
Susceptibility of Mediterranean Buffalo (Bubalus bubalis) following Experimental Infection with Lumpy Skin Disease Virus
 , ,
, ,  , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Infection
2.3. Clinical Observation
2.4. Samples
2.5. Serological Examination
2.6. DNA Extraction and Molecular Analysis
3. Results
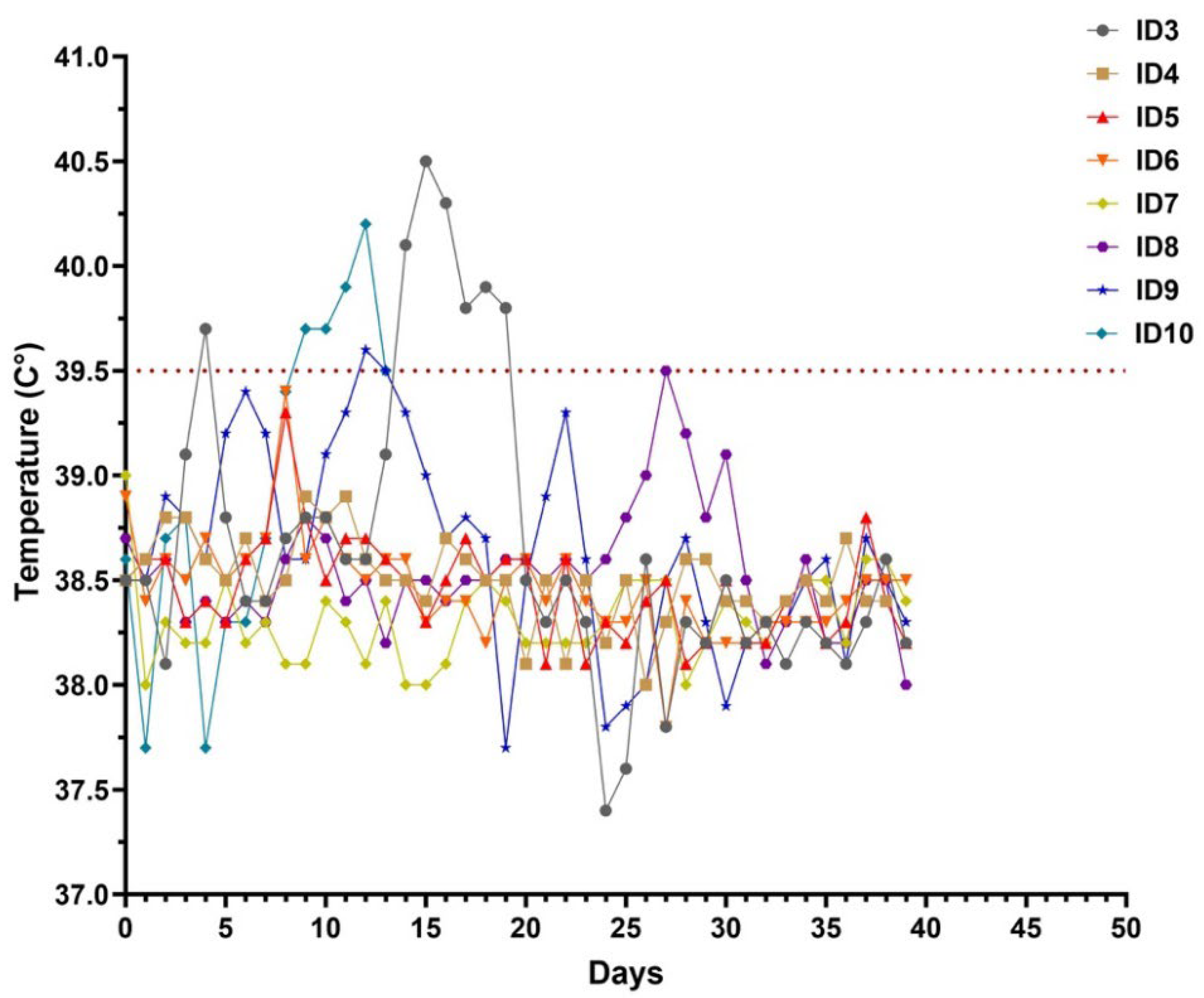
3.1. Clinical Observation
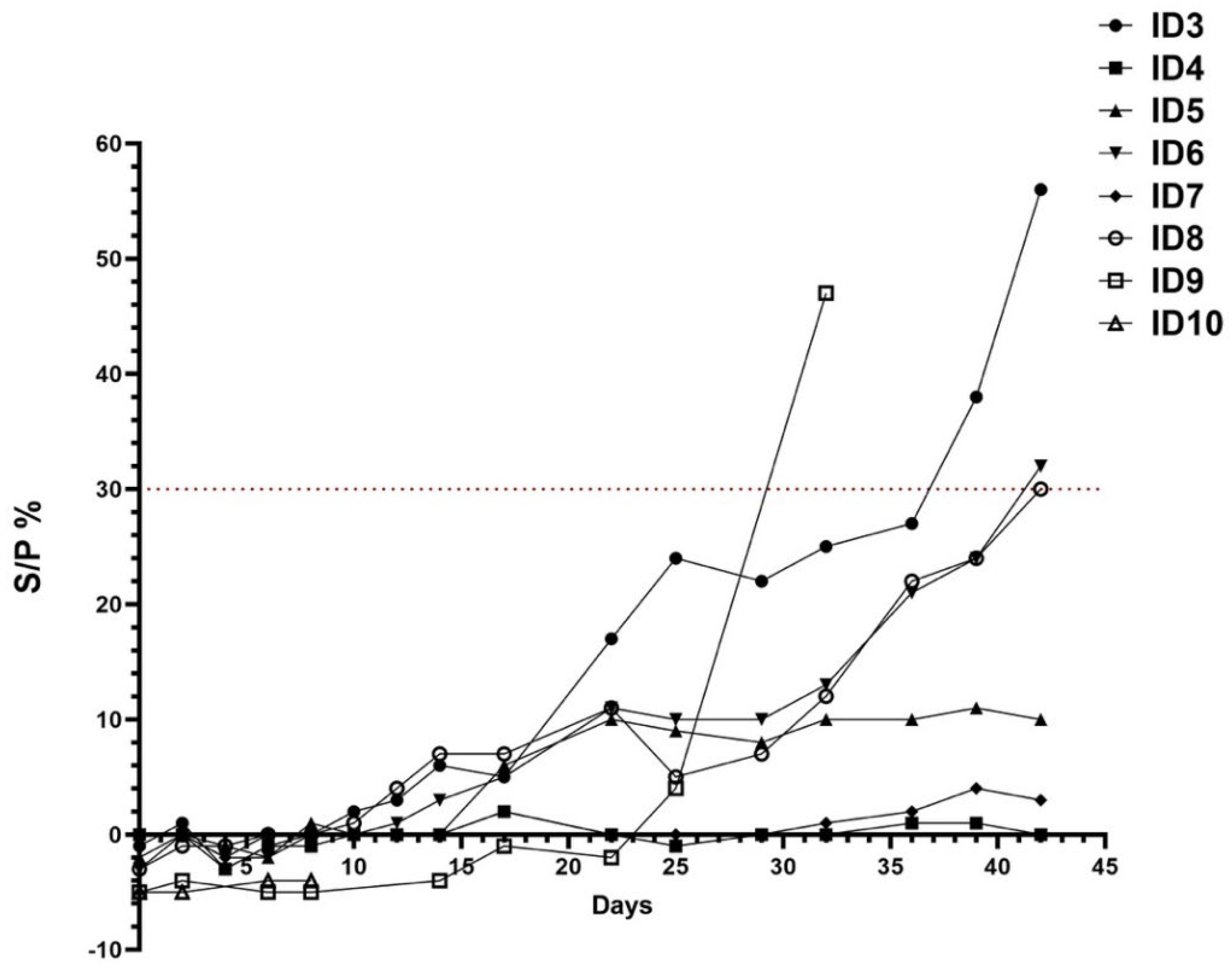
3.2. Serological Analysis
3.3. Molecular Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tuppurainen, E.S.M.; Venter, E.H.; Shisler, J.L.; Gari, G.; Mekonnen, G.A.; Juleff, N.; Lyons, N.A.; De Clercq, K.; Upton, C.; Bowden, T.R.; et al. Review: Capripoxvirus Diseases: Current Status and Opportunities for Control. Transbound. Emerg. Dis. 2017, 64, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Abera, Z.; Box, P.O.; Abera, Z.; Degefu, H.; Gari, G.; Ayana, Z. Review on Epidemiology and Economic Importance of Lumpy Skin Disease. Int. J. Basic Appl. Virol. 2015, 4, 8. [Google Scholar]
- Saegerman, C.; Bertagnoli, S.; Meyer, G.; Ganiere, J.; Caufour, P.; De Clercq, K.; Jacquiet, P.; Fournie, G.; Hautefeuille, C.; Etore, F.; et al. Risk of Introduction of Lumpy Skin Disease in France by the Import of Vectors in Animal Trucks. PLoS ONE 2018, 13, e0198506. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.S.; Venter, E.H.; Coetzer, J.A.W. The Detection of Lumpy Skin Disease Virus in Samples of Experimentally Infected Cattle using Different Diagnostic Techniques. Onderstepoort J. Vet. Res. 2005, 72, 153–164. [Google Scholar]
- Sprygin, A.; Pestova, Y.; Wallace, D.B.; Tuppurainen, E.; Kononov, A.V. Transmission of Lumpy Skin Disease Virus: A Short Review. Virus Res. 2019, 269, 197637. [Google Scholar] [CrossRef] [PubMed]
- Irons, P.C.; Tuppurainen, E.S.M.; Venter, E.H. Excretion of Lumpy Skin Disease Virus in Bull Semen. Theriogenology 2005, 63, 1290–1297. [Google Scholar] [CrossRef] [PubMed]
