1. Introduction
Hepatitis E virus (HEV) is a major cause of acute hepatitis in humans worldwide [
1]. Currently, there are eight distinct HEV genotypes (under genus
Paslahepevirus) reported, but genotypes 1–4 (GT 1–4) were identified as the cause for the most human clinical cases [
2,
3]. GT1 and 2 are obligate human pathogens transmitted through contaminated drinking water, causing large-scale epidemic outbreaks of acute hepatitis in flood-prone regions of the world with poor sanitary infrastructure [
4]. In contrast, GT 3 and 4 are zoonoses associated with sporadic cases of acute and chronic hepatitis E worldwide, which are mainly transmitted to humans through the consumption of undercooked organ meat (primarily pig products) and, rarely, through blood transfusions [
5]. Despite a diverse host range, distinct epidemiological features, and genetic diversity, GT1–4 possess a single serotype [
4]; therefore, well-developed serological assays can be useful for HEV diagnosis.
Anti-HEV IgM assays are used to diagnose acute hepatitis E in immunocompetent patients. However, the performance of these assays was found to be variable, and discordance between assays was noted [
6,
7]. Moreover, the IgM assays are not reliable for the diagnosis of chronic hepatitis E in immunosuppressed patients [
8]. Therefore, the European Association for the Study of the Liver (EASL) recommends using a combination of anti-HEV IgM and HEV NAAT (nucleic acid amplification technology) to diagnose acute hepatitis E cases and the HEV NAAT only to diagnose chronic hepatitis E cases [
8]. NAAT assays are also used for screening HEV infections in asymptomatic blood donors in European countries and Japan [
9,
10]. However, NAAT is not widely used in resource-limited countries for HEV diagnosis and screening because of the requirement for expensive specialized equipment and trained staff [
11]. A qualitative HEV antigen (ORF2) serologic assay is commercially available for research use, and the diagnostic utility of this assay was investigated in few studies involving small cohorts of patients hospitalized with acute hepatitis E [
12,
13,
14,
15,
16,
17].
The qualitative ORF2 antigen assay detected acute hepatitis E cases with a sensitivity of 91–100% [
13,
15]. The assay was able to discriminate acute and chronic hepatitis E cases of GT3 infection with a sensitivity of 95% and specificity of 88% [
16]. The ORF2 antigen assay also discriminated self-limiting GT1 infections from fulminant hepatic failure progression with 100% sensitivity and 89% specificity [
17]. The positive and negative predictive values of the ORF2 antigen ELISA for the clinical diagnosis of acute GT1 outbreaks were found to be 60–81% and 88–100%, respectively [
12]. HEV ORF2 antigen may be a useful biomarker of active HEV infection in cases where prolonged IgM response is observed after the resolution of acute hepatitis E [
14]. A logistic regression model integrating three serological markers of HEV infection, ORF2 antigen, anti-HEV IgM, and ALT, improved the sensitivity (89.5%) and specificity (95.2%) of the clinical diagnosis of ongoing HEV infections compared with an anti-HEV IgM assay or HEV antigen assay [
18]. However, the utility of ORF2 antigen in hepatitis E diagnosis is yet to be determined by developing new, improved assays and conducting studies involving large patient cohorts.
The HEV ORF2 protein was detected at high levels in the sera of human patients as well as rhesus macaques experimentally infected with HEV [
19,
20]. Three distinct ORF2 isoforms were identified in the sera of HEV-infected persons: (1) secreted glycosylated ORF2 (ORF2g), (2) a cleaved ORF2 (ORF2c), and (3) the capsid-associated ORF2 (ORF2i) [
19,
20,
21]. The former two isoforms are the major antigens in blood and primary targets of HEV antigen ELISA [
19]. In this study, we developed and optimized a quantitative sandwich ELISA protocol using two novel monoclonal antibodies specific to HEV ORF2 antigen and the biotin–streptavidin technology to improve the sensitivity of detection. The assay was tested for quantifying ORF2 antigen in a cell culture medium of human hepatocytes, serum from HEV-infected gerbils, and ORF2-spiked human plasma.
