
The Chinese Hamster Ovary Cell-Based H9 HA Subunit Avian Influenza Vaccine Provides Complete Protection against the H9N2 Virus Challenge in Chickens
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Optimization and Synthesis of H9 HA Gene
2.3. Construction of Recombinant Plasmids and Transfection
2.4. Screening and Generation of the Stable and H9-HA High-Expression Monoclonal Cell Line
2.5. Expression of HA Proteins of H9N2 AIV
2.6. Protein Purification and Vaccine Preparation
2.7. Immunization and Protection
2.8. HI Assay for Determining the Antibody Titers
3. Results
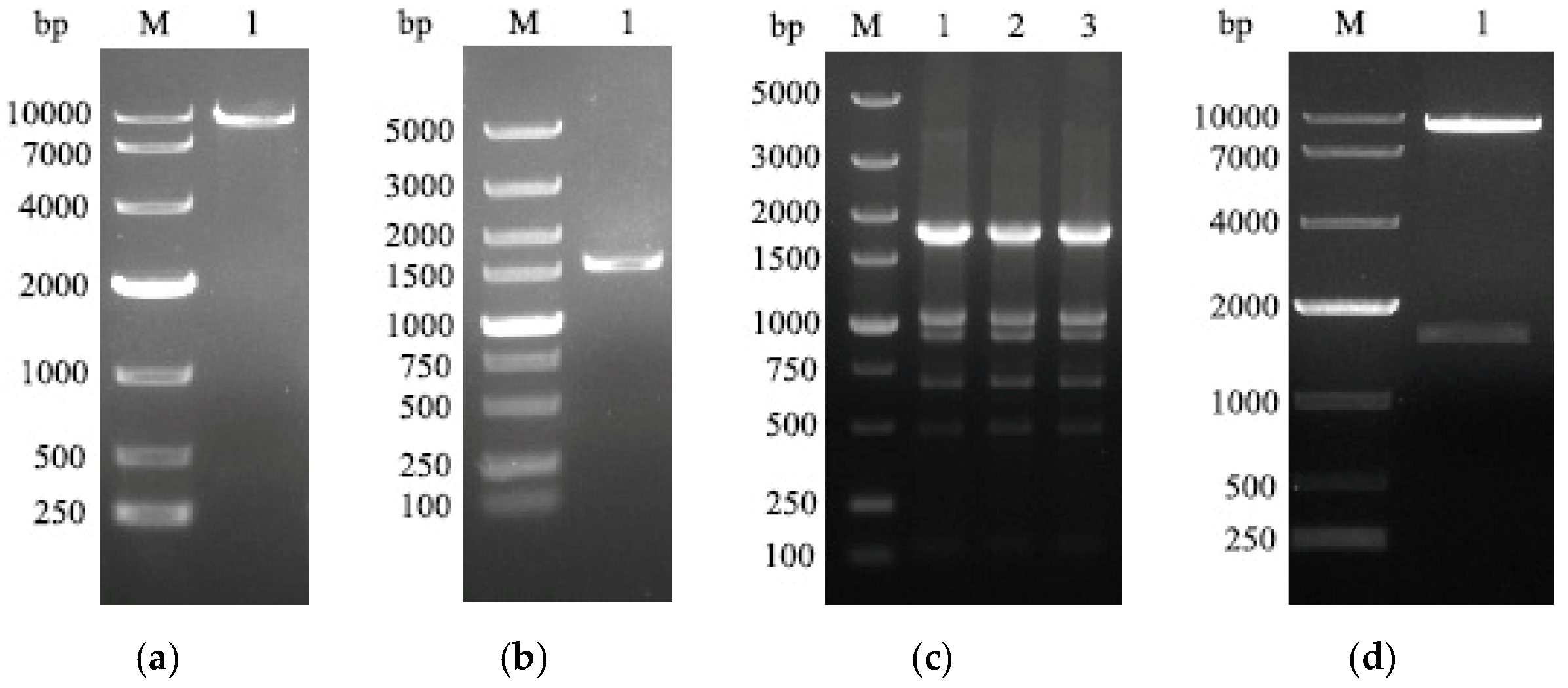
3.1. Construction of Recombinant Plasmids
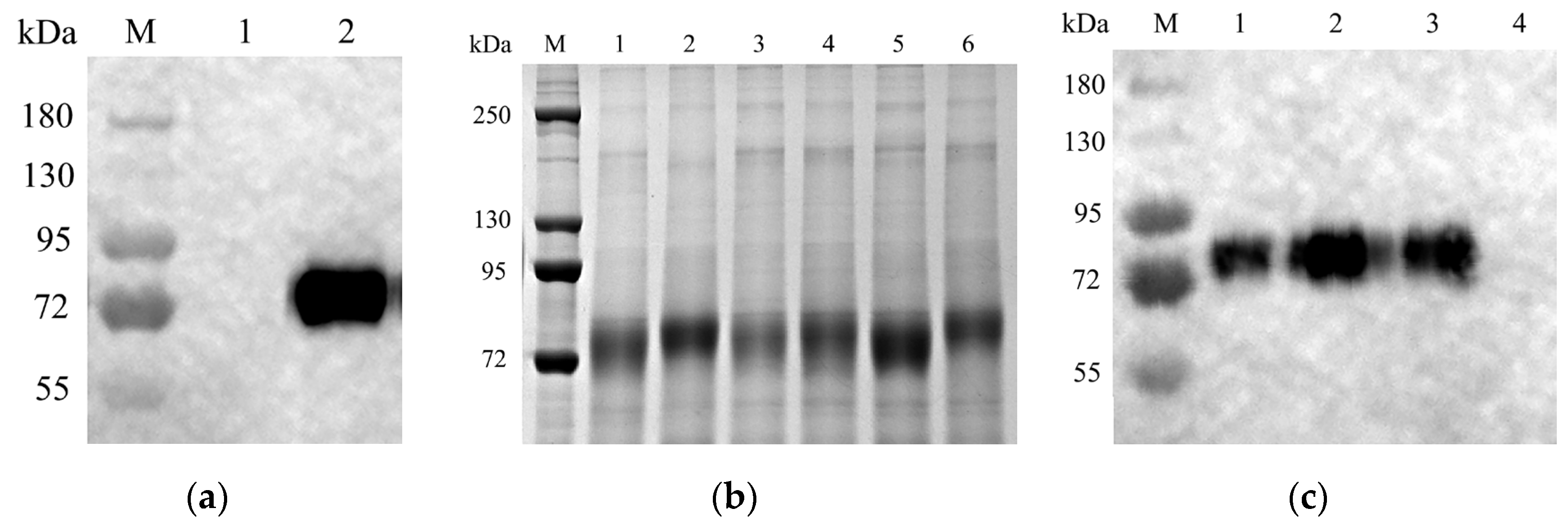
3.2. Expression of HA Protein in CHO Cells
3.3. The H9N2 Virus-Specific Antibodies Were Induced after Being Vaccinated with the H9 Subunit Vaccine Produced in CHO Cells
3.4. The H9 HA Vaccine Provides Complete Protection from the H9N2 AIV Virus Challenge
4. Discussion
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, Y.; Wu, Y.; Tefsen, B.; Shi, Y.; Gao, G.F. Bat-derived influenza-like viruses H17N10 and H18N11. Trends Microbiol. 2014, 22, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wang, Y.; Zhao, C.; Gao, X.; Zhang, Y.; Li, J.; Wang, M.; Zhang, H.; Liu, W.; Wang, C.; et al. Molecular characterization, receptor binding property, and replication in chickens and mice of H9N2 avian influenza viruses isolated from chickens, peafowls, and wild birds in eastern China. Emerg. Microbes Infect. 2021, 10, 2098–2112. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Fu, Z.; Feng, J. The prevalence and prevention of avian influenza subtype H9. In Proceedings of the Tenth Symposium of the Chinese Society of Animal Husbandry and Veterinary Medicine, Avian Diseases Branch, Hangzhou, China, 25 September 2000; pp. 142–145. [Google Scholar]
- Yan, W.; Cui, H.; Engelsma, M.; Beerens, N.; van Oers, M.M.; de Jong, M.C.M.; Li, X.; Liu, Q.; Yang, J.; Teng, Q.; et al. Molecular and Antigenic Characterization of Avian H9N2 Viruses in Southern China. Microbiol. Spectr. 2022, 10, e0082221. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Belser, J.A.; Maines, T.R. Adaptation of H9N2 Influenza Viruses to Mammalian Hosts: A Review of Molecular Markers. Viruses 2020, 12, 541. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zhou, Y.J.; Li, G.X.; Ma, J.H.; Yan, L.P.; Wang, B.; Yang, F.R.; Huang, M.; Tong, G.Z. Genetic diversity of H9N2 influenza viruses from pigs in China: A potential threat to human health? Vet. Microbiol. 2011, 149, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Xu, X.; Liu, Q.; Liang, D.; Li, C.; He, Q.; Jiang, J.; Cui, Y.; Li, J.; Zheng, L.; et al. Evidence of avian-like H9N2 influenza A virus among dogs in Guangxi, China. Infect. Genet. Evol. 2013, 20, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xuan, Y.; Shan, H.; Yang, H.; Wang, J.; Wang, K.; Li, G.; Qiao, J. Avian influenza virus H9N2 infections in farmed minks. Virol. J. 2015, 12, 180. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Schountz, T.; Ma, W. Bat Influenza Viruses: Current Status and Perspective. Viruses 2021, 13, 547. [Google Scholar] [CrossRef]
- Lloren, K.K.S.; Lee, T.; Kwon, J.J.; Song, M.S. Molecular Markers for Interspecies Transmission of Avian Influenza Viruses in Mammalian Hosts. Int. J. Mol. Sci. 2017, 18, 2706. [Google Scholar] [CrossRef]
- Pusch, E.A.; Suarez, D.L. The Multifaceted Zoonotic Risk of H9N2 Avian Influenza. Vet. Sci. 2018, 5, 82. [Google Scholar] [CrossRef]
- Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux, É.; Staubach, C.; Terregino, C.; Aznar, I.; Muñoz Guajardo, I.; et al. Avian influenza overview September–December 2021. Efsa J. 2021, 19, e07108. [Google Scholar] [CrossRef] [PubMed]
- Peacock, T.H.P.; James, J.; Sealy, J.E.; Iqbal, M. A Global Perspective on H9N2 Avian Influenza Virus. Viruses 2019, 11, 620. [Google Scholar] [CrossRef] [PubMed]
- Kimble, J.B.; Sorrell, E.; Shao, H.; Martin, P.L.; Perez, D.R. Compatibility of H9N2 avian influenza surface genes and 2009 pandemic H1N1 internal genes for transmission in the ferret model. Proc. Natl. Acad. Sci. USA 2011, 108, 12084–12088. [Google Scholar] [CrossRef]
- Sorrell, E.M.; Wan, H.; Araya, Y.; Song, H.; Perez, D.R. Minimal molecular constraints for respiratory droplet transmission of an avian-human H9N2 influenza A virus. Proc. Natl. Acad. Sci. USA 2009, 106, 7565–7570. [Google Scholar] [CrossRef]