- Babiuk, S.; Bowden, T.R.; Parkyn, G.; Dalman, B.; Manning, L.; Neufeld, J.; Embury-Hyatt, C.; Copps, J.; Boyle, D.B. Quantification of Lumpy Skin Disease Virus Following Experimental Infection in Cattle. Transbound. Emerg. Dis. 2008, 55, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Bowden, T.R.; Babiuk, S.L.; Parkyn, G.R.; Copps, J.S.; Boyle, D.B. Capripoxvirus Tissue Tropism and Shedding: A Quantitative Study in Experimentally Infected Sheep and Goats. Virology 2008, 371, 380–393. [Google Scholar] [CrossRef]
- Ahmed, E.M.; Eltarabilli, M.M.A.; Shahein, M.A.; Fawzy, M. Lumpy Skin Disease Outbreaks Investigation in Egyptian Cattle and Buffaloes: Serological Evidence and Molecular Characterization of Genome Termini. Comp. Immunol. Microbiol. Infect. Dis. 2021, 76, 101639. [Google Scholar] [CrossRef]
- Kononov, A.; Olga, B.; David, W.B.; Pavel, P.; Yana, P.; Svetlana, K.; Alexander, N.; Vladimir, R.; Dmitriy, L.; Alexander, S. Non-vector-borne transmission of lumpy skin disease virus. Sci. Rep. 2020, 10, 7436. [Google Scholar] [CrossRef]
- Sohier, C.; Haegeman, A.; Mostin, L.; De Leeuw, I.; Campe, W.V.; De Vleeschauwer, A.; Tuppurainen, E.S.M.; van den Berg, T.; De Regge, N.; De Clercq, K. Experimental Evidence of Mechanical Lumpy Skin Disease Virus Transmission by Stomoxys calcitrans Biting Flies and Haematopota spp. Horseflies. Sci. Rep. 2019, 9, 20076. [Google Scholar] [CrossRef] [PubMed]
- Prozesky, L.; Barnard, B.J. A Study of the Pathology of Lumpy Skin Disease in Cattle. Onderstepoort J. Vet. Res. 1982, 49, 167–175. [Google Scholar] [PubMed]
- Carn, V.M.; Kitching, R.P. The Clinical Response of Cattle Experimentally Infected with Lumpy Skin Disease (Neethling) Virus. Arch. Virol. 1995, 140, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Barnard, B.J. Antibodies Against some Viruses of Domestic Animals in Southern African Wild Animals. Onderstepoort J. Vet. Res. 1997, 64, 95–110. [Google Scholar] [PubMed]
