
CSF Extracellular Vesicle Aβ42 and Tau/Aβ42 Ratio Are Associated with Cognitive Impairment in Older People with HIV
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Assessment of Cognitive Function and HAND Diagnoses
2.3. Isolation and Characterization of Plasma and CSF Extracellular Vesicles
2.4. Meso Scale Discovery Assays
2.5. Proteinase K Treatment and Immunoblotting of CSF EVs
2.6. Data Analysis
3. Results
3.1. Study Participants
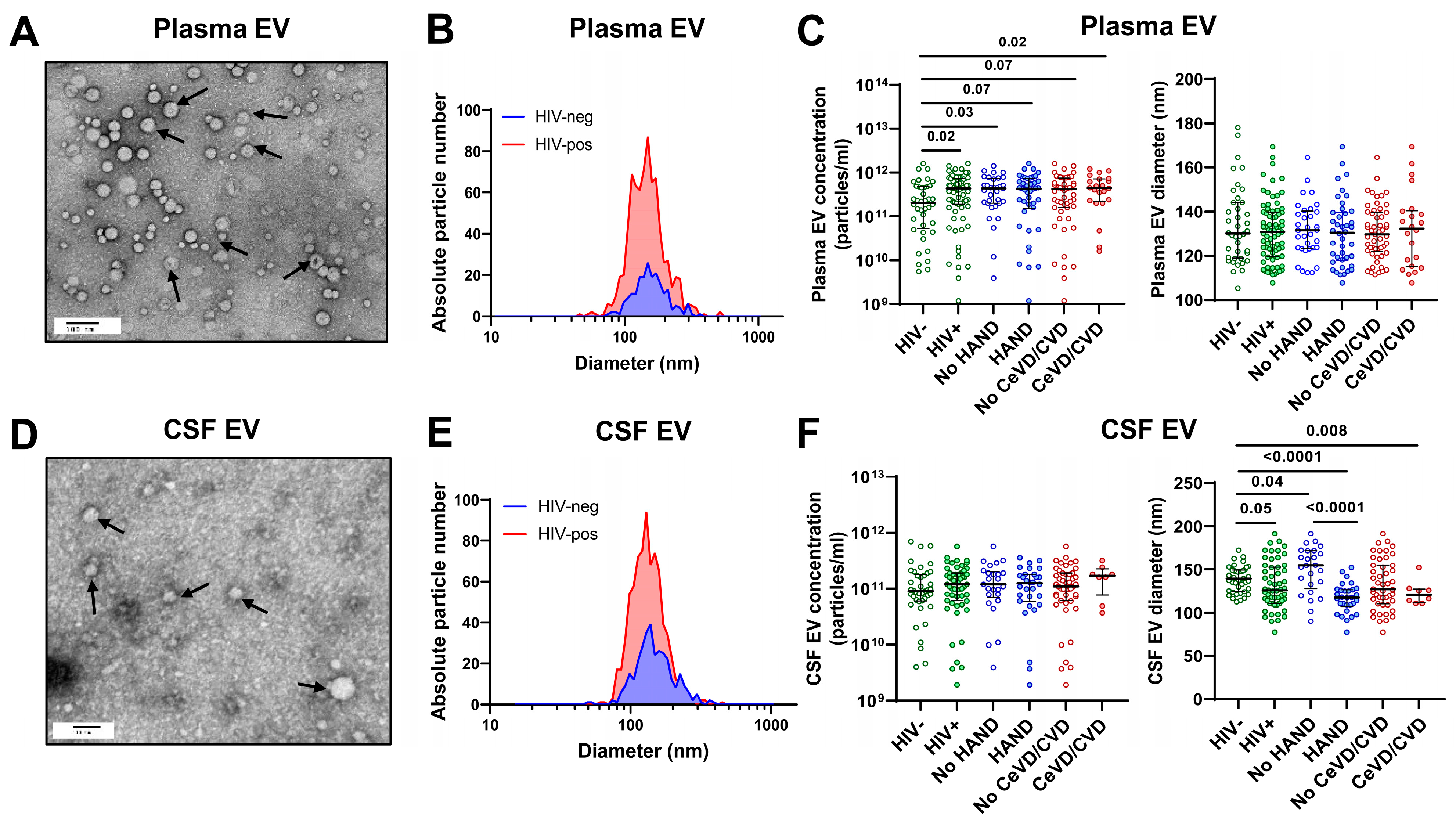
3.2. Characterization of Plasma and CSF Extracellular Vesicles
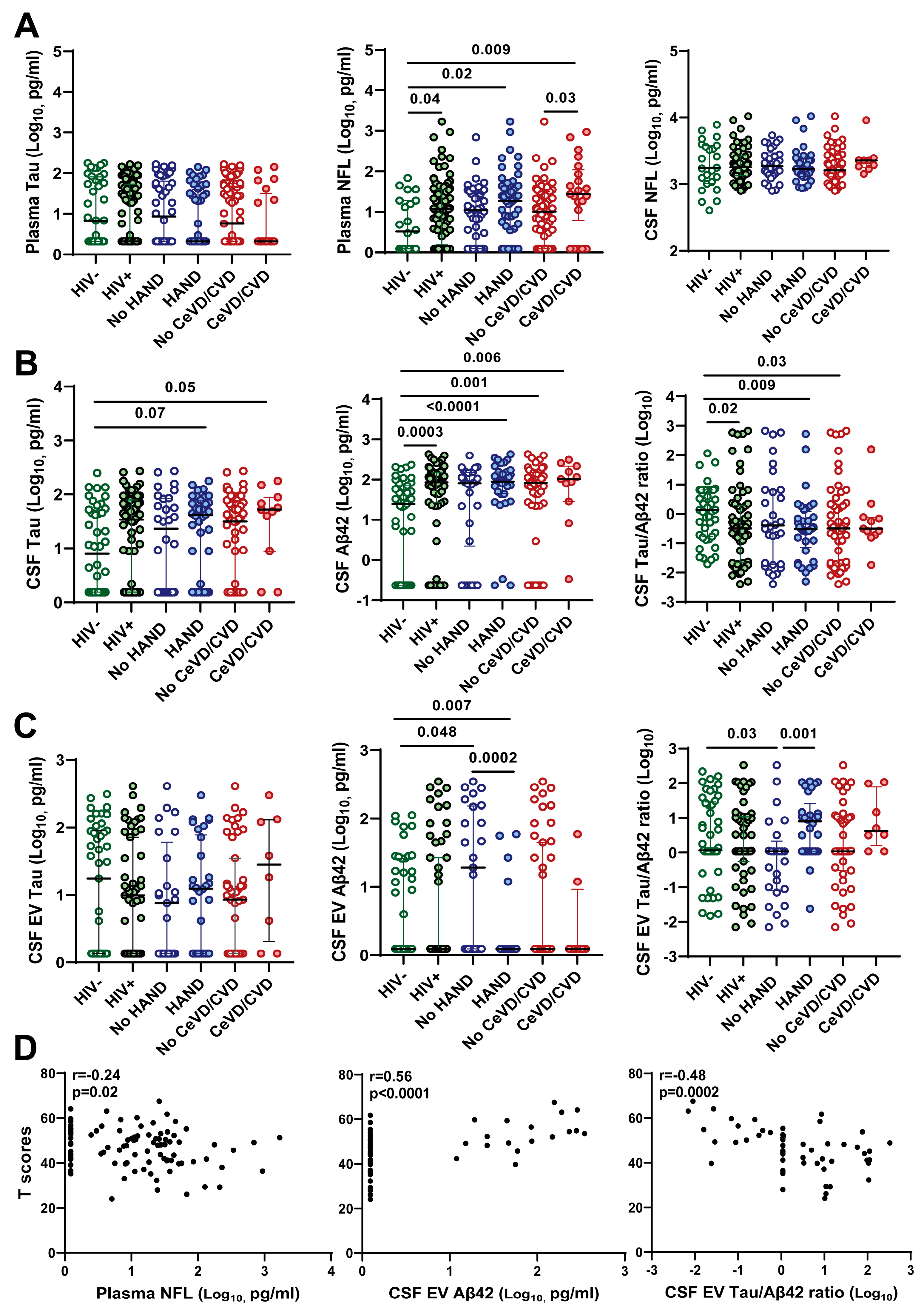
3.3. Association of CSF EV Aβ42 Levels and Tau/Aβ42 Ratio with HAND
3.4. Association of Decreased CSF EV Aβ42 and Increased CSF EV Tau/Aβ42 Ratio with Lower Neurocognitive Test Scores
3.5. Association of Soluble but Not EV-Associated Vascular Injury Markers with HAND
3.6. Comparative Abundance of Soluble vs. EV-Associated Biomarkers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aung, H.L.; Alagaratnam, J.; Chan, P.; Chow, F.C.; Joska, J.; Falutz, J.; Letendre, S.L.; Lin, W.; Munoz-Moreno, J.A.; Cinque, P.; et al. Cognitive Health in Persons With Human Immunodeficiency Virus: The Impact of Early Treatment, Comorbidities, and Aging. J. Infect. Dis. 2023, 227, S38–S47. [Google Scholar] [CrossRef] [PubMed]
- Gabuzda, D.; Jamieson, B.D.; Collman, R.G.; Lederman, M.M.; Burdo, T.H.; Deeks, S.G.; Dittmer, D.P.; Fox, H.S.; Funderburg, N.T.; Pahwa, S.G.; et al. Pathogenesis of Aging and Age-related Comorbidities in People with HIV: Highlights from the HIV ACTION Workshop. Pathog. Immun. 2020, 5, 143–174. [Google Scholar] [CrossRef] [PubMed]
- Winston, A.; Spudich, S. Cognitive disorders in people living with HIV. Lancet HIV 2020, 7, e504–e513. [Google Scholar] [CrossRef] [PubMed]
- Saylor, D.; Dickens, A.M.; Sacktor, N.; Haughey, N.; Slusher, B.; Pletnikov, M.; Mankowski, J.L.; Brown, A.; Volsky, D.J.; McArthur, J.C. HIV-associated neurocognitive disorder–pathogenesis and prospects for treatment. Nat. Rev. Neurol. 2016, 12, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.B.; Gianella, S.; Burdo, T.H.; Cinque, P.; Gisslen, M.; Letendre, S.; Nath, A.; Morgello, S.; Ndhlovu, L.C.; Spudich, S. Biotypes of Central Nervous System Complications in People With Human Immunodeficiency Virus: Virology, Immunology, and Neuropathology. J. Infect. Dis. 2023, 227, S3–S15. [Google Scholar] [CrossRef] [PubMed]
- Aung, H.L.; Aghvinian, M.; Gouse, H.; Robbins, R.N.; Brew, B.J.; Mao, L.; Cysique, L.A. Is There Any Evidence of Premature, Accentuated and Accelerated Aging Effects on Neurocognition in People Living with HIV? A Systematic Review. AIDS Behav. 2021, 25, 917–960. [Google Scholar] [CrossRef] [PubMed]
- Jakabek, D.; Rae, C.D.; Brew, B.J.; Cysique, L.A. Brain aging and cardiovascular factors in HIV: A longitudinal volume and shape MRI study. AIDS 2022, 36, 785–794. [Google Scholar] [CrossRef]
- Samboju, V.; Cobigo, Y.; Paul, R.; Naasan, G.; Hillis, M.; Tsuei, T.; Javandel, S.; Valcour, V.; Milanini, B. Cerebrovascular Disease Correlates With Longitudinal Brain Atrophy in Virally Suppressed Older People Living With HIV. J. Acquir. Immune Defic. Syndr. 2021, 87, 1079–1085. [Google Scholar] [CrossRef]