2. Materials and Methods
2.1. Human Plasma Specimens
Whole Blood was collected from volunteer donors in citrate phosphate dextrose at the NIH Division of Transfusion Medicine under approval from the NIH and FDA Institutional Review Boards (IRBs). Plasma was prepared from Whole Blood and immediately stored at −80 °C until use.
Plasma derived from deidentified blood donor samples reactive to markers of HBV or HCV infection was purchased from the American Red Cross Infectious Disease Repository (Gaithersburg, MD, USA). Plasma specimens reactive to HEV were purchased from SeraCare/LGC Diagnostics, Milford, MA, USA (AccuSet HEV Performance Panel, 0820-0503/Batch #10640609). The plasma samples were aliquoted and stored at −80 °C until use. Frozen samples were thawed and diluted directly in blocking buffer for sandwich ELISA analysis.
2.2. Animal Studies
The animal studies were conducted in strict adherence to the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The animal study protocols were reviewed and approved by FDA White Oak Animal Care and Use Committee (ACUC), Silver Spring, USA, protocol numbers 2019-12 and 2020-02.
2.3. Preparation of Recombinant ORF2 Antigens
The DNA sequence encoding ORF2 amino acids 422–637 (US-2 strain, GenBank Accession No: AAD15816.1, NCBI (Bethesda, MD, USA)) was codon optimized and chemically synthesized (IDT technologies, Coralville, IA, USA). The ORF2 DNA was inserted into the pET28a+ expression vector (Novagen, EMD Millipore, Burlington, MA, USA). The resulting plasmid was used to transform BL21 (DE3) E. coli. ORF2 expression was induced by IPTG induction. Cells were lysed, and ORF2 was purified from lysates by His tag affinity chromatography using HisPur Ni-NTA superflow agarose resin (Thermofisher Scientific, Waltham, MA, USA). The purified ORF2 protein was desalted and exchanged with PBS by ultrafiltration or dialysis. The protein concentration was measured by Bradford assay, and the protein was aliquoted for single use and stored at −80 °C.
2.4. Preparation of Mouse Monoclonal Antibodies against the ORF2 Antigen
Six-week-old female Balb/c mice were immunized with 50 µg ORF2 antigen emulsified with Titermax gold adjuvant at 1:1. Mice received booster immunizations at 4 and 6 weeks with 25 µg antigen emulsified with Freund’s incomplete adjuvant at 1:1. Antibody responses in mice were assessed by an indirect ELISA at 1 week after each booster immunization. At 10 weeks, mice were injected intravenously with 5–10 µg antigen. Three days later, mice were euthanized, and spleens were harvested for splenocyte preparation. Splenocytes were fused with Sp2/0-Ag14 myeloma cells (ATCC) to generate hybridomas. A panel of 1000 hybridoma clones was generated using ClonaCell-HY Hybridoma kit (Stem Cell technologies, Cambridge, MA, USA). Hybridoma culture supernatants were screened by an indirect ELISA for ORF2 antibodies. Five hybridoma clones positive for ORF2 antibodies were identified, cultured in bulk, and antibodies were purified from the culture supernatants using PureProteome Protein G magnetic beads (Millipore sigma, Burlington, MA, USA) according to the manufacturer’s instructions. Only four ORF2 antibodies were tested for antigen capture and detection. The antibodies for antigen capture were stored at 4 °C in Tris-glycine buffer containing 0.02% sodium azide. The antibodies used in detection were desalted, buffer exchanged with PBS, and conjugated with biotin using Lightning-Link Biotinylation Kit (Type A) (Abcam, Waltham, MA, USA) according to the manufacturer’s instructions. The biotin-conjugated antibodies for detection were stored at 4 °C in PBS mixed with sodium azide at 0.02% final concentration.
