
Sequential Immunizations with Influenza Neuraminidase Protein Followed by Peptide Nanoclusters Induce Heterologous Protection
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Lines
2.3. Expression and Purification of Recombinant rtNA Proteins
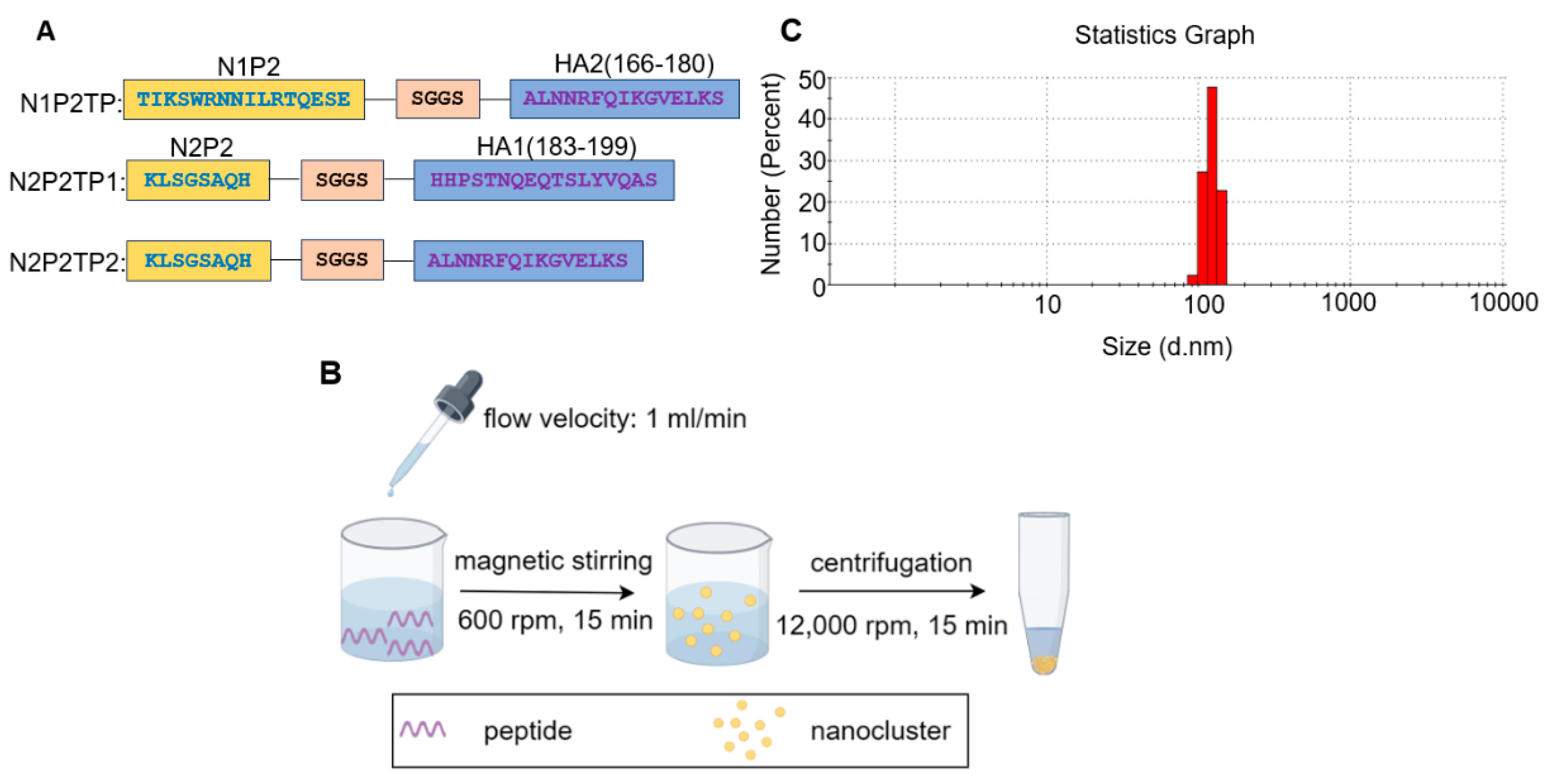
2.4. Preparation of Polypeptide Nanoclusters
2.5. Immunization and Viral Challenge Experiments
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Cell-Based ELISA
2.8. Statistical Analysis
3. Results
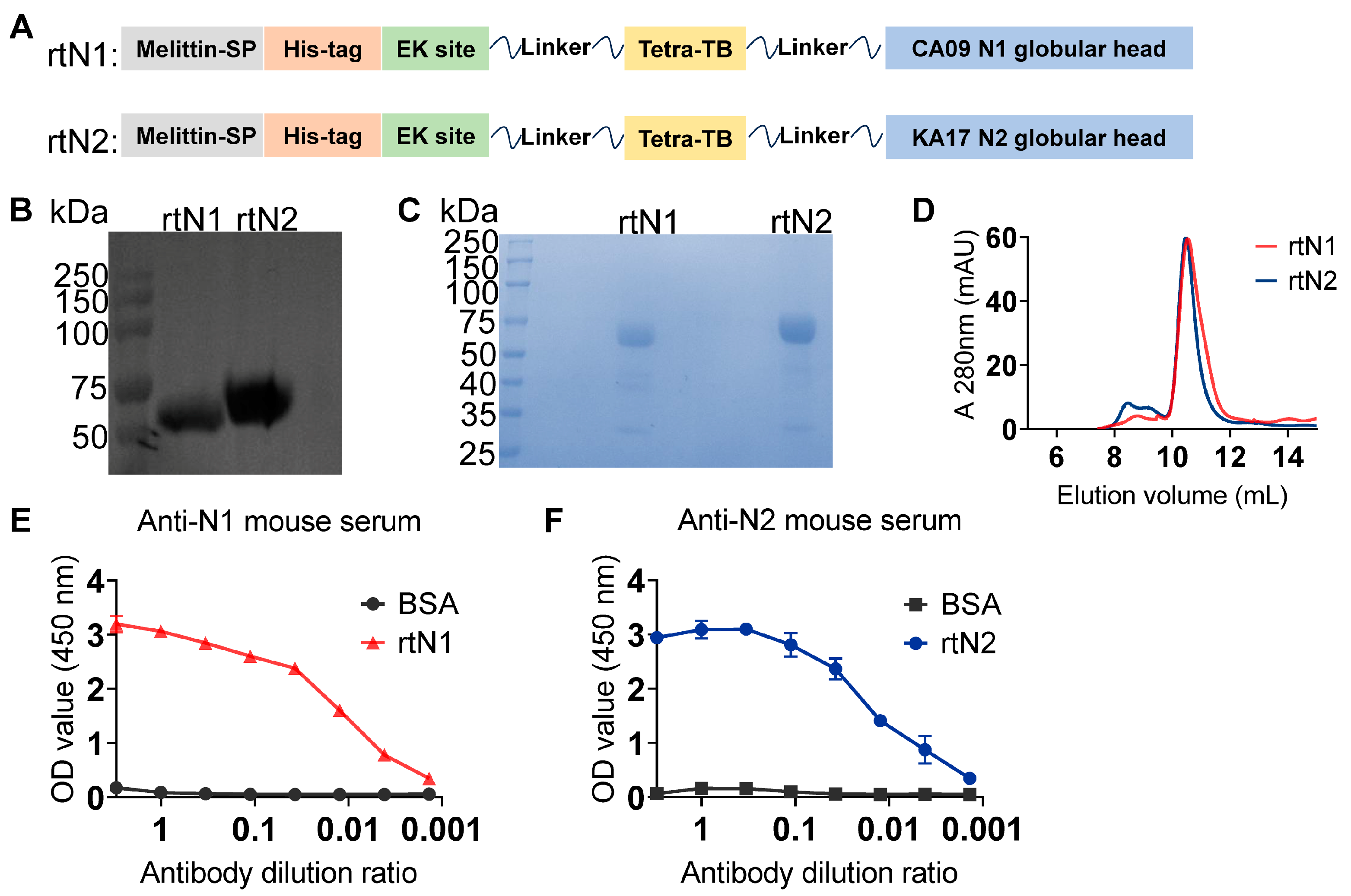
3.1. Preparation and Characterization of Recombinant NA Proteins and Peptide Nanoclusters
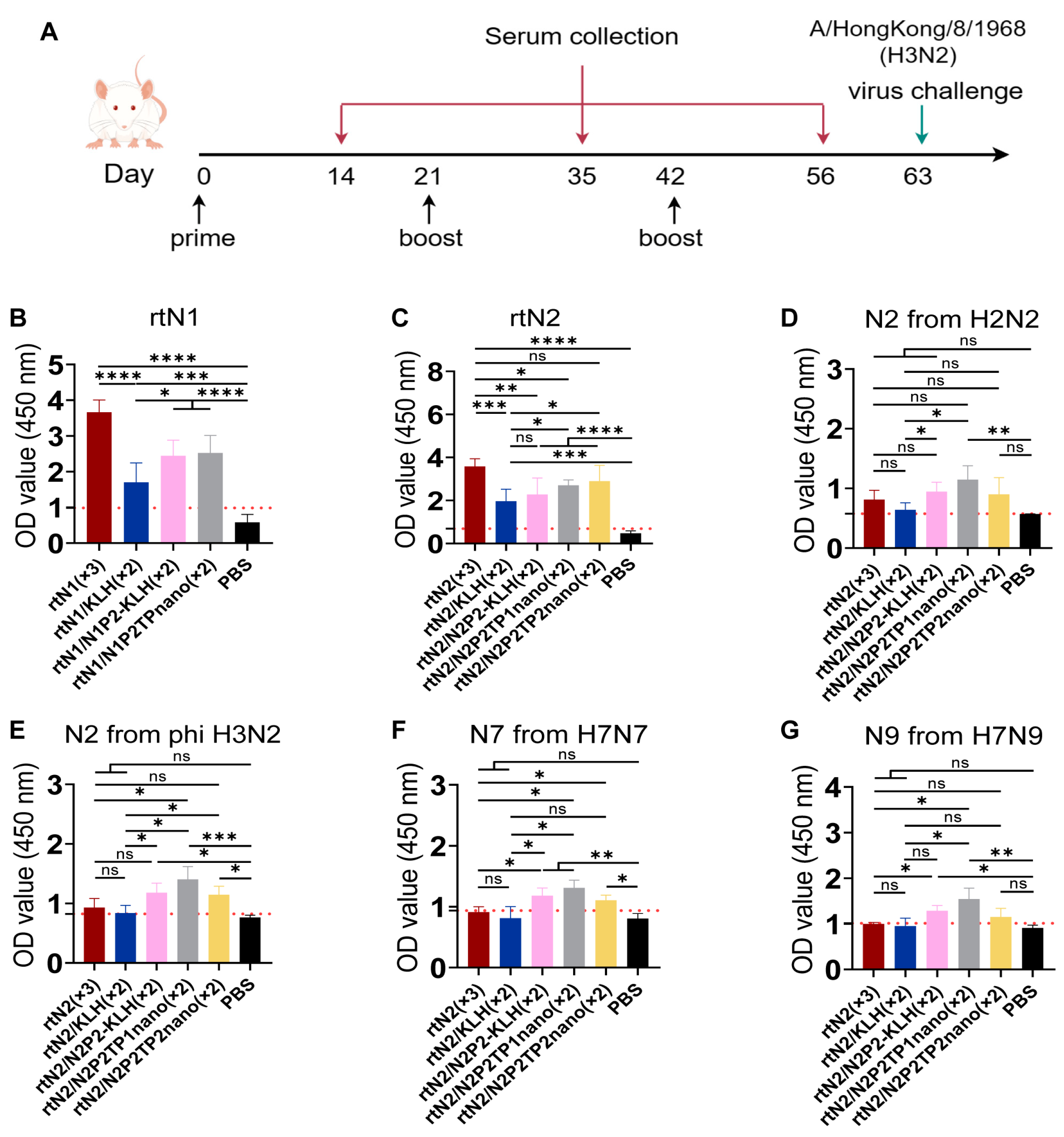
3.2. Immunizations with Peptide Nanoclusters Enhanced NA-Specific Cross-Reactive Immune Response
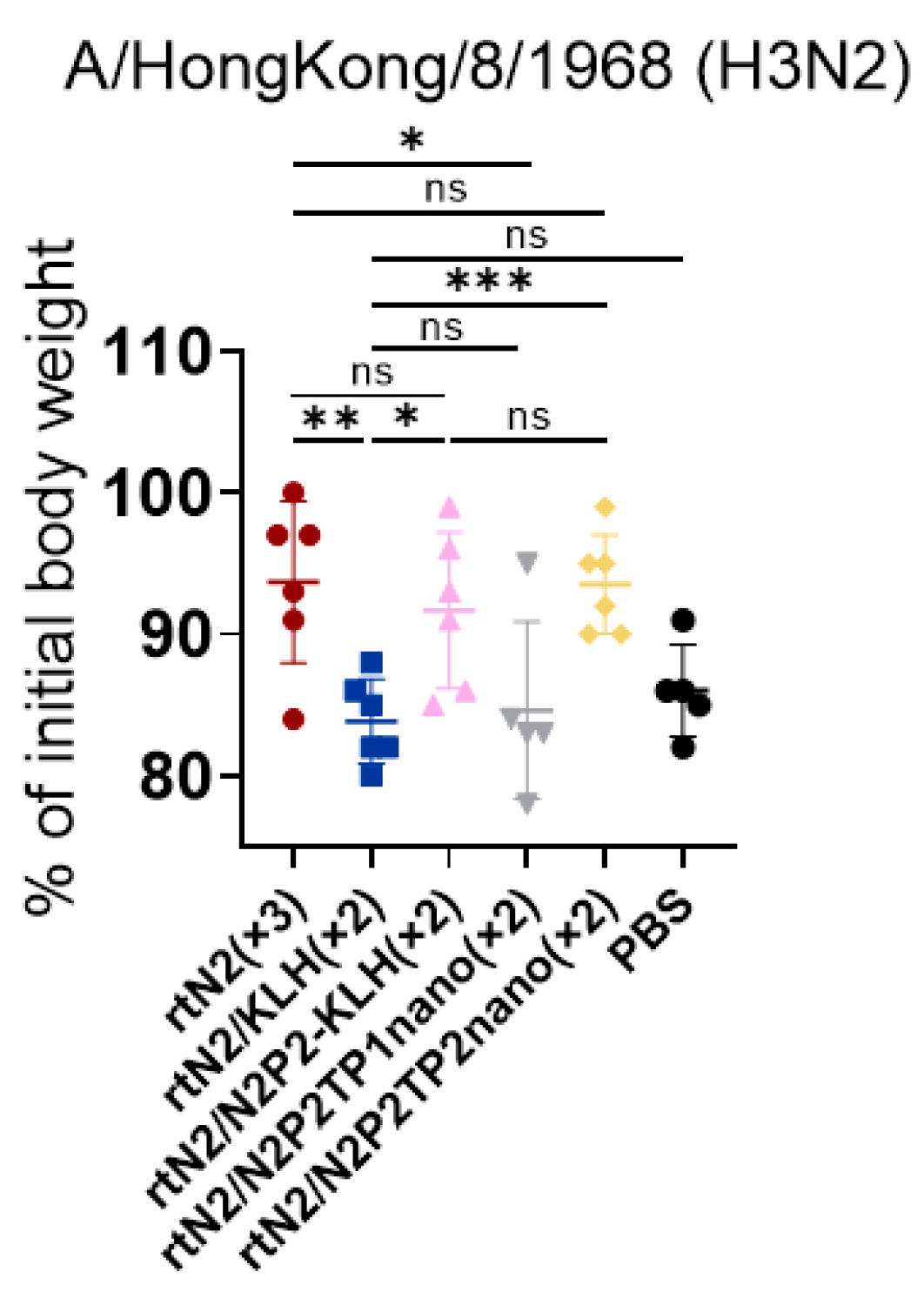
3.3. Peptide Nanoclusters Elicited Cross-Protection against Heterologous H3N2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Eichelberger, M.C.; Monto, A.S. Neuraminidase, the Forgotten Surface Antigen, Emerges as an Influenza Vaccine Target for Broadened Protection. J. Infect. Dis. 2019, 219 (Suppl. S1), S75–S80. [Google Scholar] [CrossRef] [PubMed]
- Eichelberger, M.C.; Morens, D.M.; Taubenberger, J.K. Neuraminidase as an influenza vaccine antigen: A low hanging fruit, ready for picking to improve vaccine effectiveness. Curr. Opin. Immunol. 