
AAV Vectors Pseudotyped with Capsids from Porcine and Bovine Species Mediate In Vitro and In Vivo Gene Delivery

Abstract
:1. Introduction
2. Materials and Methods
3. Results
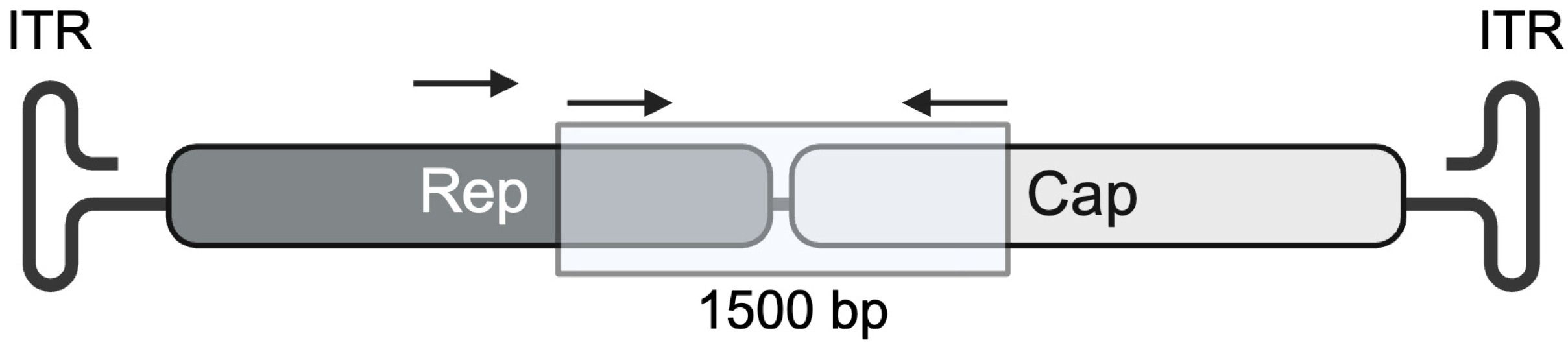
3.1. Analysis of Porcine AAV Screening Amplicon
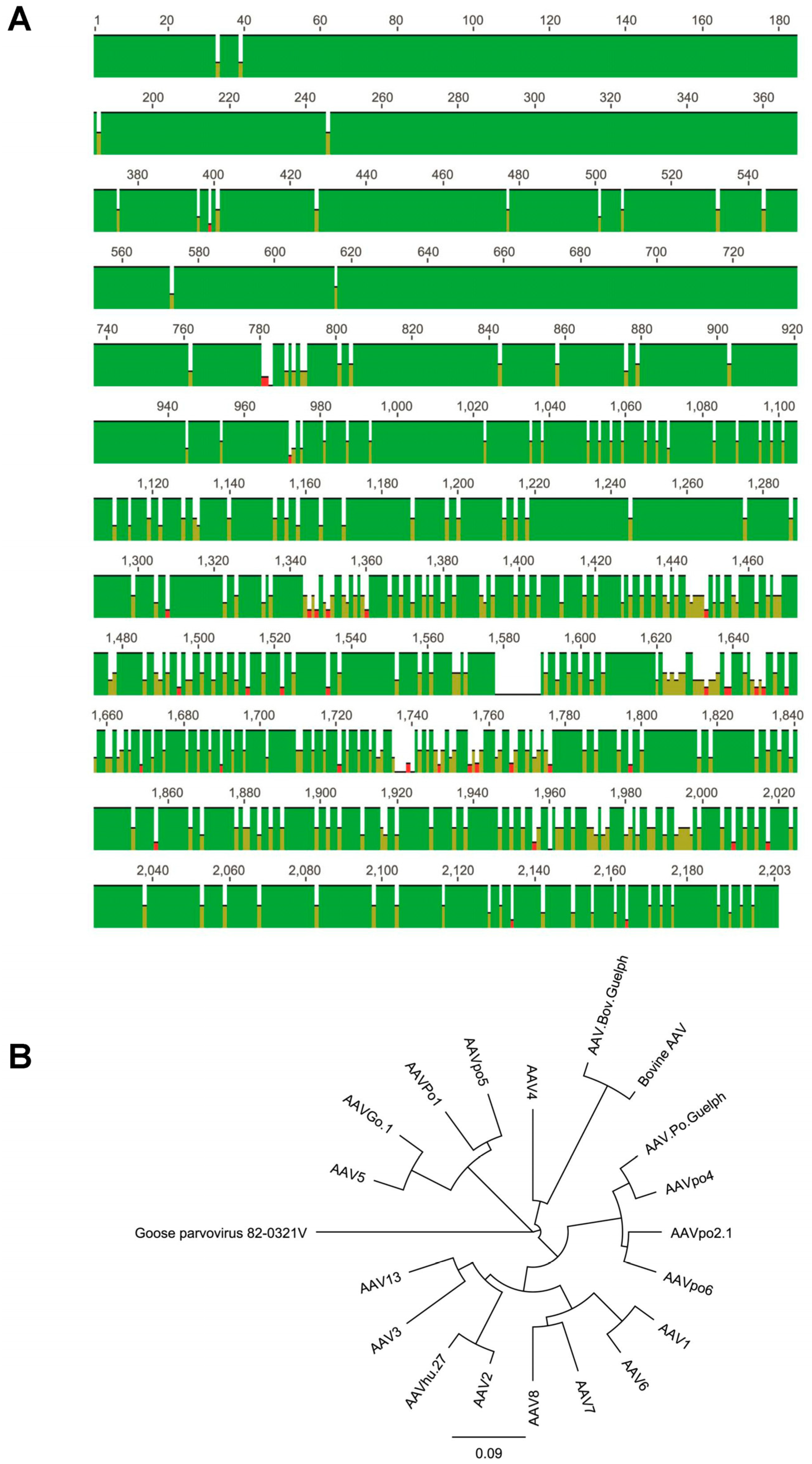
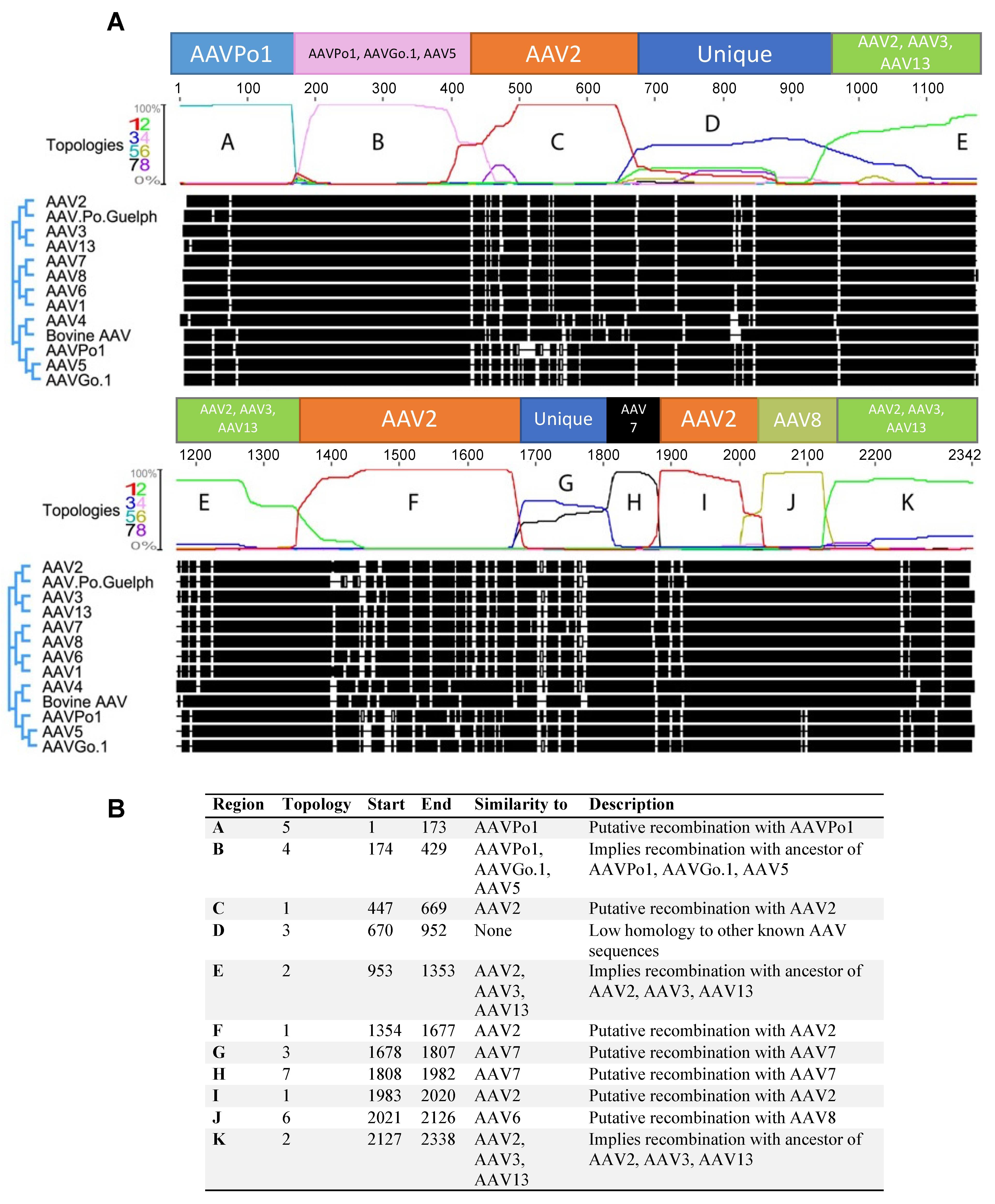
3.2. Numerous Recombination Events Have Occurred between Human AAVs and Animal-Derived AAVs to Form AAV.Po.Guelph
3.3. AAV.Bov.Guelph Capsid Protein Is Highly Similar to the Bovine AAV Capsid
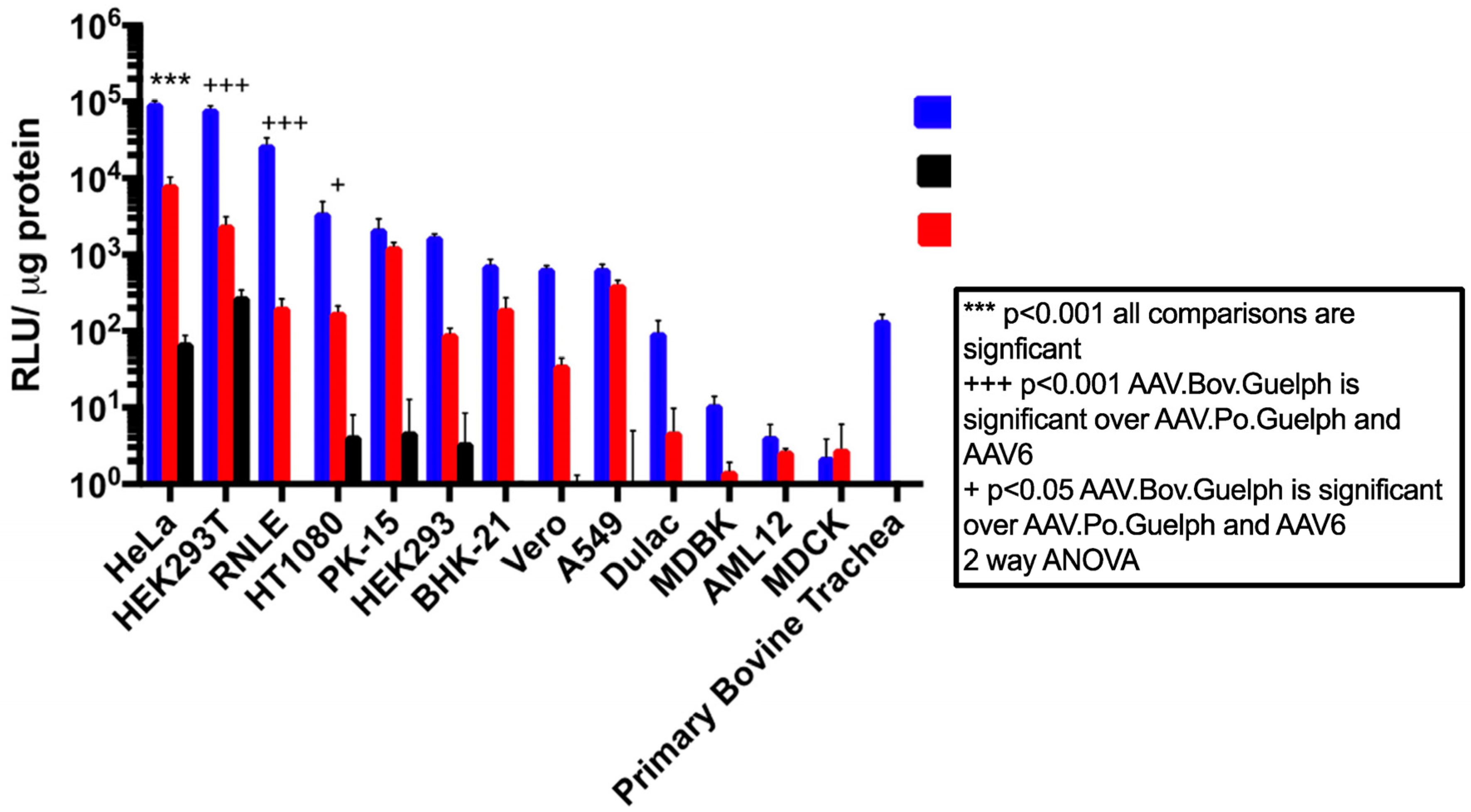
3.4. AAV.Bov.Guelph Transduces Mammalian Cell Lines More Efficiently Than AAV.Po.Guelph
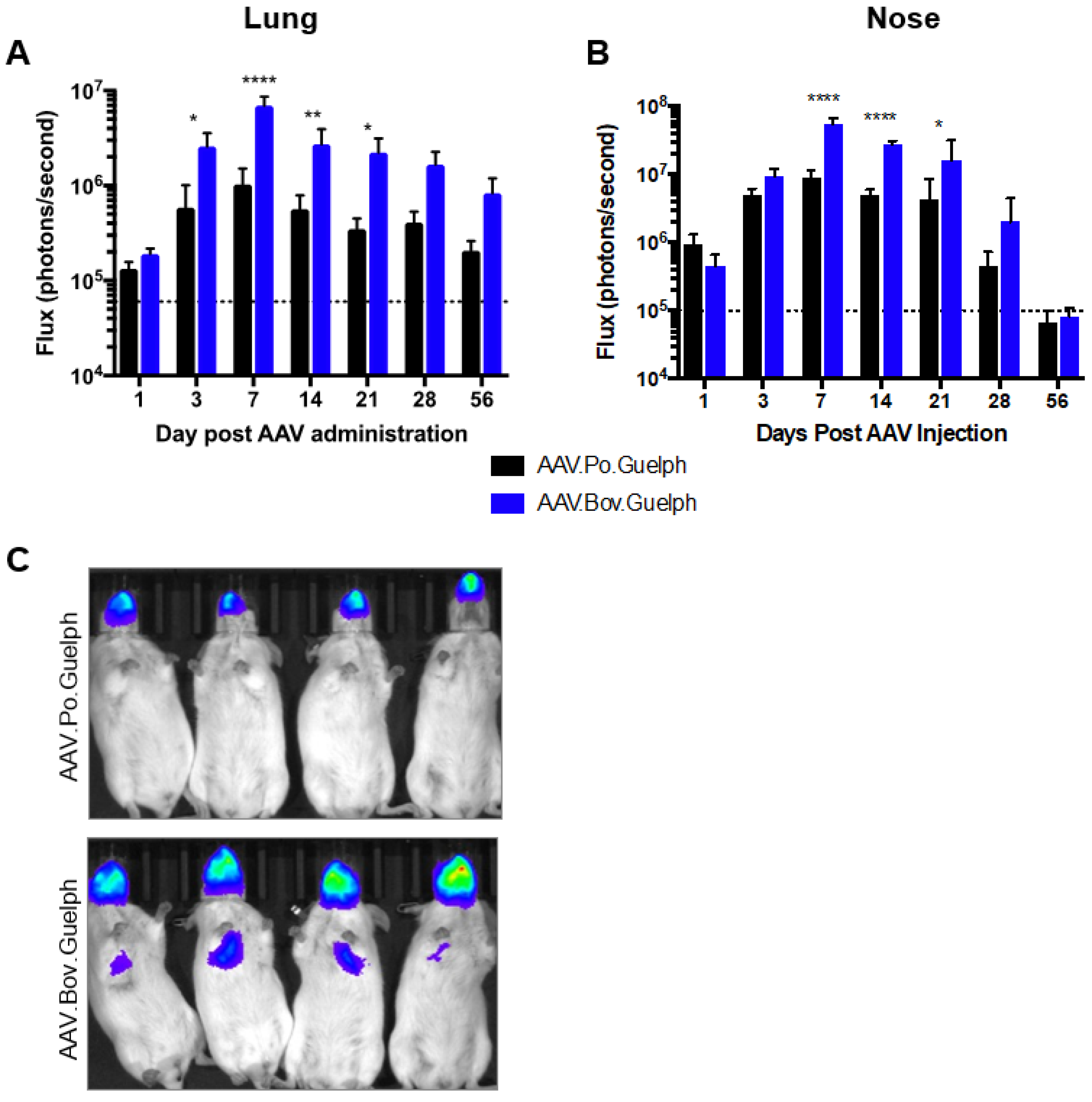
3.5. Analysis of AAV.Po.Guelph and AAV.Bov.Guelph Expressing Luciferase in Mice following Intranasal Administration
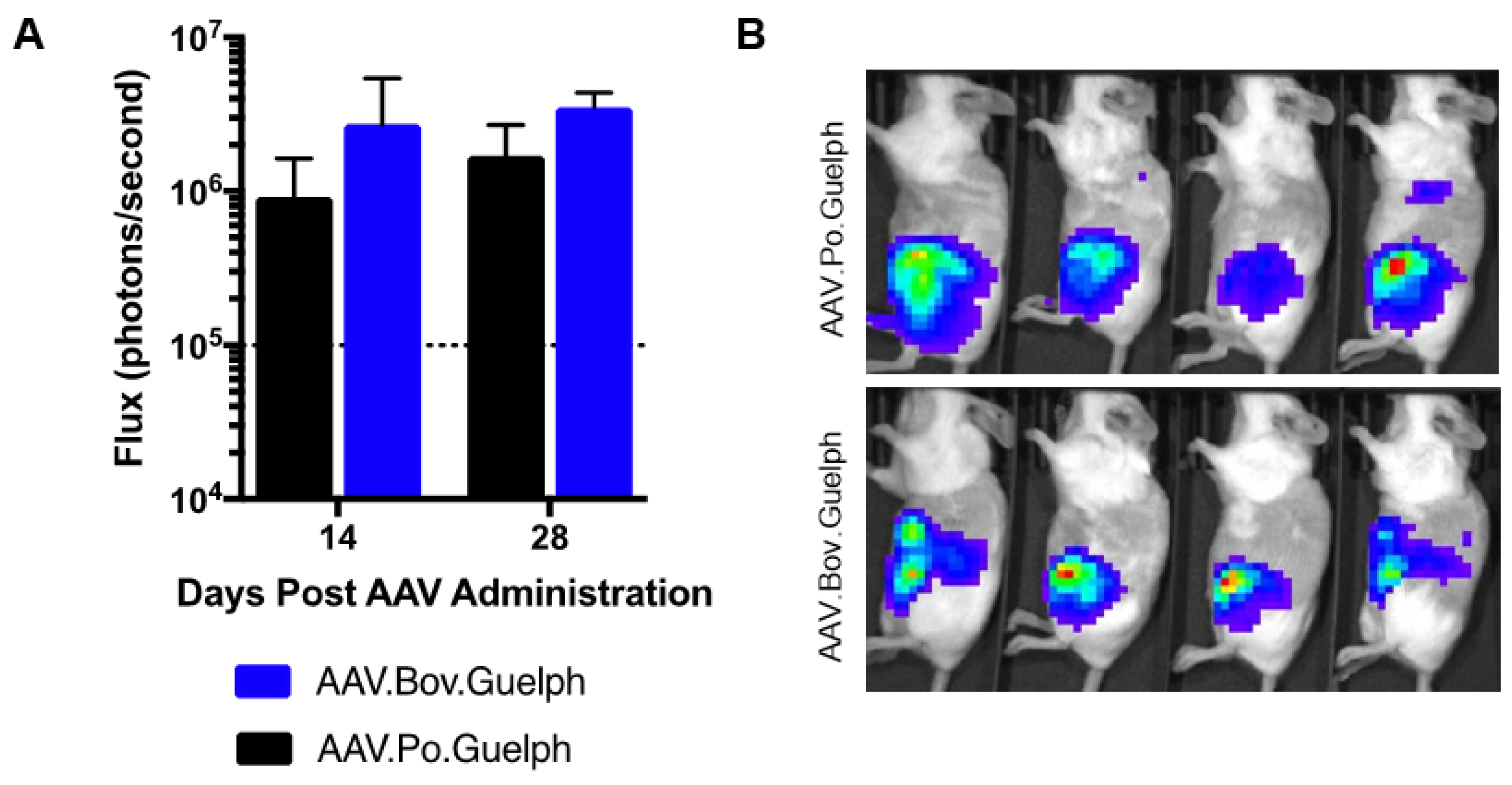
3.6. Analysis of AAV.Po.Guelph and AAV.Bov.Guelph Expressing Luciferase in Mice following Intraperitoneal Administration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuzmin, D.A.; Shutova, M.V.; Johnston, N.R.; Smith, O.P.; Fedorin, V.V.; Kukushkin, Y.S.; van der Loo, J.C.M.; Johnstone, E.C. The clinical landscape for AAV gene therapies. Nat. Rev. Drug Discov. 2021, 20, 173–174. [Google Scholar] [CrossRef] [PubMed]
- Calcedo, R.; Vandenberghe, L.H.; Gao, G.; Lin, J.; Wilson, J.M. Worldwide epidemiology of neutralizing antibodies to adeno-associated viruses. J. Infect. Dis. 2009, 199, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Kay, M.A. From virus evolution to vector revolution: Use of naturally occurring serotypes of adeno-associated virus (AAV) as novel vectors for human gene therapy. Curr. Gene Ther. 2003, 3, 281–304. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Couto, L.B.; Patarroyo-White, S.; Liu, T.; Nagy, D.; Vargas, J.A.; Zhou, S.; Scallan, C.D.; Sommer, J.; Vijay, S.; et al. Effects of transient immunosuppression on adenoassociated, virus-mediated, liver-directed gene transfer in rhesus macaques and implications for human gene therapy. Blood 2006, 108, 3321–3328. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Liu, S.; Ou, L. rAAV immunogenicity, toxicity, and durability in 255 clinical trials: A meta-analysis. Front. Immunol. 2022, 13, 1001263. [Google Scholar] [CrossRef] [PubMed]
- Mendell, J.R.; Connolly, A.M.; Lehman, K.J.; Griffin, D.A.; Khan, S.Z.; Dharia, S.D.; Quintana-Gallardo, L.; Rodino-Klapac, L.R. Testing preexisting antibodies prior to AAV gene transfer therapy: Rationale, lessons and future considerations. Mol. Ther. Methods Clin. Dev. 2022, 25, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.A. Adeno-associated virus: From defective virus to effective vector. Virol. J. 2005, 2, 43. [Google Scholar] [CrossRef]
- Gregorevic, P.; Blankinship, M.J.; Allen, J.M.; Crawford, R.W.; Meuse, L.; Miller, D.G.; Russell, D.W.; Chamberlain, J.S. Systemic delivery of genes to striated muscles using adeno-associated viral vectors. Nat. Med. 2004, 10, 828–834. [Google Scholar] [CrossRef]
- Zolotukhin, S.; Vandenberghe, L.H. AAV capsid design: A Goldilocks challenge. Trends Mol. Med. 2022, 28, 183–193. [Google Scholar] [CrossRef]