2.5. Sandwich ELISA Protocol Development
A 96-well microplate (Nunc Maxisorp, Thermofisher Scientific, Waltham, MA, USA) was coated with capture antibody (200 ng/100 μL/well, diluted in ELISA coating buffer (Bio-Rad, Hercules, CA, USA)) overnight at RT. After 3 washings with PBS-Tween 20 (0.05%) (PBS-T), blocking buffer (5% non-fat milk, 150 μL/well) was added and incubated for 1 h at 37 °C. The blocking buffer was decanted, and serially diluted recombinant ORF2 p216 antigen (0.1, 1, 10, 100, and 1000 ng/mL; 100 μL per well) was added and incubated overnight at 4 °C. Duplicate wells were prepared for each dilution. After 3 washings with PBS-T, biotin-conjugated detection antibody (1 μg/mL, 100 μL/well) was added and incubated for 1 h at 37 °C. After 5 washings with PBS-T, Streptavidin–HRP (1:20,000 dilution, 100 μL/well, Thermofisher Scientific, Waltham, MA, USA) was added and incubated for 1 h at 37 °C. After 5 washes with PBS-T, SureBlue TMB microwell peroxidase substrate (100 μL/well, KPL, SeraCare/LGC Diagnostics, Milford, MA, USA) was added and incubated for 5–30 min at RT in the dark. The reaction was stopped with 1N HCl (100 μL/well). Absorbance was measured at 450 nm in a microplate reader (Biotek Synergy neo2, Agilent, Santa Clara, CA, USA).
2.6. Screening of Matched Antibody Pairs
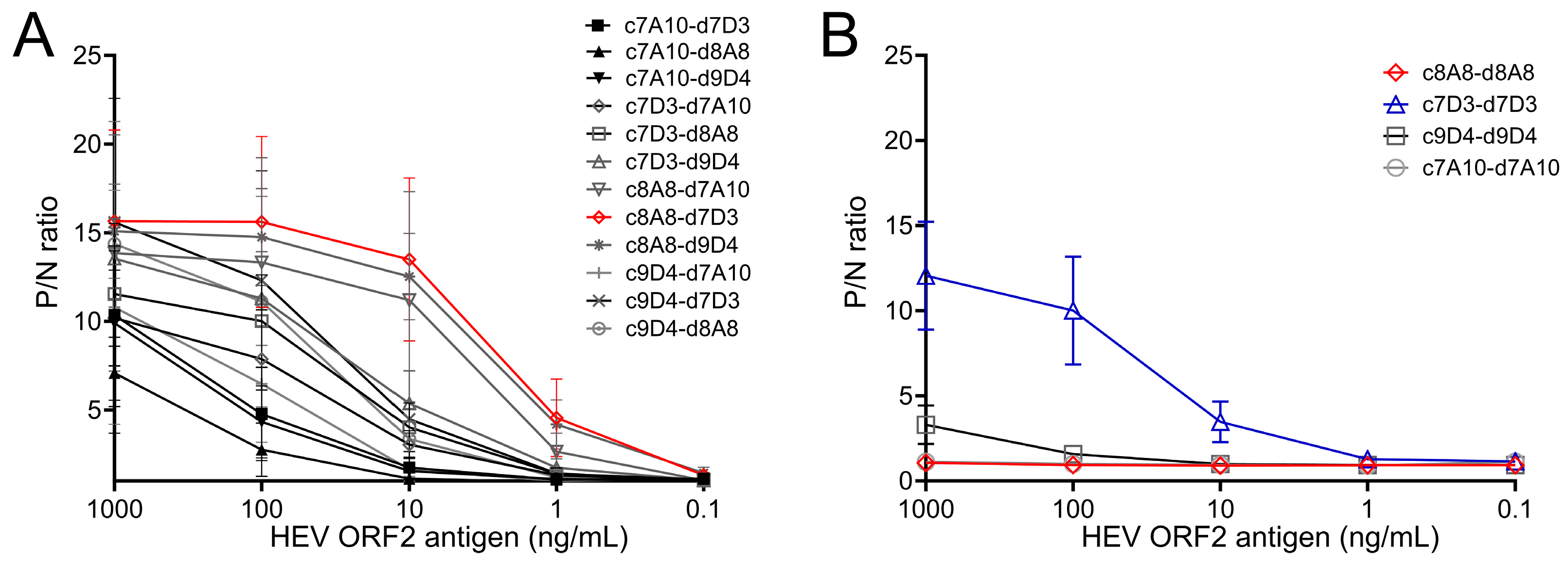
The 4 capture antibodies (c7A10, c7D3, c8A8, c9D4) were paired with 4 detection antibodies (d7A10, d7D3, d8A8, d9D4) and screened in the sandwich ELISA following the protocol indicated above. Serially diluted recombinant ORF2 p216 antigen (0.1, 1, 10, 100, and 1000 ng/mL; 100 μL per well) and blocking buffer without antigen served as positive samples and negative control in the assay, respectively. The antibody pair showing largest ratios of OD450 values between the positive samples and negative control was selected as the optimal matched antibody pair for the sandwich ELISA.