- Sanford, R.; Strain, J.; Dadar, M.; Maranzano, J.; Bonnet, A.; Mayo, N.E.; Scott, S.C.; Fellows, L.K.; Ances, B.M.; Collins, D.L. HIV infection and cerebral small vessel disease are independently associated with brain atrophy and cognitive impairment. AIDS 2019, 33, 1197–1205. [Google Scholar] [CrossRef]
- Gabuzda, D.; Yankner, B.A. Physiology: Inflammation links ageing to the brain. Nature 2013, 497, 197–198. [Google Scholar] [CrossRef]
- Rubin, L.H.; Sundermann, E.E.; Moore, D.J. The current understanding of overlap between characteristics of HIV-associated neurocognitive disorders and Alzheimer’s disease. J. Neurovirology 2019, 25, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Milanini, B.; Valcour, V. Differentiating HIV-Associated Neurocognitive Disorders From Alzheimer’s Disease: An Emerging Issue in Geriatric NeuroHIV. Curr. HIV/AIDS Rep. 2017, 14, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Gisslen, M.; Price, R.W.; Andreasson, U.; Norgren, N.; Nilsson, S.; Hagberg, L.; Fuchs, D.; Spudich, S.; Blennow, K.; Zetterberg, H. Plasma Concentration of the Neurofilament Light Protein (NFL) is a Biomarker of CNS Injury in HIV Infection: A Cross-Sectional Study. EBioMedicine 2016, 3, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, A.; Blennow, K.; Hagberg, L.; Nilsson, S.; Price, R.W.; Schouten, J.; Spudich, S.; Underwood, J.; Zetterberg, H.; Gisslen, M. Neurofilament light chain protein as a marker of neuronal injury: Review of its use in HIV-1 infection and reference values for HIV-negative controls. Expert. Rev. Mol. Diagn. 2017, 17, 761–770. [Google Scholar] [CrossRef]
- McGuire, J.L.; Gill, A.J.; Douglas, S.D.; Kolson, D.L.; CNS HIV Anti-Retroviral Therapy Effects Research (CHARTER) group. Central and peripheral markers of neurodegeneration and monocyte activation in HIV-associated neurocognitive disorders. J. Neurovirology 2015, 21, 439–448. [Google Scholar] [CrossRef]
- Guha, D.; Mukerji, S.S.; Chettimada, S.; Misra, V.; Lorenz, D.R.; Morgello, S.; Gabuzda, D. Cerebrospinal fluid extracellular vesicles and neurofilament light protein as biomarkers of central nervous system injury in HIV-infected patients on antiretroviral therapy. AIDS 2019, 33, 615–625. [Google Scholar] [CrossRef]
- De Almeida, S.M.; Ribeiro, C.E.; Rotta, I.; Piovesan, M.; Tang, B.; Vaida, F.; Raboni, S.M.; Letendre, S.; Potter, M.; Batistela Fernandes, M.S.; et al. Biomarkers of neuronal injury and amyloid metabolism in the cerebrospinal fluid of patients infected with HIV-1 subtypes B and C. J. NeuroVirology 2018, 24, 28–40. [Google Scholar] [CrossRef]
- Mackiewicz, M.M.; Overk, C.; Achim, C.L.; Masliah, E. Pathogenesis of age-related HIV neurodegeneration. J. NeuroVirology 2019, 25, 622–633. [Google Scholar] [CrossRef]
- Gonzalez, J.; Wilson, A.; Byrd, D.; Cortes, E.P.; Crary, J.F.; Morgello, S. Neuronal accumulation of hyperphosphorylated tau protein predicts stable memory impairment in people living with HIV. AIDS 2023, 37, 1247–1256. [Google Scholar] [CrossRef]