- Agianniotaki, E.I.; Tasioudi, K.E.; Chaintoutis, S.C.; Iliadou, P.; Mangana-Vougiouka, O.; Kirtzalidou, A.; Alexandropoulos, T.; Sachpatzidis, A.; Plevraki, E.; Dovas, C.I.; et al. Lumpy Skin Disease Outbreaks in Greece during 2015-16, Implementation of Emergency Immunization and Genetic Differentiation between Field Isolates and Vaccine Virus Strains. Vet. Microbiol. 2017, 201, 78–84. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Lumpy Skin Disease: I. Data Collection and Analysis. EFSA J. 2017, 15, e04773. [Google Scholar]
- EFSA Panel on Animal Health and Welfare (AHAW); Nielsen, S.S.; Alvarez, J.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Gortazar Schmidt, C.; et al. Assessment of the Control Measures for Category A Diseases of Animal Health Law: Lumpy Skin Disease. EFSA J. 2022, 20, e07121. [Google Scholar]
- El-Nahas, E.M.; El-Habbaa, A.S.; El-bagoury, G.F.; Radwan, M.E.I. Isolation and Identification of Lumpy Skin Disease Virus from Naturally Infected Buffaloes at Kaluobia, Egypt. Global Vet. 2011, 7, 234–237. [Google Scholar]
- Sharawi, S.S.A.; Abd El-Rahim, I.H.A. The Utility of Polymerase Chain Reaction for Diagnosis of Lumpy Skin Disease in Cattle and Water Buffaloes in Egypt. Rev. Sci. Tech. 2011, 30, 821–830. [Google Scholar] [CrossRef]
- Davies, F.G. Lumpy Skin Disease, an African Capripox Virus Disease of Cattle. Br. Vet. J. 1991, 147, 489–503. [Google Scholar] [CrossRef]
- Fagbo, S.; Coetzer, J.A.W.; Venter, E.H. Seroprevalence of Rift Valley Fever and Lumpy Skin Disease in African Buffalo (Syncerus caffer) in the Kruger National Park and Hluhluwe-I Mfolozi Park, South Africa. J. S. Afr. Vet. Assoc. 2014, 85, e1–e7. [Google Scholar] [CrossRef] [PubMed]
