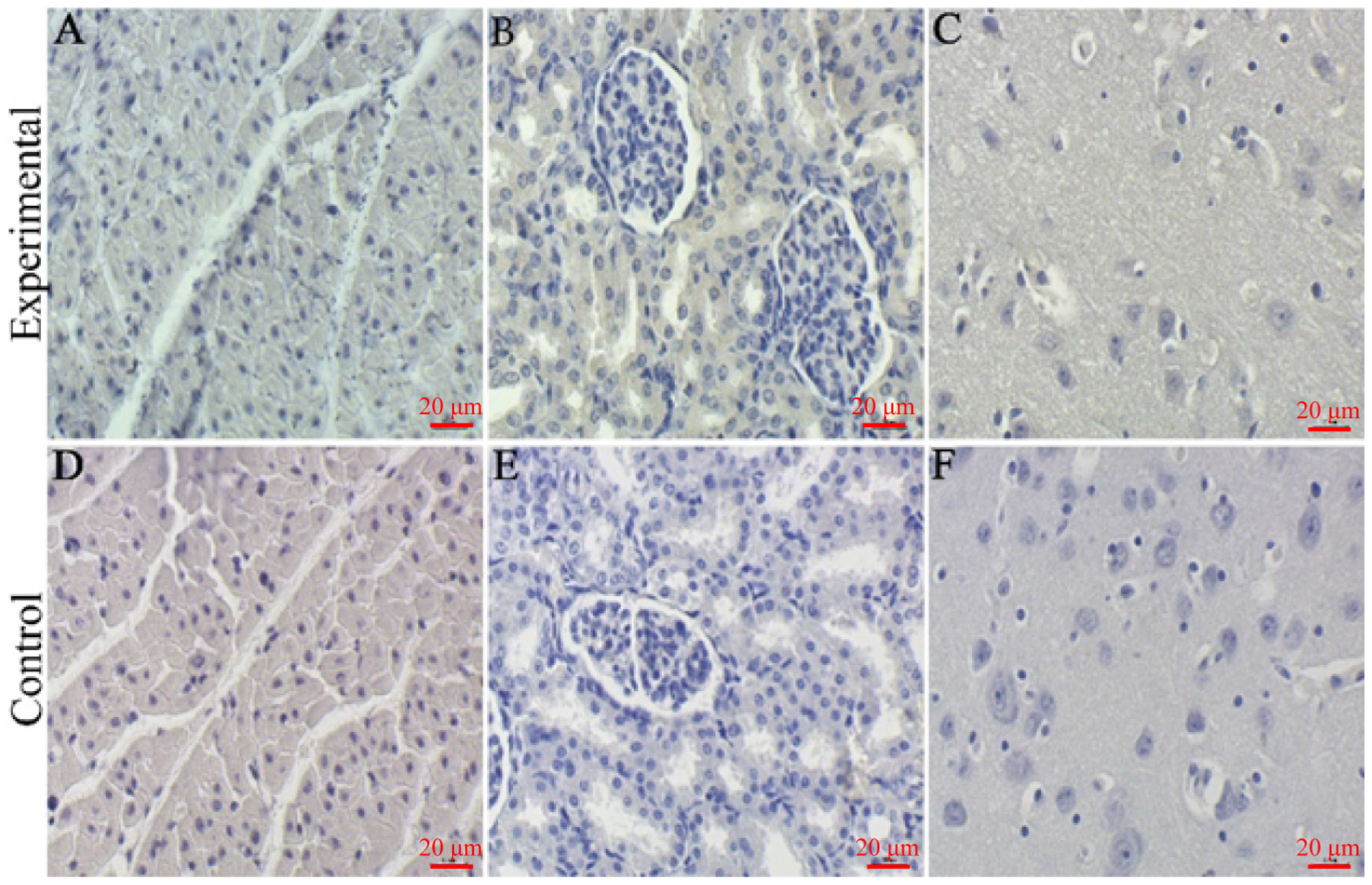
3.9. IHC Staining
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; ter Laak, E.A.; Bloemraad, M.; de Kluyver, E.P.; Kragten, C.; van Buiten, L.; den Besten, A.; Wagenaar, F.; et al. Mystery swine disease in The Netherlands: The isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, H.; Nunoya, T.; Tajima, M.; Kato, A.; Samejima, T. An outbreak of porcine reproductive and respiratory syndrome in Japan. J. Vet. Med. Sci. 1994, 56, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Albina, E. Epidemiology of porcine reproductive and respiratory syndrome (PRRS): An overview. Vet. Microbiol. 1997, 55, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Elazhary, Y.; Weber, J.; Bikour, H.; Morin, M.; Girard, C. ‘Mystery swine disease’ in Canada. Vet. Rec. 1991, 129, 495–496. [Google Scholar] [CrossRef] [PubMed]
- Nathues, H.; Alarcon, P.; Rushton, J.; Jolie, R.; Fiebig, K.; Jimenez, M.; Geurts, V.; Nathues, C. Cost of porcine reproductive and respiratory syndrome virus at individual farm level—An economic disease model. Prev. Vet. Med. 2017, 142, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Neumann, E.J.; Kliebenstein, J.B.; Johnson, C.D.; Mabry, J.W.; Bush, E.J.; Seitzinger, A.H.; Green, A.L.; Zimmerman, J.J. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J. Am. Vet. Med. Assoc. 2005, 227, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuis, N.; Duinhof, T.F.; van Nes, A. Economic analysis of outbreaks of porcine reproductive and respiratory syndrome virus in nine sow herds. Vet. Rec. 2012, 170, 225. [Google Scholar] [CrossRef] [PubMed]
- Dea, S.; Gagnon, C.A.; Mardassi, H.; Pirzadeh, B.; Rogan, D. Current knowledge on the structural proteins of porcine reproductive and respiratory syndrome (PRRS) virus: Comparison of the North American and European isolates. Arch. Virol. 2000, 145, 659–688. [Google Scholar] [CrossRef] [PubMed]
- Dokland, T. The structural biology of PRRSV. Virus Res. 2010, 154, 86–97. [Google Scholar] [CrossRef]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479–480, 475–486. [Google Scholar] [CrossRef]