2.7. Sandwich ELISA Protocol Optimization
The optimal antibody concentrations for both capture and detection antibodies were determined by checkerboard titration. Different concentrations of the capture antibody (2, 4, 6, 8, 10 µg/mL) and detection antibody (0.5, 1, 2 µg/mL) were tested in the sandwich ELISA. Serially diluted recombinant ORF2 p216 antigen (0.1, 1, 10, 100, and 1000 ng/mL; 100 μL per well) and blocking buffer without antigen served as positive samples and negative control in the assay, respectively. The antibody concentrations showing the largest difference in OD450 values between the positive samples and negative control were selected. Similarly, the different concentrations of Streptavidin–HRP (1:4000, 1:10,000, 1:20,000) were tested in the sandwich ELISA, and the concentration showing the largest difference in OD450 values between positive samples and negative control was selected.
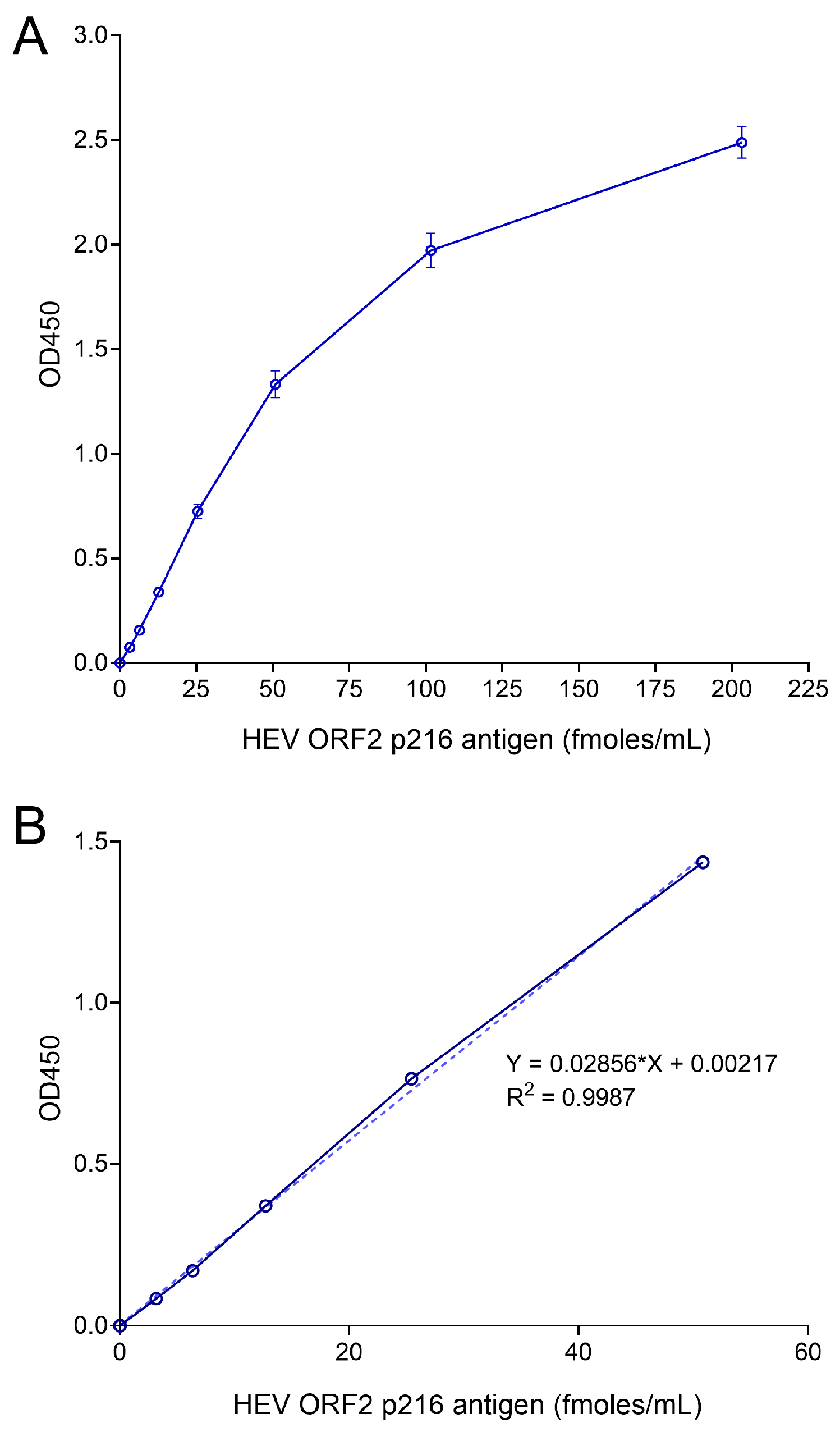
2.8. Generation of Standard Curves
The optimized sandwich ELISA protocol was followed to generate standard curves for ORF2 quantification. Recombinant US-2 ORF2 antigen (prepared in-house) was serially diluted 2-fold with blocking buffer (10, 5, 2.5, 1.25, 0.625, 0.312, 0.156, 0 ng/mL equivalent to 203.2, 101.6, 50.8, 25.4, 12.7, 6.35, 3.17, and 0 fmoles/mL, respectively). The standard curves were calculated with ORF2 antigen concentration on the X-axis and OD450 values on the Y-axis. The ORF2 antigen was quantified by interpolating from standard curve and expressed as fmoles/mL. The limit of detection (LOD) of ORF2 antigen in the ELISA was calculated using 34 negative serum samples obtained from 18 gerbils or 10 negative plasma samples obtained from 10 blood donors by the following formula: Mean Optical Density (OD) or Mean quantity (fmoles/mL) + (3× Standard Deviation (SD)). For gerbil serum samples, the calculated LOD is 6.2 fmoles/mL. For human plasma samples, the calculated LOD is 6.3 fmoles/mL or 0.011 (OD).