- Pandey, N.; Hopker, A.; Prajapati, G.; Rahangdale, N.; Gore, K.; Sargison, N. Observations on Presumptive Lumpy Skin Disease in Native Cattle and Asian Water Buffaloes Around the Tiger Reserves of the Central Indian Highlands. N. Z. Vet. J. 2022, 70, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Neamat-Allah, A.N.F.; Mahmoud, E.A. Assessing the possible causes of hemolyticanemia associated with lumpy skin disease naturally infected buffaloes. Comp. Clin. Pathol. 2019, 28, 747–753. [Google Scholar] [CrossRef]
- Aerts, L.; Haegeman, A.; De Leeuw, I.; Philips, W.; Van Campe, W.; Behaeghel, I.; Mostin, L.; De Clercq, K. Detection of Clinical and Subclinical Lumpy Skin Disease using Ear Notch Testing and Skin Biopsies. Microorganisms 2021, 9, 2171. [Google Scholar] [CrossRef] [PubMed]
- World Organization for Animal Health (WOAH). Lumpy skin disease. In Terrestrial Manual; WOAH: Paris, France, 2023; Chapter 3.4.12. [Google Scholar]
- Samojlović, M.; Polaček, V.; Gurjanov, V.; Lupulović, D.; Lazić, G.; Petrović, T.; Lazić, S. Detection of Antibodies Against Lumpy Skin Disease Virus by Virus Neutralization Test and ELISA Methods. Acta Vet. 2019, 69, 47–60. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Dessouki, A.A. Abattoir-Based Survey and Histopathological Findings of Lumpy Skin Disease in Cattle at Ismailia Abattoir. IJBBB 2013, 3, 372. [Google Scholar]
- House, J.A.; Wilson, T.M.; el Nakashly, S.; Karim, I.A.; Ismail, I.; el Danaf, N.; Moussa, A.M.; Ayoub, N.N. The Isolation of Lumpy Skin Disease Virus and Bovine Herpesvirus-4 from Cattle in Egypt. J. Vet. Diagn. Investig. 1990, 2, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Attempted Mechanical Transmission of Lumpy Skin Disease Virus by Biting Insects. Med. Vet. Entomol. 2003, 17, 294–300. [Google Scholar] [CrossRef]
- Ben-Gera, J.; Klement, E.; Khinich, E.; Stram, Y.; Shpigel, N.Y. Comparison of the efficacy of Neethling lumpy skin disease virus and x10RM65 sheep-pox live attenuated vaccines for the prevention of lumpy skin disease–The results of a randomized controlled field study. Vaccine 2015, 33, 4837–4842. [Google Scholar] [CrossRef]