- Zhao, H.Z.; Wang, F.X.; Han, X.Y.; Guo, H.; Liu, C.Y.; Hou, L.N.; Wang, Y.X.; Zheng, H.; Wang, L.; Wen, Y.J. Recent advances in the study of NADC34-like porcine reproductive and respiratory syndrome virus in China. Front. Microbiol. 2022, 13, 950402. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, C.; Li, W.; Zhao, J.; Gong, B.; Sun, Q.; Tang, Y.D.; Xiang, L.; Leng, C.; Peng, J.; et al. Novel characteristics of Chinese NADC34-like PRRSV during 2020-2021. Transbound. Emerg. Dis. 2022, 69, e3215–e3224. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.F.; Liu, Y.; Yang, J.; Li, W.Z.; Yu, X.X.; Wang, S.Y.; Li, L.A.; Yu, H. Recombination between NADC34-like and QYYZ-like strain of porcine reproductive and respiratory syndrome virus with high pathogenicity for piglets in China. Transbound. Emerg. Dis. 2022, 69, e3202–e3207. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Zhu, Z.; Fan, J.; Liu, P.; Li, Y.; Li, Q.; Sun, Z.; Yu, X.; Lee, H.S.; Tian, K.; et al. High Pathogenicity of a Chinese NADC34-like PRRSV on Pigs. Microbiol. Spectr. 2022, 10, e0154122. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.Z.; Ha, Z.; Zhang, H.; Zhang, Y.; Xie, Y.B.; Zhang, H.; Nan, F.L.; Wang, Z.; Zhang, P.; Xu, W.; et al. Pathogenicity of porcine reproductive and respiratory syndrome virus (ORF5 RFLP 1-7-4 viruses) in China. Transbound. Emerg. Dis. 2020, 67, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, L.; Zhang, J.; Ge, X.; Zhou, R.; Zheng, H.; Geng, G.; Guo, X.; Yang, H. Nsp9 and Nsp10 contribute to the fatal virulence of highly pathogenic porcine reproductive and respiratory syndrome virus emerging in China. PLoS Pathog. 2014, 10, e1004216. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, Z.; Chen, K.; Qian, J.; Hu, Y.; Fang, S.; Sun, Z.; Zhang, C.; Huang, L.; Zhang, J.; et al. Efficacy of the Synergy Between Live-Attenuated and Inactivated PRRSV Vaccines Against a NADC30-Like Strain of Porcine Reproductive and Respiratory Syndrome Virus in 4-Week Piglets. Front. Vet. Sci. 2022, 9, 812040. [Google Scholar] [CrossRef] [PubMed]
- Wernike, K.; Bonilauri, P.; Dauber, M.; Errington, J.; LeBlanc, N.; Revilla-Fernandez, S.; Hjulsager, C.; Isaksson, M.; Stadejek, T.; Beer, M.; et al. Porcine reproductive and respiratory syndrome virus: Interlaboratory ring trial to evaluate real-time reverse transcription polymerase chain reaction detection methods. J. Vet. Diagn. Investig. 2012, 24, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Budwit-Novotny, D.A.; McCarty, K.S.; Cox, E.B.; Soper, J.T.; Mutch, D.G.; Creasman, W.T.; Flowers, J.L.; McCarty, K.S., Jr. Immunohistochemical analyses of estrogen receptor in endometrial adenocarcinoma using a monoclonal antibody. Cancer Res. 1986, 46, 5419–5425. [Google Scholar]