- Lisowski, L.; Tay, S.S.; Alexander, I.E. Adeno-associated virus serotypes for gene therapeutics. Curr. Opin. Pharmacol. 2015, 24, 59–67. [Google Scholar] [CrossRef]
- Thwaite, R.; Pagès, G.; Chillón, M.; Bosch, A. AAVrh.10 immunogenicity in mice and humans. Relevance of antibody cross-reactivity in human gene therapy. Gene Ther. 2015, 22, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Goedeker, N.L.; Dharia, S.D.; Griffin, D.A.; Coy, J.; Truesdale, T.; Parikh, R.; Whitehouse, K.; Santra, S.; Asher, D.R.; Zaidman, C.M. Evaluation of rAAVrh74 gene therapy vector seroprevalence by measurement of total binding antibodies in patients with Duchenne muscular dystrophy. Ther. Adv. Neurol. Disord. 2023, 16, 17562864221149781. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Katano, H.; Bossis, I.; Chiorini, J.A. Cloning and characterization of a bovine adeno-associated virus. J. Virol. 2004, 78, 6509–6516. [Google Scholar] [CrossRef] [PubMed]
- Bossis, I.; Chiorini, J.A. Cloning of an avian adeno-associated virus (AAAV) and generation of recombinant AAAV particles. J. Virol. 2003, 77, 6799–6810. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.K.; McFerran, J.B.; McKillop, E.R.; Curran, W.L. Isolation of an adeno associated virus from sheep. Arch. Virol. 1979, 60, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Olson, E.J.; Haskell, S.R.; Frank, R.K.; Lehmkuhl, H.D.; Hobbs, L.A.; Warg, J.V.; Landgraf, J.G.; Wünschmann, A. Isolation of an adenovirus and an adeno-associated virus from goat kids with enteritis. J. Vet. Diagn. Investig. 2004, 16, 461–464. [Google Scholar] [CrossRef]