- Solomon, I.H.; De Girolami, U.; Chettimada, S.; Misra, V.; Singer, E.J.; Gabuzda, D. Brain and liver pathology, amyloid deposition, and interferon responses among older HIV-positive patients in the late HAART era. BMC Infect. Dis. 2017, 17, 151. [Google Scholar] [CrossRef]
- Hansson, O.; Seibyl, J.; Stomrud, E.; Zetterberg, H.; Trojanowski, J.Q.; Bittner, T.; Lifke, V.; Corradini, V.; Eichenlaub, U.; Batrla, R.; et al. CSF biomarkers of Alzheimer’s disease concord with amyloid-β PET and predict clinical progression: A study of fully automated immunoassays in BioFINDER and ADNI cohorts. Alzheimers Dement. 2018, 14, 1470–1481. [Google Scholar] [CrossRef]
- Li, G.; Sokal, I.; Quinn, J.F.; Leverenz, J.B.; Brodey, M.; Schellenberg, G.D.; Kaye, J.A.; Raskind, M.A.; Zhang, J.; Peskind, E.R.; et al. CSF tau/Abeta42 ratio for increased risk of mild cognitive impairment: A follow-up study. Neurology 2007, 69, 631–639. [Google Scholar] [CrossRef]
- Park, J.C.; Han, S.H.; Yi, D.; Byun, M.S.; Lee, J.H.; Jang, S.; Ko, K.; Jeon, S.Y.; Lee, Y.S.; Kim, Y.K.; et al. Plasma tau/amyloid-beta1-42 ratio predicts brain tau deposition and neurodegeneration in Alzheimer’s disease. Brain 2019, 142, 771–786. [Google Scholar] [CrossRef]
- Pereira, J.B.; Janelidze, S.; Stomrud, E.; Palmqvist, S.; van Westen, D.; Dage, J.L.; Mattsson-Carlgren, N.; Hansson, O. Plasma markers predict changes in amyloid, tau, atrophy and cognition in non-demented subjects. Brain 2021, 144, 2826–2836. [Google Scholar] [CrossRef]
- Sundermann, E.E.; Campbell, L.M.; Villers, O.; Bondi, M.W.; Gouaux, B.; Salmon, D.P.; Galasko, D.; Soontornniyomkij, V.; Ellis, R.J.; Moore, D.J. Alzheimer’s Disease Pathology in Middle Aged and Older People with HIV: Comparisons with Non-HIV Controls on a Healthy Aging and Alzheimer’s Disease Trajectory and Relationships with Cognitive Function. Viruses 2023, 15, 1319. [Google Scholar] [CrossRef]
- Cooley, S.A.; Nelson, B.; Boerwinkle, A.; Yarasheski, K.E.; Kirmess, K.M.; Meyer, M.R.; Schindler, S.E.; Morris, J.C.; Fagan, A.; Ances, B.M.; et al. Plasma Abeta42/Abeta40 Ratios in Older People With Human Immunodeficiency Virus. Clin. Infect. Dis. 2023, 76, 1776–1783. [Google Scholar] [CrossRef]
- Cooley, S.A.; Strain, J.F.; Beaumont, H.; Boerwinkle, A.H.; Doyle, J.; Morris, J.C.; Benzinger, T.L.; Ances, B.M. Tau Positron Emission Tomography Binding Is Not Elevated in HIV-Infected Individuals. J. Infect. Dis. 2019, 220, 68–72. [Google Scholar] [CrossRef]
- Albrecht, D.S.; Sagare, A.; Pachicano, M.; Sweeney, M.D.; Toga, A.; Zlokovic, B.; Chui, H.; Joe, E.; Schneider, L.; Morris, J.C.; et al. Early neuroinflammation is associated with lower amyloid and tau levels in cognitively normal older adults. Brain Behav. Immun. 2021, 94, 299–307. [Google Scholar] [CrossRef]
- Fields, J.A.; Swinton, M.K.; Soontornniyomkij, B.; Carson, A.; Achim, C.L. Beta amyloid levels in cerebrospinal fluid of HIV-infected people vary by exposure to antiretroviral therapy. AIDS 2020, 34, 1001–1007. [Google Scholar] [CrossRef]
- Lobo, J.D.; Moore, D.J.; Bondi, M.W.; Soontornniyomkij, V.; Soontornniyomkij, B.; Gouaux, B.; Achim, C.L.; Ellis, R.J.; Sundermann, E.E. CSF markers of AD-related pathology relate specifically to memory impairment in older people with HIV: A pilot study. J. NeuroVirology 2022, 28, 162–167. [Google Scholar] [CrossRef]