2.9. Cross-Reactivity Analysis
The cross-reactivity of the capture and detection antibodies to the ORF2 antigens of Paslahepevirus balayani (formerly HEV-A) GT1 (Genbank: AF444002), GT3 (Genbank: AF060669), GT4 (Genbank: HQ634346), and Rocahepevirus ratti (formerly HEV-C1 or rat HEV; Genbank: MK050105) was measured by the optimized sandwich ELISA. The ORF2 antigens were expressed in bacteria as described above. Two-fold serially diluted recombinant ORF2 p216 antigen (80–0.078 ng/mL; 100 μL per well) were tested, and blocking buffer without antigen served as negative control in the assay. The ratios of OD450 values between positive samples and negative control were calculated for different ORF2 antigens.
2.10. HEV RNA Preparation and Transfection in Huh7 Cells
In vitro, transcribed HEV Kernow C1 p6 RNAs or p6 GND (replication-incompetent) RNAs were prepared and electroporated into Huh7 S10-3 cells, as previously described [
22]. Capped and uncapped HEV Kernow C1 p6 RNAs were prepared with or without ARCA (Anti-Reverse Cap Analog, Thermofisher Scientific, Waltham, MA, USA), respectively. Capped p6 GND RNAs were prepared with ARCA to serve as negative control. The Huh7 culture medium was collected and replaced with an equivalent volume of fresh medium every 2 days. The harvested culture medium was centrifuged at 500×
g for 5 min to remove cell debris and stored at −80 °C until analysis. At the time of sandwich ELISA analysis, Huh7 culture supernatant was thawed and diluted up to 1:400 in the blocking buffer. Culture supernatant from capped p6 GND RNA transfected cells (at all time points) and from capped p6 RNA transfected cells (at 0 days post-transfection) were confirmed negative for ORF2 antigen at the lowest dilution 1:5 in the sandwich ELISA.