- Hsi, J.; Mietzsch, M.; Chipman, P.; Afione, S.; Zeher, A.; Huang, R.; Chiorini, J.; McKenna, R. Structural and antigenic characterization of the avian adeno-associated virus capsid. J. Virol. 2023, 97, e0078023. [Google Scholar] [CrossRef]
- Katano, H.; Afione, S.; Schmidt, M.; Chiorini, J.A. Identification of adeno-associated virus contamination in cell and virus stocks by PCR. Biotechniques 2004, 36, 676–680. [Google Scholar] [CrossRef]
- Suchard, M.A.; Weiss, R.E.; Dorman, K.S.; Sinsheimer, J.S. Inferring Spatial Phylogenetic Variation Along Nucleotide Sequences. J. Am. Stat. Assoc. 2003, 98, 427–437. [Google Scholar] [CrossRef]
- Minin, V.N.; Dorman, K.S.; Fang, F.; Suchard, M.A. Dual multiple change-point model leads to more accurate recombination detection. Bioinformatics 2005, 21, 3034–3042. [Google Scholar] [CrossRef]
- Huang, X.; Hartley, A.V.; Yin, Y.; Herskowitz, J.H.; Lah, J.J.; Ressler, K.J. AAV2 production with optimized N/P ratio and PEI-mediated transfection results in low toxicity and high titer for in vitro and in vivo applications. J. Virol. Methods 2013, 193, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.M.; Halbert, C.L.; Miller, A.D. Improved adeno-associated virus vector production with transfection of a single helper adenovirus gene, E4orf6. Mol. Ther. 2000, 1, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Halbert, C.L.; Metzger, M.J.; Lam, S.L.; Miller, A.D. Capsid-expressing DNA in AAV vectors and its elimination by use of an oversize capsid gene for vector production. Gene Ther. 2011, 18, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Halbert, C.; Lam, S.; Miller, A. High-efficiency promoter-dependent transduction by adeno-associated virus type 6 vectors in mouse lung. Hum. Gene Ther. 2007, 18, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.S.; Yao, M.K.; Tran, K.N.; Croyle, M.A.; Strong, J.E.; Feldmann, H.; Kobinger, G.P. Enhanced protection against Ebola virus mediated by an improved adenovirus-based vaccine. PLoS ONE 2009, 4, e5308. [Google Scholar] [CrossRef]
- Aurnhammer, C.; Haase, M.; Muether, N.; Hausl, M.; Rauschhuber, C.; Huber, I.; Nitschko, H.; Busch, U.; Sing, A.; Ehrhardt, A.; et al. Universal real-time PCR for the detection and quantification of adeno-associated virus serotype 2-derived inverted terminal repeat sequences. Hum. Gene Ther. Methods 2012, 23, 18–28. [Google Scholar] [CrossRef]
- Khan, I.F.; Hirata, R.K.; Russell, D.W. AAV-mediated gene targeting methods for human cells. Nat. Protoc. 2011, 6, 482–501. [Google Scholar] [CrossRef]
- Berghuis, L.; Abdelaziz, K.T.; Bierworth, J.; Wyer, L.; Jacob, G.; Karrow, N.A.; Sharif, S.; Clark, M.E.; Caswell, J.L. Comparison of innate immune agonists for induction of tracheal antimicrobial peptide gene expression in tracheal epithelial cells of cattle. Vet. Res. 2014, 45, 105. [Google Scholar] [CrossRef]
- van Lieshout, L.P.; Domm, J.M.; Wootton, S.K. AAV-Mediated Gene Delivery to the Lung. Methods Mol. Biol. 2019, 1950, 361–372. [Google Scholar] [CrossRef]