- Wang, F.X.; Song, N.; Chen, L.Z.; Cheng, S.P.; Wu, H.; Wen, Y.J. Non-structural protein 2 of the porcine reproductive and respiratory syndrome (PRRS) virus: A crucial protein in viral pathogenesis, immunity and diagnosis. Res. Vet. Sci. 2013, 95, 1–7. [Google Scholar] [CrossRef]
- Zhang, H.L.; Zhang, W.L.; Xiang, L.R.; Leng, C.L.; Tian, Z.J.; Tang, Y.D.; Cai, X.H. Emergence of novel porcine reproductive and respiratory syndrome viruses (ORF5 RFLP 1-7-4 viruses) in China. Vet. Microbiol. 2018, 222, 105–108. [Google Scholar] [CrossRef]
- Liu, J.; Wei, C.; Lin, Z.; Xia, W.; Ma, Y.; Dai, A.; Yang, X. Full genome sequence analysis of a 1-7-4-like PRRSV strain in Fujian Province, China. PeerJ 2019, 7, e7859. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.J.; Xie, W.; Chen, X.X.; Qiao, S.; Zhao, M.; Gu, Y.; Zhao, B.L.; Zhang, G. Molecular epidemiology of porcine reproductive and respiratory syndrome virus in Central China since 2014: The prevalence of NADC30-like PRRSVs. Microb. Pathog. 2017, 109, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Xu, H.; Zhao, J.; Leng, C.; Xiang, L.; Li, C.; Fu, J.; Tang, Y.D.; Peng, J.; Wang, Q.; et al. Pathogenicity of NADC34-like PRRSV HLJDZD32-1901 isolated in China. Vet. Microbiol. 2020, 246, 108727. [Google Scholar] [CrossRef] [PubMed]
- Du, T.; Nan, Y.; Xiao, S.; Zhao, Q.; Zhou, E.M. Antiviral Strategies against PRRSV Infection. Trends Microbiol. 2017, 25, 968–979. [Google Scholar] [CrossRef]
- Straw, B.E.; Zimmerman, J.J.; D’Allaire, S.; Taylor, D.J. Diseases of Swine; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Guo, B.; Lager, K.M.; Henningson, J.N.; Miller, L.C.; Schlink, S.N.; Kappes, M.A.; Kehrli, M.E., Jr.; Brockmeier, S.L.; Nicholson, T.L.; Yang, H.C.; et al. Experimental infection of United States swine with a Chinese highly pathogenic strain of porcine reproductive and respiratory syndrome virus. Virology 2013, 435, 372–384. [Google Scholar] [CrossRef]
- Karniychuk, U.U.; Geldhof, M.; Vanhee, M.; Van Doorsselaere, J.; Saveleva, T.A.; Nauwynck, H.J. Pathogenesis and antigenic characterization of a new East European subtype 3 porcine reproductive and respiratory syndrome virus isolate. BMC Vet. Res. 2010, 6, 30. [Google Scholar] [CrossRef]