- Haegeman, A.; De Leeuw, I.; Mostin, L.; Van Campe, W.; Aerts, L.; Vastag, M.; De Clercq, K. An Immunoperoxidase Monolayer Assay (IPMA) for the Detection of Lumpy Skin Disease Antibodies. J. Virol. Methods 2020, 277, 113800. [Google Scholar] [CrossRef]
- Tuppurainen, E.; Alexandrov, T.; Beltrán-Alcrudo, D. Lumpy Skin Disease Field Manual—A Manual for Veterinarians; FAO Animal Production and Health Manual No. 20; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2017; 60p. [Google Scholar]
- Elhaig, M.M.; Selim, A.; Mahmoud, M.; Elhaig, M. Lumpy skin disease in cattle: Frequency of occurrence in a dairy farm and a preliminary assessment of its possible impact on Egyptian buffaloes. Onderstepoort J. Vet. Res. 2017, 84, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Bernardo, B.; Haga, I.R.; Wijesiriwardana, N.; Basu, S.; Larner, W.; Diaz, A.V.; Langlands, Z.; Denison, E.; Stoner, J.; White, M.; et al. Quantifying and Modeling the Acquisition and Retention of Lumpy Skin Disease Virus by Hematophagus Insects Reveals Clinically but Not Subclinically Affected Cattle are Promoters of Viral Transmission and Key Targets for Control of Disease Outbreaks. J. Virol. 2021, 95, e02239-20. [Google Scholar] [CrossRef] [PubMed]
- Okur-Gumusova, S.; Tamer, C.; Ozan, E.; Cavunt, A.; Kadi, H.; Muftuoglu, B.; Eisa Elhag, A.; Yazici, Z.; Albayrak, H. An Investigation of the Seroprevalence of Crimean-Congo Hemorrhagic Fever and Lumpy Skin Disease in Domesticated Water Buffaloes in Northern Turkey. Trop. Biomed. 2020, 37, 165. [Google Scholar]
- European Food Safety Authority Panel on Animal Health and Welfare. Scientific Opinion on lumpy skin disease. EFSA J. 2015, 13, 3986. [Google Scholar] [CrossRef]
- World Organization for Animal Health (WOAH). 2021 Terrestrial Animal Health Code, Chapter 11.9: Infection with Lumpy Skin Disease. 2021. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahc/current/chapitre_lsd.pdf (accessed on 3 March 2023).