- Bello, A.; Tran, K.; Chand, A.; Doria, M.; Allocca, M.; Hildinger, M.; Beniac, D.; Kranendonk, C.; Auricchio, A.; Kobinger, G.P. Isolation and evaluation of novel adeno-associated virus sequences from porcine tissues. Gene Ther. 2009, 16, 1320–1328. [Google Scholar] [CrossRef]
- Bello, A.; Chand, A.; Aviles, J.; Soule, G.; Auricchio, A.; Kobinger, G.P. Novel adeno-associated viruses derived from pig tissues transduce most major organs in mice. Sci. Rep. 2014, 4, 6644. [Google Scholar] [CrossRef] [PubMed]
- Puppo, A.; Bello, A.; Manfredi, A.; Cesi, G.; Marrocco, E.; Della Corte, M.; Rossi, S.; Giunti, M.; Bacci, M.L.; Simonelli, F.; et al. Recombinant vectors based on porcine adeno-associated viral serotypes transduce the murine and pig retina. PLoS ONE 2013, 8, e59025. [Google Scholar] [CrossRef]
- Govindasamy, L.; Padron, E.; McKenna, R.; Muzyczka, N.; Kaludov, N.; Chiorini, J.A.; Agbandje-McKenna, M. Structurally mapping the diverse phenotype of adeno-associated virus serotype 4. J. Virol. 2006, 80, 11556–11570. [Google Scholar] [CrossRef] [PubMed]
- Ellis, B.L.; Hirsch, M.L.; Barker, J.C.; Connelly, J.P.; Steininger, R.J.; Porteus, M.H. A survey of ex vivo/in vitro transduction efficiency of mammalian primary cells and cell lines with Nine natural adeno-associated virus (AAV1-9) and one engineered adeno-associated virus serotype. Virol. J. 2013, 10, 74. [Google Scholar] [CrossRef] [PubMed]
- Bowles, D.E.; Rabinowitz, J.E.; Samulski, R.J. Marker rescue of adeno-associated virus (AAV) capsid mutants: A novel approach for chimeric AAV production. J. Virol. 2003, 77, 423–432. [Google Scholar] [CrossRef]