- Morgello, S.; Cortes, E.P.; Gensler, G.; Meloni, G.; Jacobs, M.M.; Murray, J.; Borukov, V.; Crary, J.F. HIV disease duration, but not active brain infection, predicts cortical amyloid beta deposition. AIDS 2021, 35, 1403–1412. [Google Scholar] [CrossRef]
- Mukerji, S.S.; Petersen, K.J.; Pohl, K.M.; Dastgheyb, R.M.; Fox, H.S.; Bilder, R.M.; Brouillette, M.J.; Gross, A.L.; Scott-Sheldon, L.A.J.; Paul, R.H.; et al. Machine Learning Approaches to Understand Cognitive Phenotypes in People With HIV. J. Infect. Dis. 2023, 227, S48–S57. [Google Scholar] [CrossRef]
- Thompson, A.G.; Gray, E.; Heman-Ackah, S.M.; Mager, I.; Talbot, K.; Andaloussi, S.E.; Wood, M.J.; Turner, M.R. Extracellular vesicles in neurodegenerative disease–pathogenesis to biomarkers. Nat. Rev. Neurol. 2016, 12, 346–357. [Google Scholar] [CrossRef]
- Guha, D.; Lorenz, D.R.; Misra, V.; Chettimada, S.; Morgello, S.; Gabuzda, D. Proteomic analysis of cerebrospinal fluid extracellular vesicles reveals synaptic injury, inflammation, and stress response markers in HIV patients with cognitive impairment. J. Neuroinflammation 2019, 16, 254. [Google Scholar] [CrossRef]
- Fiandaca, M.S.; Kapogiannis, D.; Mapstone, M.; Boxer, A.; Eitan, E.; Schwartz, J.B.; Abner, E.L.; Petersen, R.C.; Federoff, H.J.; Miller, B.L.; et al. Identification of preclinical Alzheimer’s disease by a profile of pathogenic proteins in neurally derived blood exosomes: A case-control study. Alzheimers Dement. 2015, 11, 600–607.e1. [Google Scholar] [CrossRef]
- Lim, C.Z.J.; Zhang, Y.; Chen, Y.; Zhao, H.; Stephenson, M.C.; Ho, N.R.Y.; Chen, Y.; Chung, J.; Reilhac, A.; Loh, T.P.; et al. Subtyping of circulating exosome-bound amyloid beta reflects brain plaque deposition. Nat. Commun. 2019, 10, 1144. [Google Scholar] [CrossRef]
- Pulliam, L.; Sun, B.; Mustapic, M.; Chawla, S.; Kapogiannis, D. Plasma neuronal exosomes serve as biomarkers of cognitive impairment in HIV infection and Alzheimer’s disease. J. NeuroVirology 2019, 25, 702–709. [Google Scholar] [CrossRef]
- Coleman, B.M.; Hill, A.F. Extracellular vesicles--Their role in the packaging and spread of misfolded proteins associated with neurodegenerative diseases. Semin. Cell Dev. Biol. 2015, 40, 89–96. [Google Scholar] [CrossRef]
- Wang, Y.; Balaji, V.; Kaniyappan, S.; Kruger, L.; Irsen, S.; Tepper, K.; Chandupatla, R.; Maetzler, W.; Schneider, A.; Mandelkow, E.; et al. The release and trans-synaptic transmission of Tau via exosomes. Mol. Neurodegener. 2017, 12, 5. [Google Scholar] [CrossRef]
- Ruan, Z.; Pathak, D.; Venkatesan Kalavai, S.; Yoshii-Kitahara, A.; Muraoka, S.; Bhatt, N.; Takamatsu-Yukawa, K.; Hu, J.; Wang, Y.; Hersh, S.; et al. Alzheimer’s disease brain-derived extracellular vesicles spread tau pathology in interneurons. Brain 2021, 144, 288–309. [Google Scholar] [CrossRef]
- Winston, C.N.; Goetzl, E.J.; Akers, J.C.; Carter, B.S.; Rockenstein, E.M.; Galasko, D.; Masliah, E.; Rissman, R.A. Prediction of conversion from mild cognitive impairment to dementia with neuronally derived blood exosome protein profile. Alzheimers Dement. 2016, 3, 63–72. [Google Scholar] [CrossRef]