2018, 53, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Impagliazzo, A.; Milder, F.; Kuipers, H.; Wagner, M.V.; Zhu, X.; Hoffman, R.M.; van Meersbergen, R.; Huizingh, J.; Wanningen, P.; Verspuij, J.; et al. A stable trimeric influenza hemagglutinin stem as a broadly protective immunogen. Science 2015, 349, 1301–1306. [Google Scholar] [PubMed]
- Stadlbauer, D.; Zhu, X.; McMahon, M.; Turner, J.S.; Wohlbold, T.J.; Schmitz, A.J.; Strohmeier, S.; Yu, W.; Nachbagauer, R.; Mudd, P.A.; et al. Broadly protective human antibodies that target the active site of influenza virus neuraminidase. Science 2019, 366, 499–504. [Google Scholar] [CrossRef]
- Madsen, A.; Dai, Y.N.; McMahon, M.; Schmitz, A.J.; Turner, J.S.; Tan, J.; Lei, T.; Alsoussi, W.B.; Strohmeier, S.; Amor, M.; et al. Human Antibodies Targeting Influenza B Virus Neuraminidase Active Site Are Broadly Protective. Immunity 2020, 53, 852–863 e7. [Google Scholar] [CrossRef]
- Johansson, B.E.; Moran, T.M.; Kilbourne, E.D. Antigen-presenting B cells and helper T cells cooperatively mediate intravirionic antigenic competition between influenza A virus surface glycoproteins. Proc. Natl. Acad. Sci. USA 1987, 84, 6869–6873. [Google Scholar] [CrossRef] [PubMed]
- Johansson, B.E.; Kilbourne, E.D. Dissociation of influenza virus hemagglutinin and neuraminidase eliminates their intravirionic antigenic competition. J. Virol. 1993, 67, 5721–5723. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.J.; Liu, C.C.; Zhong, X.Q.; Wu, X.; Zhang, H.H.; Lu, S.W.; Shen, Z.L.; Song, W.W.; Zhao, S.L.; Peng, Y.S.; et al. Boost immunizations with NA-derived peptide conjugates achieve induction of NA inhibition antibodies and heterologous influenza protections. Cell Rep. 2023, 42, 112766. [Google Scholar] [CrossRef]