- Gao, G.; Alvira, M.R.; Somanathan, S.; Lu, Y.; Vandenberghe, L.H.; Rux, J.J.; Calcedo, R.; Sanmiguel, J.; Abbas, Z.; Wilson, J.M. Adeno-associated viruses undergo substantial evolution in primates during natural infections. Proc. Natl. Acad. Sci. USA 2003, 100, 6081–6086. [Google Scholar] [CrossRef]
- Rapti, K.; Louis-Jeune, V.; Kohlbrenner, E.; Ishikawa, K.; Ladage, D.; Zolotukhin, S.; Hajjar, R.J.; Weber, T. Neutralizing antibodies against AAV serotypes 1, 2, 6, and 9 in sera of commonly used animal models. Mol. Ther. 2012, 20, 73–83. [Google Scholar] [CrossRef]
- Schmidt, M.; Chiorini, J.A. Gangliosides are essential for bovine adeno-associated virus entry. J. Virol. 2006, 80, 5516–5522. [Google Scholar] [CrossRef]
- Yu, R.K.; Tsai, Y.T.; Ariga, T.; Yanagisawa, M. Structures, biosynthesis, and functions of gangliosides--an overview. J. Oleo Sci. 2011, 60, 537–544. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Ostedgaard, L.; Vermeer, D.; Swaim, W.D.; Karp, P.; Chiorini, J.A. Bovine AAV transcytosis inhibition by tannic acid results in functional expression of CFTR in vitro and altered biodistribution in vivo. Gene Ther. 2012, 19, 576–581. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Chiorini, J.A. AAV transcytosis through barrier epithelia and endothelium. Mol. Ther. 2006, 13, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Bello, A.J.A. Linking the Tropism and Transduction Efficiency of Porcine-Derived Adeno-Associated Viruses to Their Transgene-Mediated Protective Efficacy. 2014. Available online: https://mspace.lib.umanitoba.ca/server/api/core/bitstreams/8ad9bf19-411b-4c60-a8d9-08ef2d1ccdb4/content (accessed on 28 December 2023).