- Jia, L.; Qiu, Q.; Zhang, H.; Chu, L.; Du, Y.; Zhang, J.; Zhou, C.; Liang, F.; Shi, S.; Wang, S.; et al. Concordance between the assessment of Abeta42, T-tau, and P-T181-tau in peripheral blood neuronal-derived exosomes and cerebrospinal fluid. Alzheimers Dement. 2019, 15, 1071–1080. [Google Scholar] [CrossRef]
- Chettimada, S.; Lorenz, D.R.; Misra, V.; Dillon, S.T.; Reeves, R.K.; Manickam, C.; Morgello, S.; Kirk, G.D.; Mehta, S.H.; Gabuzda, D. Exosome markers associated with immune activation and oxidative stress in HIV patients on antiretroviral therapy. Sci. Rep. 2018, 8, 7227. [Google Scholar] [CrossRef]
- Kodidela, S.; Gerth, K.; Haque, S.; Gong, Y.; Ismael, S.; Singh, A.; Tauheed, I.; Kumar, S. Extracellular Vesicles: A Possible Link between HIV and Alzheimer’s Disease-Like Pathology in HIV Subjects? Cells 2019, 8, 968. [Google Scholar] [CrossRef]
- Kodidela, S.; Gerth, K.; Sinha, N.; Kumar, A.; Kumar, P.; Kumar, S. Circulatory Astrocyte and Neuronal EVs as Potential Biomarkers of Neurological Dysfunction in HIV-Infected Subjects and Alcohol/Tobacco Users. Diagnostics 2020, 10, 349. [Google Scholar] [CrossRef]
- Hu, G.; Yang, L.; Cai, Y.; Niu, F.; Mezzacappa, F.; Callen, S.; Fox, H.S.; Buch, S. Emerging roles of extracellular vesicles in neurodegenerative disorders: Focus on HIV-associated neurological complications. Cell Death Dis. 2016, 7, e2481. [Google Scholar] [CrossRef]
- Heaton, R.K.; Clifford, D.B.; Franklin, D.R., Jr.; Woods, S.P.; Ake, C.; Vaida, F.; Ellis, R.J.; Letendre, S.L.; Marcotte, T.D.; Atkinson, J.H.; et al. HIV-associated neurocognitive disorders persist in the era of potent antiretroviral therapy: CHARTER Study. Neurology 2010, 75, 2087–2096. [Google Scholar] [CrossRef]
- Morgello, S.; Gelman, B.B.; Kozlowski, P.B.; Vinters, H.V.; Masliah, E.; Cornford, M.; Cavert, W.; Marra, C.; Grant, I.; Singer, E.J. The National NeuroAIDS Tissue Consortium: A new paradigm in brain banking with an emphasis on infectious disease. Neuropathol. Appl. Neurobiol. 2001, 27, 326–335. [Google Scholar] [CrossRef]
- Woods, S.P.; Rippeth, J.D.; Frol, A.B.; Levy, J.K.; Ryan, E.; Soukup, V.M.; Hinkin, C.H.; Lazzaretto, D.; Cherner, M.; Marcotte, T.D.; et al. Interrater reliability of clinical ratings and neurocognitive diagnoses in HIV. J. Clin. Exp. Neuropsychol. 2004, 26, 759–778. [Google Scholar] [CrossRef]
- Sundermann, E.E.; Bondi, M.W.; Campbell, L.M.; Gouaux, B.; Moore, R.C.; Soontornniyomkij, V.; Moore, D.J. Distinguishing Amnestic Mild Cognitive Impairment From HIV-Associated Neurocognitive Disorders. J. Infect. Dis. 2021, 224, 435–442. [Google Scholar] [CrossRef]
- Antinori, A.; Arendt, G.; Becker, J.T.; Brew, B.J.; Byrd, D.A.; Cherner, M.; Clifford, D.B.; Cinque, P.; Epstein, L.G.; Goodkin, K.; et al. Updated research nosology for HIV-associated neurocognitive disorders. Neurology 2007, 69, 1789–1799. [Google Scholar] [CrossRef]
- Lorenz, D.R.; Mukerji, S.S.; Misra, V.; Uno, H.; Gelman, B.B.; Moore, D.J.; Singer, E.J.; Morgello, S.; Gabuzda, D. Multimorbidity networks associated with frailty among middle-aged and older people with HIV. AIDS 2021, 35, 2451–2461. [Google Scholar] [CrossRef]