- Sanchez-Carvajal, J.M.; Rodriguez-Gomez, I.M.; Ruedas-Torres, I.; Larenas-Munoz, F.; Diaz, I.; Revilla, C.; Mateu, E.; Dominguez, J.; Martin-Valls, G.; Barranco, I.; et al. Activation of pro- and anti-inflammatory responses in lung tissue injury during the acute phase of PRRSV-1 infection with the virulent strain Lena. Vet. Microbiol. 2020, 246, 108744. [Google Scholar] [CrossRef] [PubMed]
- Murtaugh, M.P.; Xiao, Z.; Zuckermann, F. Immunological responses of swine to porcine reproductive and respiratory syndrome virus infection. Viral Immunol. 2002, 15, 533–547. [Google Scholar] [CrossRef]
- Plagemann, P.G. Neutralizing antibody formation in swine infected with seven strains of porcine reproductive and respiratory syndrome virus as measured by indirect ELISA with peptides containing the GP5 neutralization epitope. Viral Immunol. 2006, 19, 285–293. [Google Scholar] [CrossRef]
- Lee, J.A.; Kwon, B.; Osorio, F.A.; Pattnaik, A.K.; Lee, N.H.; Lee, S.W.; Park, S.Y.; Song, C.S.; Choi, I.S.; Lee, J.B. Protective humoral immune response induced by an inactivated porcine reproductive and respiratory syndrome virus expressing the hypo-glycosylated glycoprotein 5. Vaccine 2014, 32, 3617–3622. [Google Scholar] [CrossRef] [PubMed]
- Delputte, P.L.; Meerts, P.; Costers, S.; Nauwynck, H.J. Effect of virus-specific antibodies on attachment, internalization and infection of porcine reproductive and respiratory syndrome virus in primary macrophages. Vet. Immunol. Immunopathol. 2004, 102, 179–188. [Google Scholar] [CrossRef] [PubMed]

 indicates the reference virus.
indicates the reference virus.  and
and  indicate the strain isolated from Inner Mongolia. Blue box: group one. Green box: group two. Red box: group three).
indicates the reference virus. and indicate the strain isolated from Inner Mongolia. Blue box: group one. Green box: group two. Red box: group three).
indicate the strain isolated from Inner Mongolia. Blue box: group one. Green box: group two. Red box: group three).
indicates the reference virus. and indicate the strain isolated from Inner Mongolia. Blue box: group one. Green box: group two. Red box: group three).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequences (5′–3′) | Annealing Temperature (°C) | Size (bp) |
|---|---|---|---|
| 5′UTR-A | F:ATGACGTATAGGTGTTGGCTC | 52 | 335 |
| R:CAAGCCCAACACTCCAAG | |||