- Zincarelli, C.; Soltys, S.; Rengo, G.; Rabinowitz, J.E. Analysis of AAV serotypes 1–9 mediated gene expression and tropism in mice after systemic injection. Mol. Ther. 2008, 16, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.; Li, J.; Gessler, D.J.; Su, Q.; Wei, Q.; Li, H.; Gao, G. Adeno-associated virus serotype rh.10 displays strong muscle tropism following intraperitoneal delivery. Sci. Rep. 2017, 7, 40336. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Round 1—Initial PCR | Round 2—Hemi-Nested PCR | |
|---|---|---|
| Forward Primer | 5′-ATGNTNATNTGGTGGGA GGA-3′ | 5′-ACCTTNGAACACCAGCAGC-3′ |
| Reverse Primer | 5′-CCANNNGGAATCGCAATGCCAAT-3′ | |
| PCR Components | 20 µL Platinum PCR Supermix | 20 µL Platinum PCR Supermix |
| 1 µL (2 µM) Primers (Each) | 1 µL (2 µM) Primers (Each) | |
| 200 ng DNA | 2 uL of PCR Product | |
| PCR Conditions | 94 °C—2 min | 94 °C—2 min |
| 94 °C—30 s × 40 cycles | 94 °C—30 s × 30 cycles | |
| 55 °C—30 s × 40 cycles | 55 °C—30 s × 30 cycles | |
| 72 °C—30 s × 40 cycles | 72 °C—30 s × 30 cycles | |
| 72 °C—2 min | 72 °C—2 min | |
| Round 1—Initial PCR | Round 2—Hemi-Nested PCR | |
|---|---|---|
| Porcine Capsid Forward Primer | 5′-ATGTCGTTTGTTGATCACC-3′ | 5′-TTACAGGTTGCGGGTGAAGGTAGC-3′ |
| Porcine Capsid Reverse Primer | 5′-TTACAGRTTRCGRGTRAGGTAGC-3′ | |
| PCR Components | 20 µL Platinum PCR Supermix | 20 µL Platinum PCR Supermix |
| 1 µL (2 µM) Primers (Each) | 1 µL (2 µM) Primers (Each) | |
| 200 ng DNA | 4 µL of PCR Product | |
| PCR Conditions | 94 °C—2 min | |
| 94 °C—15 s | ||
| 70 °C (−0.3 °C per cycle) 30 s | 34 cycles | |
| 72 °C—2.5 min | ||
| 94 °C—15 s | ||
| 55 °C—30 s | 14 cycles | |
| 72 °C—2.5 min | ||
| 72 °C—5 min | ||
| Type of Sample | Canine | Feline | Bovine | Porcine | Ovine | Equine | Exotic | Total |
|---|---|---|---|---|---|---|---|---|
| Extraction Method | Qiagen | Qiagen | Qiagen | MagNA Pure | Qiagen | Qiagen | Qiagen | – |
| Number Screened | 10 | 4 | 8 | 74 | 2 | 3 | 3 | 104 |
| Positive PCR Screening | 1 | 0 | 0 | 46 | 2 | 1 | 1 | 51 |
| Change | Position | Variable Region 1 | Role in AAV2 Capsid (Govindasamy et al., 2006) 2 |
|---|---|---|---|
| A to P | 182 | None | NA |
| G to S | 207 | None | NA |
| D to E | 327 | II (262–268) | Transduction |
| Q to H | 385 | III (326–330) | Transduction and A20 neutralization |
| A to Q | 449 | IV (449–468) | HS and IVIG neutralization |
| G to N | 450 | IV (449–468) | HS and IVIG neutralization |
| A to G | 453 | IV (449–468) | HS and IVIG neutralization |
| T to S | 461 | IV (449–468) | HS and IVIG neutralization |
| T to A | 498 | V (487–504) | Transduction, HB, HS, and IVIG neutralization |
| R to K | 500 | V (487–504) | Transduction, HB, HS, and IVIG neutralization |
| D to N | 507 | None | NA |
| RKTV | 529 | VI (525–541) | Transduction, HS, and IVIG neutralization |
| A to T | 544 | VII (544–556) | A20, HS, and IVIG neutralization |
| S to G | 582 | VIII (579–594) | HB, HS, and IVIG neutralization |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, D.L.; van Lieshout, L.P.; Stevens, B.A.Y.; Near, K.J.; Stodola, J.K.; Stinson, K.J.; Slavic, D.; Wootton, S.K. AAV Vectors Pseudotyped with Capsids from Porcine and Bovine Species Mediate In Vitro and In Vivo Gene Delivery. Viruses 2024, 16, 57. https://doi.org/10.3390/v16010057
Yu DL, van Lieshout LP, Stevens BAY, Near KJ, Stodola JK, Stinson KJ, Slavic D, Wootton SK. AAV Vectors Pseudotyped with Capsids from Porcine and Bovine Species Mediate In Vitro and In Vivo Gene Delivery. Viruses. 2024; 16(1):57. https://doi.org/10.3390/v16010057
Chicago/Turabian StyleYu, Darrick L., Laura P. van Lieshout, Brenna A. Y. Stevens, Kelsie J. (Jagt) Near, Jenny K. Stodola, Kevin J. Stinson, Durda Slavic, and Sarah K. Wootton. 2024. "AAV Vectors Pseudotyped with Capsids from Porcine and Bovine Species Mediate In Vitro and In Vivo Gene Delivery" Viruses 16, no. 1: 57. https://doi.org/10.3390/v16010057