- Chatterjee, P.; Pedrini, S.; Stoops, E.; Goozee, K.; Villemagne, V.L.; Asih, P.R.; Verberk, I.M.W.; Dave, P.; Taddei, K.; Sohrabi, H.R.; et al. Plasma glial fibrillary acidic protein is elevated in cognitively normal older adults at risk of Alzheimer’s disease. Transl. Psychiatry 2021, 11, 27. [Google Scholar] [CrossRef]
- Pereira, J.B.; Janelidze, S.; Smith, R.; Mattsson-Carlgren, N.; Palmqvist, S.; Teunissen, C.E.; Zetterberg, H.; Stomrud, E.; Ashton, N.J.; Blennow, K.; et al. Plasma GFAP is an early marker of amyloid-beta but not tau pathology in Alzheimer’s disease. Brain 2021, 144, 3505–3516. [Google Scholar] [CrossRef]
- Guha, D.; Misra, V.; Yin, J.; Horiguchi, M.; Uno, H.; Gabuzda, D. Vascular injury markers associated with cognitive impairment in people with HIV on suppressive antiretroviral therapy. AIDS 2023, 37, 2137–2147. [Google Scholar] [CrossRef]
- Busche, M.A.; Hyman, B.T. Synergy between amyloid-beta and tau in Alzheimer’s disease. Nat. Neurosci. 2020, 23, 1183–1193. [Google Scholar] [CrossRef]
- Pereira, J.B.; Janelidze, S.; Ossenkoppele, R.; Kvartsberg, H.; Brinkmalm, A.; Mattsson-Carlgren, N.; Stomrud, E.; Smith, R.; Zetterberg, H.; Blennow, K.; et al. Untangling the association of amyloid-beta and tau with synaptic and axonal loss in Alzheimer’s disease. Brain 2021, 144, 310–324. [Google Scholar] [CrossRef]
- Mohamed, M.; Skolasky, R.L.; Zhou, Y.; Ye, W.; Brasic, J.R.; Brown, A.; Pardo, C.A.; Barker, P.B.; Wong, D.F.; Sacktor, N. Beta-amyloid (Abeta) uptake by PET imaging in older HIV+ and HIV- individuals. J. Neurovirology 2020, 26, 382–390. [Google Scholar] [CrossRef]
- Howdle, G.C.; Quide, Y.; Kassem, M.S.; Johnson, K.; Rae, C.D.; Brew, B.J.; Cysique, L.A. Brain amyloid in virally suppressed HIV-associated neurocognitive disorder. Neurol. Neuroimmunol. Neuroinflammation 2020, 7, e739. [Google Scholar] [CrossRef]
- Peterson, J.; Gisslen, M.; Zetterberg, H.; Fuchs, D.; Shacklett, B.L.; Hagberg, L.; Yiannoutsos, C.T.; Spudich, S.S.; Price, R.W. Cerebrospinal fluid (CSF) neuronal biomarkers across the spectrum of HIV infection: Hierarchy of injury and detection. PLoS ONE 2014, 9, e116081. [Google Scholar] [CrossRef]
- Anthony, I.C.; Ramage, S.N.; Carnie, F.W.; Simmonds, P.; Bell, J.E. Accelerated Tau deposition in the brains of individuals infected with human immunodeficiency virus-1 before and after the advent of highly active anti-retroviral therapy. Acta Neuropathol. 2006, 111, 529–538. [Google Scholar] [CrossRef]
- Brew, B.J.; Pemberton, L.; Blennow, K.; Wallin, A.; Hagberg, L. CSF amyloid beta42 and tau levels correlate with AIDS dementia complex. Neurology 2005, 65, 1490–1492. [Google Scholar] [CrossRef]
- Gisslen, M.; Krut, J.; Andreasson, U.; Blennow, K.; Cinque, P.; Brew, B.J.; Spudich, S.; Hagberg, L.; Rosengren, L.; Price, R.W.; et al. Amyloid and tau cerebrospinal fluid biomarkers in HIV infection. BMC Neurol. 2009, 9, 63. [Google Scholar] [CrossRef]
- Ellis, R.J.; Chenna, A.; Petropoulos, C.J.; Lie, Y.; Curanovic, D.; Crescini, M.; Winslow, J.; Sundermann, E.; Tang, B.; Letendre, S.L. Higher cerebrospinal fluid biomarkers of neuronal injury in HIV-associated neurocognitive impairment. J. Neurovirology 2022, 28, 438–445. [Google Scholar] [CrossRef]