- Choi, A.; Bouzya, B.; Cortes Franco, K.D.; Stadlbauer, D.; Rajabhathor, A.; Rouxel, R.N.; Mainil, R.; Van der Wielen, M.; Palese, P.; Garcia-Sastre, A.; et al. Chimeric Hemagglutinin-Based Influenza Virus Vaccines Induce Protective Stalk-Specific Humoral Immunity and Cellular Responses in Mice. Immunohorizons 2019, 3, 133–148. [Google Scholar] [CrossRef]
- Nachbagauer, R.; Feser, J.; Naficy, A.; Bernstein, D.I.; Guptill, J.; Walter, E.B.; Berlanda-Scorza, F.; Stadlbauer, D.; Wilson, P.C.; Aydillo, T.; et al. A chimeric hemagglutinin-based universal influenza virus vaccine approach induces broad and long-lasting immunity in a randomized, placebo-controlled phase I trial. Nat. Med. 2021, 27, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Han, J.; Sun, W.; Puente-Massaguer, E.; Yu, W.; Palese, P.; Krammer, F.; Ward, A.B.; Wilson, I.A. Influenza chimeric hemagglutinin structures in complex with broadly protective antibodies to the stem and trimer interface. Proc. Natl. Acad. Sci. USA 2022, 119, e2200821119. [Google Scholar] [CrossRef] [PubMed]