2.11. HEV Infection in Human Hepatocytes
Human hepatocytes (HH) isolated from chimeric mice with humanized livers were purchased from Phoenix Bio (Edmonton, AB, Canada), cultured on 24 well plates, and maintained according to the manufacturer’s instructions. HH cultures were infected with the genotype 3 HEV Kernow C1 p6 (Genbank JQ679013) at a MOI (multiplicity of infection) of 0.006. The MOI is calculated using focus forming units (FFU)/mL of the virus stock. The cell culture medium was collected and replaced with fresh medium every 1–4 days. The culture medium was subjected to centrifugation at 500× g for 5 min to remove cell debris and stored at −80 °C until analysis. HH culture supernatant was diluted up to 1:25 in the blocking buffer for sandwich ELISA analysis. Mock-infected HH culture supernatants and HEV-infected HH culture supernatants at 0 days post-infection were confirmed negative for ORF2 antigen at the lowest dilution of 1:5 in the sandwich ELISA.
2.12. HEV RT-qPCR
RNA was isolated from serum and cell culture supernatant using the QiaAmp Viral RNA mini kit (Qiagen) according to the manufacturer’s instructions. Frozen samples were thawed and adjusted to 140 µL with RNase-free water before RNA isolation. RNA was used fresh in RT-qPCR reactions or stored at −80 °C until use. Reverse Transcription, Quantitative Polymerase Chain Reaction (RT-qPCR) for HEV was based upon a previously published method [
23] with modifications to increase sensitivity [
24]. A standard curve was generated using a HEV RNA standard (full-length Kernow C1 p6; Genbank JQ679013) serially diluted 10-fold with nuclease-free water. “No template control” (NTC) was used as negative control and HEV RNA standard was used as positive control in each RT-qPCR plate. HEV RNA copies were calculated by interpolating from the standard curve and expressed as copies per mL.
2.13. Quantification of ORF2 Antigen in Gerbil Serum Samples
Clotted gerbil blood specimens were collected during a previously reported animal experiment [
25]. Serum samples were prepared by centrifugation at 1500×
g for 10 min at RT. Aliquots of serum samples were prepared for single use and stored at −80 °C. Serum samples were diluted up to 1:6400 in the blocking buffer for sandwich ELISA analysis. The serum samples from negative control gerbils were confirmed negative for ORF2 antigen at the lowest dilution of 1:10 in the sandwich ELISA.
2.14. Preparation of ORF2 Antigen-Spiked Human Plasma Samples
Clotted blood samples were obtained from 4 different human donors through NIH blood bank. Fresh plasma was prepared by centrifugation at 1500× g for 10 min at 4 °C. Plasma was spiked with culture medium from HEV-infected human hepatocyte cultures containing ORF2 antigen (414–608 fmoles/mL) at different volume ratios (1:2, 1:4, 1:8, 1:16, 1:32, 1:64). The spiked plasma was mixed thoroughly, aliquoted for single use and stored at −80 °C until further use. Spiked plasma samples were thawed and diluted 1:10 in the blocking buffer for sandwich ELISA analysis. Non-spiked plasma samples served as negative controls in the assay.