- Rehman, S.U.; Hassan, F.U.; Luo, X.; Li, Z.; Liu, Q. Whole-Genome Sequencing and Characterization of Buffalo Genetic Resources: Recent Advances and Future Challenges. Animals 2021, 11, 904. [Google Scholar] [CrossRef] [PubMed]
- Iannaccone, M.; Cosenza, G.; Pauciullo, A.; Martino, G.; Capparelli, R. The SNP g.4667G>A at 3′-UTR of IFNG gene is associated with susceptibility to bovine tuberculosis in Mediterranean water buffalo (Bubalus bubalis). Anim. Genet. 2018, 49, 496–497. [Google Scholar] [CrossRef] [PubMed]
- El-Halawany, N.; Abd-El-Monsif, S.A.; Al-Tohamy Ahmed, F.M.; Hegazy, L.; Abdel-Shafy, H.; Abdel-Latif, M.A.; Ghazi, Y.A.; Neuhoff, C.; Salilew-Wondim, D.; Schellander, K. Complement component 3: Characterization and association with mastitis resistance in Egyptian water buffalo and cattle. J. Genet. 2017, 96, 65–73. [Google Scholar] [CrossRef]
- El-Halawany, N.; Shawky, A.A.; Al-Tohamy, A.F.M.; Abdel-Latif, M.A.; Abdel-Shafy, H.; Ghazi, Y.A.; Neuhoff, C.; Schellander, K. Effect of complement component 5 polymorphisms on mastitis resistance in Egyptian buffalo and cattle. Res. Vet. Sci. 2018, 119, 45–51. [Google Scholar] [CrossRef]
- Othman, O.E.; Khodary, M.G.; El-Deeb, A.H.; Hussein, H.A. Five BoLA-DRB3 genotypes detected in Egyptian buffalo infected with Foot and Mouth disease virus serotype O. J. Genet. Eng. Biotechnol. 2018, 16, 513–518. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| General Health Status | Food Intake | Nasal Discharge | Number of Nodules | Dissemination of Nodules | Lymphadenomegaly | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Normal | 0 | Normal | 0 | Normal | 0 | No Nodules | 0 | No Nodules | 0 | No Lymphadenomegaly | 0 |
| Mild Illness | 1 | Slightly Decreased | 1 | Mild | 1 | ≤10 | 1 | Localized | 1 | Localized | 1 |
| Decreased | 2 | Marked Mucous | 2 | <20 | 2 | ||||||
| Severe Illness | 2 | Does Not eat | 3 | Purulent | 3 | ≥20 | 3 | Generalized | 2 | Generalized | 2 |
| Clinical Finding | Inoculated Animals | |||||||
|---|---|---|---|---|---|---|---|---|
| Buffalo | Cattle | |||||||
| 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 * | |
| Decreased food intake | For 2 days | - | - | - | - | For 2 days | For 2 days | For 4 days |
| Nodules | ||||||||
| Number | >20 | 2 | 4 | 4 | 3 | 12 | 4 | >20 |
| Size (ø) | 0.5–5.0 cm | 0.2–0.5 cm | 0.5–2.0 cm | 0.5–2.5 cm | 0.2–2.0 cm | 0.5–3.5 cm | 2.0–6.5 cm | 2.0–7.0 cm |
| Location | Generalized | Inoculation sites | Inoculation sites | Inoculation sites | Inoculation sites | Generalized | Inoculation sites | Generalized |
| Lymphadenomegaly | Generalized | Prescapular and prefemoral | Prescapular and prefemoral | Prescapular and prefemoral | Not detected | Generalized | Generalized | Generalized |
| Oedema | ND | ND | ND | ND | ND | ND | ND | Dewlap |
| Nasal and ocular discharge | ND | ND | ND | ND | ND | ND | From mucopurulent to serous; bilateral | From mucopurulent to serous; bilateral |
| Viremia | ND | ND | ND | ND | ND | ND | From day 9 (Ct 28.54) to day 14 (Ct 38.40) | From day 5 (Ct 35.57) to day 9 (Ct 32.97) * |
| Inoculated Animals | ||||||||
|---|---|---|---|---|---|---|---|---|
| Buffalo | Cattle | |||||||
| Tissue | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| Skin | ND | ND | ND | ND | ND | ND | ND | Ct 19.63 |
| Skin nodule | ND b | a | a | ND | a | ND b | a | Ct 20.39 Ct 19.39 and 18.47 c |
| Spleen | ND | ND | ND | ND | ND | ND | ND | Ct 29.14 |
| Kidney | ND | ND | ND | ND | ND | ND | ND | Ct 29.84 |