- Puente-Massaguer, E.; Vasilev, K.; Beyer, A.; Loganathan, M.; Francis, B.; Scherm, M.J.; Arunkumar, G.A.; Gonzalez-Dominguez, I.; Zhu, X.; Wilson, I.A.; et al. Chimeric hemagglutinin split vaccines elicit broadly cross-reactive antibodies and protection against group 2 influenza viruses in mice. Sci. Adv. 2023, 9, eadi4753. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Chang, T.Z.; Wang, Y.; Li, S.; Wang, S.; Matsuyama, S.; Yu, G.; Compans, R.W.; Li, J.D.; Prausnitz, M.R.; et al. Heterosubtypic influenza protection elicited by double-layered polypeptide nanoparticles in mice. Proc. Natl. Acad. Sci. USA 2018, 115, E7758–E7767. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Wang, B.Z. A Perspective on Nanoparticle Universal Influenza Vaccines. ACS Infect. Dis. 2018, 4, 1656–1665. [Google Scholar] [CrossRef] [PubMed]
- Simeckova-Rosenberg, J.; Yun, Z.; Wyde, P.R.; Atassi, M.Z. Protection of mice against lethal viral infection by synthetic peptides corresponding to B- and T-cell recognition sites of influenza A hemagglutinin. Vaccine 1995, 13, 927–932. [Google Scholar] [CrossRef]
- Deliyannis, G.; Jackson, D.C.; Ede, N.J.; Zeng, W.; Hourdakis, I.; Sakabetis, E.; Brown, L.E. Induction of long-term memory CD8(+) T cells for recall of viral clearing responses against influenza virus. J. Virol. 2002, 76, 4212–4221. [Google Scholar] [CrossRef] [PubMed]
- Li, G.M.; Chiu, C.; Wrammert, J.; McCausland, M.; Andrews, S.F.; Zheng, N.Y.; Lee, J.H.; Huang, M.; Qu, X.; Edupuganti, S.; et al. Pandemic H1N1 influenza vaccine induces a recall response in humans that favors broadly cross-reactive memory B cells. Proc. Natl. Acad. Sci. USA 2012, 109, 9047–9052. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Wohlbold, T.J.; Zheng, N.Y.; Huang, M.; Huang, Y.; Neu, K.E.; Lee, J.; Wan, H.; Rojas, K.T.; Kirkpatrick, E.; et al. Influenza Infection in Humans Induces Broadly Cross-Reactive and Protective Neuraminidase-Reactive Antibodies. Cell 2018, 173, 417–429 e10. [Google Scholar] [CrossRef]
- Gilles, J.G.; Saint-Remy, J.M. Healthy subjects produce both anti-factor VIII and specific anti-idiotypic antibodies. J. Clin. Investig. 1994, 94, 1496–1505. [Google Scholar] [CrossRef]
- Al-Halifa, S.; Gauthier, L.; Arpin, D.; Bourgault, S.; Archambault, D. Nanoparticle-Based Vaccines Against Respiratory Viruses. Front. Immunol. 2019, 10, 22. [Google Scholar] [CrossRef]
- Pati, R.; Shevtsov, M.; Sonawane, A. Nanoparticle Vaccines Against Infectious Diseases. Front. Immunol. 2018, 9, 2224. [Google Scholar] [CrossRef]
- Pham, D.T.; Tiyaboonchai, W. Fibroin nanoparticles: A promising drug delivery system. Drug Deliv. 2020, 27, 431–448. [Google Scholar] [CrossRef] [PubMed]
- Madkhali, O.A. Drug Delivery of Gelatin Nanoparticles as a Biodegradable Polymer for the Treatment of Infectious Diseases: Perspectives and Challenges. Polymers 2023, 15, 4327. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.O.; Koutsakos, M.; van de Sandt, C.E.; Crawford, J.C.; Loh, L.; Sant, S.; Grzelak, L.; Allen, E.K.; Brahm, T.; Clemens, E.B.; et al. Immune cellular networks underlying recovery from influenza virus infection in acute hospitalized patients. Nat. Commun. 2021, 12, 2691. [Google Scholar] [CrossRef] [PubMed]