- Clifford, D.B.; Fagan, A.M.; Holtzman, D.M.; Morris, J.C.; Teshome, M.; Shah, A.R.; Kauwe, J.S. CSF biomarkers of Alzheimer disease in HIV-associated neurologic disease. Neurology 2009, 73, 1982–1987. [Google Scholar] [CrossRef]
- Gisslen, M.; Keating, S.M.; Spudich, S.; Arechiga, V.; Stephenson, S.; Zetterberg, H.; Di Germanio, C.; Blennow, K.; Fuchs, D.; Hagberg, L.; et al. Compartmentalization of cerebrospinal fluid inflammation across the spectrum of untreated HIV-1 infection, central nervous system injury and viral suppression. PLoS ONE 2021, 16, e0250987. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| HIV− (n = 86) | HIV+ (n = 98) | No HAND (n = 47) | HAND * (n = 51) | p-Value | |
|---|---|---|---|---|---|
| Age (years) | 54 (46–61) | 53 (47–59) | 52 (47–57) | 53 (49–61) | 0.55 |
| Male gender (n, %) | 60 (70) | 82 (84) | 47 (100) | 40 (78) | 0.0006 |
| Race (n, %) # | 0.68 | ||||
| Black | 18 (28) | 27 (28) | 15 (32) | 12 (24) | |
| White | 46 (71) | 68 (69) | 31 (66) | 37 (72) | |
| Other | 1 (1) | 3 (3) | 1 (2) | 2 (4) | |
| Current smoking (n, %) | 27 (66) | 48 (49) | 19 (40) | 29 (56.9) | 0.11 |
| Current alcohol use (n, %) | 45 (46) | 23 (49) | 22 (43) | 0.69 | |
| Diabetes (n, %) | 16 (16) | 6 (13) | 10 (20) | 0.42 | |
| Hypertension (n, %) | 50 (51.0) | 22 (47) | 28 (55) | 0.54 | |
| Hyperlipidemia (n, %) | 28 (29) | 10 (21) | 18 (35) | 0.18 | |
| Cerebrovascular disease/CVD (n, %) | 23 (24) | 5 (11) | 18 (35) | 0.005 | |
| Global neurocognitive T score | 48 (41–53) | 53 (50–57) | 41 (37–44) | <0.0001 | |
| Global clinical rating | 4 (3–5) | 3 (2–4) | 5 (5–6) | <0.0001 | |
| CSF protein | 37 (31–47) | 42 (33–52) | 35 (27–44) | 0.04 | |
| CSF WBC (cells/µL) | 1 (1–3) | 1 (0–3) | 1 (1–3) | 0.77 | |
| Duration of HIV infection (years) | 15 (11–21) | 15 (11–20) | 15 (11–22) | 0.89 | |
| Plasma viral load (<200 copies/mL) | 94 (96) | 45 (96) | 49 (96) | 1.00 | |
| CSF viral load (<50 copies/mL) Ϯ | 87 (96) | 43 (98) | 44 (94) | 0.62 | |
| CD4 count (cells/µL) | 540 (372–738) | 589 (382–845) | 537 (343–679) | 0.41 | |
| Nadir CD4 count (cells/µL) | 84 (14–223) | 88 (12–262) | 75 (16–183) | 0.62 | |
| ART use (n, %) | 98 (100) | 47 (100) | 51 (100) | 1.00 | |
| Protease inhibitors | 59 (60) | 29 (62) | 30 (59) | ||
| Integrase inhibitors | 19 (19) | 9 (19) | 10 (20) | ||
| Duration of ART (years) | 9 (5–14) | 9 (5–14) | 10 (5–14) | 0.84 |
| Variable | β Coefficient | SE | 95% CI | p-Value |
|---|---|---|---|---|
| Single-marker models | ||||
| CSF EV Aβ42 | 1.851 | 0.3873 | 1.074 to 2.628 | <0.0001 |
| CSF EV Tau/Aβ42 ratio | −1.218 | 0.3116 | −1.843 to −0.593 | 0.0003 |
| Plasma NFL | −0.6464 | 0.3862 | −1.413 to 0.1206 | 0.098 |
| Plasma GFAP | −0.9767 | 0.6751 | −2.318 to 0.3641 | 0.15 |
| Optimal model | ||||
| CSF EV Aβ42 | 1.824 | 0.372 | 1.077–2.571 | <0.0001 |
| Plasma NFL | −1.276 | 0.548 | −2.377 to −0.175 | 0.024 |
| CSF EV Ab42 Top/Middle Tertile (n = 18) | CSF EV Ab42 Bottom Tertile (n = 37) | p-Value | CSF EV Tau/Ab42 Ratio Bottom/Middle Tertile (n = 36) | CSF EV Tau/Ab42 Ratio Top Tertile (n = 19) | p-Value | |
|---|---|---|---|---|---|---|
| Age (years) | 53 [46, 57] | 50 [47, 54] | 0.83 | 52 [47, 57] | 49 [44, 53] | 0.43 |