| Lung | ND | ND | ND | ND | ND | ND | ND | Ct 29.02 |
| Liver | ND | ND | ND | ND | ND | ND | ND | Ct 30.49 |
| Bronchial LN | ND | ND | ND | ND | ND | ND | ND | Ct 29.58 |
| Inguinal LN | ND | ND | ND | ND | ND | ND | ND | Ct 24.11 |
| Mesenteric LN | ND | ND | ND | ND | ND | ND | ND | Ct 28.44 |
| Right prescapolar LN | ND | ND | ND | ND | ND | ND | Ct 35.11 | Ct 24.70 |
| Left prescapolar LN | a | a | a | a | a | a | ND | Ct 25.70 |
| Submandibolar LN | ND | ND | ND | ND | ND | ND | ND | Ct 23.63 |
| Rumen | ND | ND | ND | ND | ND | ND | ND | Ct 28.43 |
| Nasal mucosa | ND | ND | ND | ND | ND | ND | ND | Ct 18.73 d |
| Day Post Inoculation | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample | Animal | 2 | 3 | 4 | 5 | 7 | 9 | 14 | 17 | 21 | 24 | 31 | 35 | 38 |
| Nasal swab | Buffalo 3 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| Buffalo 4 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Buffalo 5 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Buffalo 6 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Buffalo 7 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Buffalo 8 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Cattle 9 | ND | ND | ND | ND | ND | ND | ND | 37.12 | 37.42 | 39.44 | 35.92 | 37.16 | ND | |
| Cattle 10 | ND | ND | ND | ND | 34.04 | 31.97 | ||||||||
| Oral swab | Buffalo 3 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| Buffalo 4 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Buffalo 5 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Buffalo 6 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Buffalo 7 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Buffalo 8 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Cattle 9 | ND | ND | ND | ND | ND | 37.45 | ND | 38.45 | 36.83 | ND | 34.78 | ND | ND | |
| Cattle 10 | ND | ND | ND | ND | 37.37 | 37.97 | ||||||||
| Conjunctival swab | Buffalo 3 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| Buffalo 4 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Buffalo 5 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Buffalo 6 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Buffalo 7 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Buffalo 8 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Cattle 9 | ND | ND | ND | ND | ND | ND | ND | ND | 36.93 | ND | ND | 38.45 | ND | |
| Cattle 10 | ND | ND | ND | ND | ND | 37.05 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Felice, E.; Pinoni, C.; Rossi, E.; Amatori, G.; Mancuso, E.; Iapaolo, F.; Taraschi, A.; Di Teodoro, G.; Di Donato, G.; Ronchi, G.F.; et al. Susceptibility of Mediterranean Buffalo (Bubalus bubalis) following Experimental Infection with Lumpy Skin Disease Virus. Viruses 2024, 16, 466. https://doi.org/10.3390/v16030466
Di Felice E, Pinoni C, Rossi E, Amatori G, Mancuso E, Iapaolo F, Taraschi A, Di Teodoro G, Di Donato G, Ronchi GF, et al. Susceptibility of Mediterranean Buffalo (Bubalus bubalis) following Experimental Infection with Lumpy Skin Disease Virus. Viruses. 2024; 16(3):466. https://doi.org/10.3390/v16030466
Chicago/Turabian StyleDi Felice, Elisabetta, Chiara Pinoni, Emanuela Rossi, Giorgia Amatori, Elisa Mancuso, Federica Iapaolo, Angela Taraschi, Giovanni Di Teodoro, Guido Di Donato, Gaetano Federico Ronchi, and et al. 2024. "Susceptibility of Mediterranean Buffalo (Bubalus bubalis) following Experimental Infection with Lumpy Skin Disease Virus" Viruses 16, no. 3: 466. https://doi.org/10.3390/v16030466