4. Discussion
We developed and optimized a quantitative sandwich ELISA for HEV ORF2 antigen using two novel monoclonal antibodies and biotin–streptavidin technology. Previous reports showed that HEV ORF2 antigen was detected in the blood of HEV-infected patients and blood donors [
19,
26] and experimental animals infected with HEV [
20,
27]. In experimentally infected monkeys, the ORF2 antigen was readily detected during the early phase of acute HEV infection before IgM antibodies appeared [
27]. Similarly, ORF2 antigen was detected at high levels in chronic HEV patients when IgM/IgG antibodies were minimal or absent [
16]. Immunological assays detecting viral antigens were used for the diagnosis of viral infections in clinical settings with limited resources [
28]. Currently, there are no quantitative assays available for HEV ORF2 antigen. There is one qualitative sandwich ELISA (Wantai Biological Pharmacy, Beijing) available for HEV ORF2 antigen detection in clinical samples, which is being repurposed for the quantitation of ORF2 protein in an HEV-infected culture medium and experimental rabbit model using p239 virus-like particles as a standard [
20,
29]. Our sandwich ELISA is optimized for the quantitation of HEV ORF2 protein in different matrix types, including gerbil serum, human plasma, and the cell culture medium of HEV-infected human hepatocytes or hepatoma cells, utilizing the recombinant ORF2 p216 antigen (E2s dimer) as the standard. The assay showed significant cross-reactivity to ORF2 from the genotype 1 and 4 strains. It successfully detected low levels of ORF2 antigen secreted in the culture medium of human hepatocytes infected with the HEV GT3 strain or Huh7 cells transfected with HEV GT3 RNA. The assay was used to assess and compare the magnitude and duration of ORF2 antigen detection in the serum of gerbils experimentally infected with HEV GT3 with or without immunosuppressive drug treatment. Our sandwich ELISA detected at least 359 pg/mL (equivalent to 7.3 fmoles/mL) of HEV ORF2 antigen in human plasma samples spiked with HEV GT3 ORF2 from a culture medium of infected human hepatocytes. The assay successfully detected the HEV ORF2 antigen in human plasma samples positive for HEV RNA but negative for anti-HEV antibodies. The assay was specific for HEV antigen because donor plasma samples positive for other hepatitis viruses (HBV and HCV) were non-reactive in the assay.
A previous report suggested polyclonal antibodies are not suitable for capturing the HEV ORF2 antigen, while monoclonal antibodies are efficient in capture and detection of ORF2 antigen in a sandwich immunoassay format [
27]. Moreover, indirect biotin amplification technology improved the antigen detection efficiency necessary for the analysis of clinical samples [
27]. We developed a panel of mouse monoclonal antibodies specific to an HEV GT3 (US-2) ORF2, targeting E2s dimer [
30] and identified 8A8 mAb as the best antibody to capture ORF2 antigen in the solid phase and 7D3 as the best detection antibody when using the biotin–streptavidin complex. When 8A8 mAb was both the capture and detection Ab in the competition experiments, a negative result was obtained in all Ab concentrations tested (up to 1000 ng/mL). This phenomenon is more commonly observed in high-affinity antibodies specific to the ORF2 E2s dimer [
31,
32]. On the other hand, the 7D3 antibody yielded a positive result when it was both the capture and detection Ab, indicating that two 7D3 molecules can separately bind to two epitopes on the ORF2 dimer in a noncompetitive manner. Few E2s-specific antibodies nonblocking to themselves were previously reported [
32].
The HEV ORF2 protein has three isoforms differing in molecular weight, protein structure, and glycosylation pattern [
20,
21]. Both capture and detection antibodies were developed specific to the protruding domain (P domain) [
33], which is present in all three isoforms of the ORF2 protein. Nevertheless, the secreted isoform of ORF2 (ORF2S) and its truncated versions are the exclusive targets (more than 99.9%) of the ORF2 antigen ELISA in the hepatocyte culture medium, in the patient sera and urine samples [
20,
34,
35]. Consistent with this, our sandwich ELISA detected large amounts of native ORF2S in the culture medium of human hepatocytes infected with HEV genotype 3 (Kernow C1 p6) and of Huh7 cells transfected with capped HEV RNA. Uncapped HEV RNA can initiate HEV replication, although inefficiently, and secrete low amounts of ORF2 antigen into culture supernatants [
36]. Consistent with this, our assay detected low levels of ORF2 antigen secreted from Huh7 cells transfected with uncapped HEV Kernow C1 p6 RNA.
A previous study identified eight distinct groups of ORF2 variants in HEV patients showing continued viral infection despite ribavirin therapy [
37]. These ORF2 variants possess single amino acid substitutions in the region between amino acid positions 25 to 324. The monoclonal antibodies 8A8 and 7D3 bind to domains outside this region (position 422–637); therefore, our sandwich ELISA is expected to detect these natural ORF2 variants in patients’ plasma. The antigenic target of our assay has a highly conserved glycosylation site at N562, and only six out of 261 sequences in GenBank showed a mutation (D562) at this site [
38], which may affect the antibody binding to the antigen in our assay. Indeed, our assay successfully detected non-glycosylated forms of ORF2 p216 antigen produced in bacteria, suggesting N-glycan at position 562 is dispensable for binding of both 8A8 (coating) and 7D3 (detection) antibodies to the antigen in our assay.