| 34-1-B | F:GCACCTTGCTTCTGGAGT | 54 | 2591 |
| R: GCGTTGTAGTTGTTAGTTTCG | |||
| 34-2-C | F:TGCCAGATTGTAAGCCCGTCCCT | 57 | 2646 |
| R:ACCAGCGTAACCAGCAAGGAA | |||
| 34-3-D | F: CCCGTTTGCCGTTCCTGGTT | 56 | 2350 |
| R: TGCGTCCGTGTTGTCGTG | |||
| 34-4-E | F: CCCGTCGGCAGTATCTTT | 54 | 2758 |
| R: AAGACTTCGTCCAGAGGG | |||
| 34-5-F | F: GAACTTGTGGTTGGGATG | 54 | 2758 |
| R: TGCTCAGAGTGAACGGTAG | |||
| 34-6-G | F: GGGAAGATTACAATGACGC | 53 | 2573 |
| R: TTAAGAGGTGCAAGAGCC | |||
| 34-7-H | F: GCGGAACAATGGGGTCGT | 55 | 1047 |
| R: AACCATGCGGCCGTAATTAA |
| Sublineage 1.5 | Sublineage 1.8 | Sublineage 3.5 | Sublineage 5.1 | Sublineage 8.1 | ||
|---|---|---|---|---|---|---|
| Nucleotides | JS2021NADC34 | IA/2014/NADC34 | NADC30 | QYYZ | VR2332 | Ch-1R |
| Complete genome | 97.7 | 94.9 | 84.9 | 81.2 | 83.1 | 83.1 |
| 5′UTR | 94.7 | 93.6 | 91.5 | 92.0 | 89.5 | 89.4 |
| Nsp1 | 98.7 | 94.5 | 84.8 | 81.3 | 83.8 | 82.9 |
| Nsp2 | 97.0 | 92.1 | 75.1 | 71.1 | 73.7 | 74.7 |
| Nsp3 | 98.5 | 96.1 | 84.9 | 79.6 | 83.7 | 81.8 |
| Nsp4 | 98.4 | 96.4 | 81.4 | 79.7 | 82.7 | 83.2 |
| Nsp5 | 96.9 | 92.2 | 83.5 | 79.8 | 84.3 | 83.7 |
| Nsp6 | 97.9 | 95.8 | 89.6 | 85.4 | 91.7 | 85.4 |
| Nsp7 | 97.2 | 93.3 | 81.5 | 78.9 | 81.6 | 80.1 |
| Nsp8 | 97.0 | 96.3 | 90.4 | 88.9 | 89.6 | 88.9 |
| Nsp9 | 98.2 | 96.0 | 88.6 | 85.6 | 86.8 | 86.6 |
| Nsp10 | 98.6 | 96.0 | 90.2 | 84.5 | 85.7 | 85.8 |
| Nsp11 | 99.0 | 96.4 | 85.3 | 85.6 | 85.3 | 87.1 |
| Nsp12 | 97.8 | 95.5 | 83.3 | 83.5 | 82.6 | 82.1 |
| ORF2a | 96.6 | 95.1 | 85.7 | 86.1 | 86.9 | 87.0 |
| ORF2b | 96.8 | 95.9 | 87.8 | 91.0 | 89.2 | 88.7 |
| ORF3 | 96.5 | 94.8 | 85.0 | 82.1 | 83.4 | 83.0 |
| ORF4 | 97.8 | 94.6 | 93.1 | 84.4 | 86.4 | 85.5 |
| ORF5 | 97.0 | 95.9 | 86.9 | 83.1 | 85.7 | 85.7 |
| ORF6 | 97.9 | 97.7 | 93.5 | 88.8 | 88.6 | 87.6 |
| ORF7 | 96.2 | 96.0 | 94.1 | 87.6 | 89.8 | 90.9 |
| 3′UTR | 99.3 | 99.3 | 96.7 | 90.1 | 88.0 | 88.2 |
| Nucleotides | LNWK96 | LNWK130 | FJ0908 | CH/SCMY-2/2019 | HLHDZD32-1901 | HLJPY18-2009 | HLJZD30-1902 | CH/SCLS-2/2020 | PRRSV-ZDXYL-CHINA-2018-1 | PRRSV-ZDXYL-China-2018-2 |
|---|---|---|---|---|---|---|---|---|---|---|
| Complete genome | 92.2 | 92.8 | 93.3 | 93.2 | 93.5 | 88.4 | 94.3 | 94.4 | 94 | 93.8 |
| 5′UTR | 91.0 | 94.0 | 91.5 | 93.6 | 93.6 | 89.9 | 93.0 | 93.1 | 93.6 | 93.1 |
| Nsp1 | 89.8 | 89.8 | 91.8 | 93.0 | 93.0 | 80.7 | 94.1 | 94.3 | 93.0 | 92.9 |