- Van de Ven, K.; de Heij, F.; van Dijken, H.; Ferreira, J.A.; de Jonge, J. Systemic and respiratory T-cells induced by seasonal H1N1 influenza protect against pandemic H2N2 in ferrets. Commun. Biol. 2020, 3, 564. [Google Scholar] [CrossRef] [PubMed]
- Eliasson, D.G.; Omokanye, A.; Schon, K.; Wenzel, U.A.; Bernasconi, V.; Bemark, M.; Kolpe, A.; El Bakkouri, K.; Ysenbaert, T.; Deng, L.; et al. M2e-tetramer-specific memory CD4 T cells are broadly protective against influenza infection. Mucosal Immunol. 2018, 11, 273–289. [Google Scholar] [CrossRef]
- Van Doorn, E.; Liu, H.; Ben-Yedidia, T.; Hassin, S.; Visontai, I.; Norley, S.; Frijlink, H.W.; Hak, E. Evaluating the immunogenicity and safety of a BiondVax-developed universal influenza vaccine (Multimeric-001) either as a standalone vaccine or as a primer to H5N1 influenza vaccine: Phase IIb study protocol. Medicine 2017, 96, e6339. [Google Scholar] [CrossRef] [PubMed]
- Atsmon, J.; Kate-Ilovitz, E.; Shaikevich, D.; Singer, Y.; Volokhov, I.; Haim, K.Y.; Ben-Yedidia, T. Safety and immunogenicity of multimeric-001--a novel universal influenza vaccine. J. Clin. Immunol. 2012, 32, 595–603. [Google Scholar] [CrossRef]
- Pleguezuelos, O.; James, E.; Fernandez, A.; Lopes, V.; Rosas, L.A.; Cervantes-Medina, A.; Cleath, J.; Edwards, K.; Neitzey, D.; Gu, W.; et al. Efficacy of FLU-v, a broad-spectrum influenza vaccine, in a randomized phase IIb human influenza challenge study. NPJ Vaccines 2020, 5, 22. [Google Scholar] [CrossRef]