The HEV ORF2 antigen was detected in the blood of both acute and chronic HEV patients, and the HEV chronicity is associated with long-term immunosuppressive drug treatment [
16]. In the absence of significant antibody responses, HEV ORF2 was readily detectable in blood during the initial phase of acute HEV infection in experimentally infected monkeys [
27]. Consistent with this, gerbils showed significant levels of ORF2 antigen in serum in the first 2 weeks after HEV infection when the antibody response was absent or minimal [
25]. However, the ORF2 antigen was not detectable from 3 wpi when most serum samples turned positive for HEV IgM and IgG [
25]. This is consistent with the report that ORF2 antigen was not detected in the majority of serum samples of acute HEV patients positive for HEV IgM and IgG [
27]. Moreover, ORF2 protein was found at persistently high levels in the serum of immunosuppressed gerbils, as also observed in solid organ transplant recipients during acute HEV infection that eventually progressed to chronicity [
39]. We found that ORF2 antigen levels were strongly correlated with HEV RNA levels in gerbil serum, similar to that reported in acute HEV-infected patients [
12,
17].
The sensitivity of ORF2 antigen detection is lower compared to HEV RNA detection in clinical samples [
16,
40]. Recent studies showed that urine samples are more suitable for sensitive detection of ORF2 antigen as compared to serum [
35] but not suitable for discriminating acute and chronic HEV patients [
39]. Our sandwich ELISA was developed specifically for the ORF2 E2s antigen and is, therefore, predicted to detect the ORF2 E2s-like antigen (amino acids—459–606) excreted in urine samples [
35]. Further studies are warranted to evaluate the utility of our sandwich ELISA in detecting the HEV ORF2 antigen in urine samples.
To assess the clinical sensitivity and specificity of the sandwich ELISA, the assay needs to be evaluated with known HEV-positive and -negative clinical samples [
15]. Because HEV-positive clinical samples are rare and difficult to obtain in North America, we analyzed the analytical sensitivity of the assay with human plasma samples spiked with different concentrations of HEV GT3 ORF2 from infected human hepatocyte culture medium. The HEV ORF2 antigen was detected efficiently close to the limit of detection of the assay in spiked human plasma samples, suggesting the assay may be suitable to test human plasma samples from HEV GT3-infected patients containing low amounts of the ORF2 antigen. When tested with a commercial HEV panel comprising human plasma samples positive for HEV RNA, our ELISA detected the ORF2 antigen only in samples negative for anti-HEV antibodies, similar to the observation in acute HEV patients [
27]. Further, the assay was found to be specific to the HEV ORF2 antigen as plasma samples positive for other hepatitis viruses, such as HBV and HCV, as well as negative control plasma samples were non-reactive in our ELISA.
In conclusion, we developed and optimized a novel sandwich ELISA to detect and quantify the HEV ORF2 protein in different sample matrix types utilizing an ORF2 standard. Our study demonstrated the ORF2 antigen ELISA may be used to monitor HEV infection in vitro in a human hepatocyte culture medium, assess the magnitude and duration of acute and chronic HEV infection in a gerbil model, and detect active HEV infection in human plasma samples from HEV-infected patients negative for anti-HEV IgM or IgG. The sandwich ELISA detected ORF2 antigen produced in two different species, human cells and gerbils, using a secondary detection system (Streptavidin–HRP) that is suitable for samples from other species as well. Therefore, our sandwich ELISA is expected to detect the HEV antigen in serum and plasma samples from other animal models of the HEV infection, such as rhesus macaque, pigs, rabbits, and rats. Additional studies are needed to assess the clinical utility of the ORF2 antigen ELISA in the screening and diagnosis of HEV infections in humans.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