| Female gender (n, %) | 1 (6) | 8 (22) | 0.26 | 3 (8) | 6 (32) | 0.067 |
| Race (n, %) | 0.68 | |||||
| Black | 8 (44) | 9 (24) | 0.37 | 11 (31) | 6 (32) | 0.55 |
| White | 10 (56) | 27 (73) | 25 (69) | 12 (63) | ||
| Other | 0 (0) | 1 (3) | 0 (0) | 1 (6) | ||
| Duration of HIV infection (years) | 14 [9, 19] | 14 [8, 22] | 0.50 | 15 [9, 22] | 14 [8, 17] | 0.67 |
| Nadir CD4 count (cells/µL) | 181 [61, 256] | 101 [21, 200] | 0.17 | 146 [42, 223] | 101 [39, 191] | 0.68 |
| CD4 count (cells/µL) | 602 [463, 884] | 537 [414, 706] | 0.43 | 637 [448, 860] | 506 [418, 603] | 0.19 |
| APOE4 carrier (n, %) | 6 (35) | 7 (28) | 0.87 | 11 (39) | 2 (14) | 0.19 |
| HAND diagnosis (n, %) | 4 (22) | 26 (70) | 0.0021 | 15 (42) | 15 (79) | 0.019 |
| HAND category (n, %) | ||||||
| ANI | 3 (17) | 18 (49) | 0.0035 | 11 (31) | 10 (53) | 0.028 |
| MND | 1 (6) | 8 (22) | 4 (11) | 5 (26) | ||
| Progression to HAD within 2.5 years (n, %) | 0 (0) | 6 (16) | 0.20 | 2 (6) | 4 (21) | 0.21 |
| Global T score | 53 [49, 59] | 44 [38, 50] | <0.001 | 50 [42, 55] | 42 [35, 48] | 0.010 |
| Domain T score | ||||||
| Abstraction/Executive function | 55 [50, 61] | 46 [40, 57] | 0.0090 | 51 [46, 59] | 45 [40, 54] | 0.062 |
| Speed of information processing | 58 [51, 63] | 45 [36, 50] | <0.001 | 51 [45, 60] | 43 [31, 49] | 0.017 |
| Attention/Working memory | 56 [52, 59] | 44 [37, 50] | <0.001 | 52 [43, 57] | 42 [36, 50] | 0.010 |
| Learning | 53 [41, 58] | 40 [34, 48] | 0.0021 | 47 [37, 55] | 40 [33, 45] | 0.061 |
| Memory | 54 [45, 58] | 42 [33, 48] | 0.0032 | 48 [36, 55] | 38 [32, 50] | 0.056 |
| Verbal fluency | 54 [50, 60] | 46 [41, 53] | 0.021 | 52 [45, 59] | 43 [39, 50] | 0.053 |
| Motor | 50 [41, 57] | 38 [31, 49] | 0.010 | 48 [36, 55] | 38 [26, 48] | 0.025 |
| Global T score < 40 (n, %) | 1 (6) | 14 (38) | 0.012 | 8 (22) | 7 (37) | 0.401 |
| Domain T score < 40 (n, %) | ||||||
| Abstraction/Executive function | 1 (6) | 10 (27) | 0.080 | 6 (17) | 5 (26) | 0.62 |
| Speed of information processing | 1 (6) | 13 (35) | 0.022 | 6 (17) | 8 (42) | 0.083 |
| Attention/Working memory | 0 (0) | 13 (35) | 0.0049 | 5 (14) | 8 (42) | 0.045 |
| Learning | 5 (28) | 19 (51) | 0.15 | 14 (39) | 10 (53) | 0.49 |
| Memory | 3 (17) | 18 (49) | 0.037 | 11 (31) | 10 (53) | 0.19 |
| Verbal fluency | 3 (17) | 9 (24) | 0.73 | 6 (17) | 6 (32) | 0.35 |
| Motor | 4 (22) | 20 (54) | 0.042 | 12 (33) | 12 (63) | 0.067 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guha, D.; Misra, V.; Chettimada, S.; Yin, J.; Gabuzda, D. CSF Extracellular Vesicle Aβ42 and Tau/Aβ42 Ratio Are Associated with Cognitive Impairment in Older People with HIV. Viruses 2024, 16, 72. https://doi.org/10.3390/v16010072
Guha D, Misra V, Chettimada S, Yin J, Gabuzda D. CSF Extracellular Vesicle Aβ42 and Tau/Aβ42 Ratio Are Associated with Cognitive Impairment in Older People with HIV. Viruses. 2024; 16(1):72. https://doi.org/10.3390/v16010072
Chicago/Turabian StyleGuha, Debjani, Vikas Misra, Sukrutha Chettimada, Jun Yin, and Dana Gabuzda. 2024. "CSF Extracellular Vesicle Aβ42 and Tau/Aβ42 Ratio Are Associated with Cognitive Impairment in Older People with HIV" Viruses 16, no. 1: 72. https://doi.org/10.3390/v16010072