- Fitzmaurice, C.J.; Brown, L.E.; McInerney, T.L.; Jackson, D.C. The assembly and immunological properties of non-linear synthetic immunogens containing T-cell and B-cell determinants. Vaccine 1996, 14, 553–560. [Google Scholar] [CrossRef]
- Zhao, R.Z.; Chen, X.; Ma, W.W.; Zhang, J.Y.; Guo, J.; Zhong, X.; Yao, J.C.; Sun, J.H.; Rubinfien, J.; Zhou, X.Y.; et al. A GPR174–CCL21 module imparts sexual dimorphism to humoral immunity. Nature 2020, 577, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, W.-W.; Wan, M.-Y.; She, J.-Y.; Zhao, S.-L.; Liu, D.-J.; Chang, H.-Y.; Deng, L. Sequential Immunizations with Influenza Neuraminidase Protein Followed by Peptide Nanoclusters Induce Heterologous Protection. Viruses 2024, 16, 77. https://doi.org/10.3390/v16010077
Song W-W, Wan M-Y, She J-Y, Zhao S-L, Liu D-J, Chang H-Y, Deng L. Sequential Immunizations with Influenza Neuraminidase Protein Followed by Peptide Nanoclusters Induce Heterologous Protection. Viruses. 2024; 16(1):77. https://doi.org/10.3390/v16010077
Chicago/Turabian StyleSong, Wen-Wen, Mu-Yang Wan, Jia-Yue She, Shi-Long Zhao, De-Jian Liu, Hai-Yan Chang, and Lei Deng. 2024. "Sequential Immunizations with Influenza Neuraminidase Protein Followed by Peptide Nanoclusters Induce Heterologous Protection" Viruses 16, no. 1: 77. https://doi.org/10.3390/v16010077