| Nsp2 | 88.9 | 88.6 | 89.4 | 91.3 | 90.3 | 72.3 | 91.0 | 91.0 | 91.3 | 90.9 |
| Nsp3 | 93.6 | 93.3 | 94.2 | 94.8 | 94.5 | 91.0 | 95.1 | 96.1 | 94.8 | 94.4 |
| Nsp4 | 95.3 | 94.1 | 95.1 | 96.2 | 95.8 | 95.1 | 95.9 | 96.1 | 96.2 | 95.6 |
| Nsp5 | 90.8 | 91.6 | 91.0 | 91.6 | 90.6 | 90.6 | 91.6 | 91.6 | 91.6 | 90.2 |
| Nsp6 | 95.8 | 95.8 | 93.8 | 97.9 | 93.8 | 97.9 | 93.8 | 95.8 | 97.9 | 95.8 |
| Nsp7 | 92.4 | 93.2 | 93.4 | 93.3 | 91.0 | 92.4 | 93.3 | 92.9 | 93.3 | 92.1 |
| Nsp8 | 96.3 | 93.3 | 94.8 | 94.1 | 95.6 | 96.3 | 95.6 | 95.6 | 94.1 | 94.8 |
| Nsp9 | 94.8 | 94.7 | 95.1 | 94.9 | 94.9 | 89.3 | 95.6 | 96.0 | 94.9 | 94.8 |
| Nsp10 | 95.0 | 95.2 | 94.9 | 95.2 | 94.7 | 93.3 | 95.7 | 96.0 | 95.6 | 95.3 |
| Nsp11 | 95.5 | 96.0 | 95.8 | 94.0 | 95.7 | 95.7 | 96.3 | 96.1 | 95.4 | 96.3 |
| Nsp12 | 94.4 | 94.2 | 94.2 | 89.6 | 94.6 | 94.0 | 94.8 | 95.0 | 95.0 | 94.0 |
| ORF2α | 94.2 | 94.3 | 94.6 | 94.0 | 93.6 | 93.4 | 94.2 | 94.7 | 93.6 | 93.9 |
| ORF2β | 94.6 | 94.6 | 95.5 | 95.5 | 95.0 | 95.5 | 95.0 | 95.5 | 95.0 | 95.5 |
| ORF3 | 89.9 | 92.2 | 92.2 | 92.3 | 93.5 | 93.1 | 94.6 | 94.4 | 94.1 | 94.0 |
| ORF4 | 91.2 | 92.4 | 92.2 | 89.8 | 93.9 | 93.5 | 95.0 | 94.4 | 94.4 | 95.0 |
| ORF5 | 85.2 | 94.2 | 94.4 | 91.9 | 95.0 | 95.2 | 95.5 | 91.7 | 94.4 | 95.9 |
| ORF6 | 93.3 | 95.8 | 95.8 | 91.0 | 96.6 | 97.1 | 96.2 | 97.1 | 95.6 | 95.8 |
| ORF7 | 91.7 | 93.3 | 93.5 | 92.7 | 94.9 | 93.3 | 95.4 | 95.2 | 95.2 | 94.9 |
| 3′UTR | 96.0 | 97.1 | 98.7 | 94.7 | 96.1 | 98.0 | 98.0 | 100.0 | 98.5 | 96.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.-Z.; Liu, C.-Y.; Meng, H.; Sun, C.-L.; Yang, H.-W.; Wang, H.; Zou, J.; Li, P.; Han, F.-Y.; Qi, G.; et al. Evolution Characterization and Pathogenicity of an NADC34-like PRRSV Isolated from Inner Mongolia, China. Viruses 2024, 16, 683. https://doi.org/10.3390/v16050683
Zhao H-Z, Liu C-Y, Meng H, Sun C-L, Yang H-W, Wang H, Zou J, Li P, Han F-Y, Qi G, et al. Evolution Characterization and Pathogenicity of an NADC34-like PRRSV Isolated from Inner Mongolia, China. Viruses. 2024; 16(5):683. https://doi.org/10.3390/v16050683
Chicago/Turabian StyleZhao, Hong-Zhe, Chun-Yu Liu, Hai Meng, Cheng-Long Sun, Hong-Wen Yang, Hao Wang, Jian Zou, Peng Li, Feng-Ye Han, Gen Qi, and et al. 2024. "Evolution Characterization and Pathogenicity of an NADC34-like PRRSV Isolated from Inner Mongolia, China" Viruses 16, no. 5: 683. https://doi.org/10.3390/v16050683