- Sun, Y.; Qin, K.; Wang, J.; Pu, J.; Tang, Q.; Hu, Y.; Bi, Y.; Zhao, X.; Yang, H.; Shu, Y.; et al. High genetic compatibility and increased pathogenicity of reassortants derived from avian H9N2 and pandemic H1N1/2009 influenza viruses. Proc. Natl. Acad. Sci. USA 2011, 108, 4164–4169. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Hu, J.; Wang, J.; Xu, J.; Cheng, H.; Xu, Y.; Li, Q.; He, D.; Liu, X.; Wang, X.; et al. Reassortant H5N1 avian influenza viruses containing PA or NP gene from an H9N2 virus significantly increase the pathogenicity in mice. Vet. Microbiol. 2016, 192, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Lam, T.T.-Y.; Wang, J.; Shen, Y.; Zhou, B.; Duan, L.; Cheung, C.-L.; Ma, C.; Lycett, S.J.; Leung, C.Y.-H.; Chen, X.; et al. The genesis and source of the H7N9 influenza viruses causing human infections in China. Nature 2013, 502, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Xu, L.; Wang, X.; Liu, X. Current situation of H9N2 subtype avian influenza in China. Vet. Res. 2017, 48, 49. [Google Scholar] [CrossRef]
- Cong, Y.; Zhong, Y.; Sun, Y.; Ding, Z. Progress in the epidemiology of H9N2 subtype of avian influenza virus and its vaccine. China Vet. J. 2017, 37, 386–392. [Google Scholar] [CrossRef]
- Dong, J.; Zhou, Y.; Pu, J.; Liu, L. Status and Challenges for Vaccination against Avian H9N2 Influenza Virus in China. Life 2022, 12, 1326. [Google Scholar] [CrossRef]
- Ren, Z.; Lu, Z.; Wang, L.; Huo, Z.; Cui, J.; Zheng, T.; Dai, Q.; Chen, C.; Qin, M.; Chen, M.; et al. Rapid production of a H9 N2 influenza vaccine from MDCK cells for protecting chicken against influenza virus infection. Appl. Microbiol. Biotechnol. 2015, 99, 2999–3013. [Google Scholar] [CrossRef]
- Petrovsky, N. Comparative Safety of Vaccine Adjuvants: A Summary of Current Evidence and Future Needs. Drug Saf. 2015, 38, 1059–1074. [Google Scholar] [CrossRef] [PubMed]
- Suarez, D.L. Overview of avian influenza DIVA test strategies. Biologicals 2005, 33, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Quan, F.S.; Lee, Y.T.; Kim, K.H.; Kim, M.C.; Kang, S.M. Progress in developing virus-like particle influenza vaccines. Expert Rev. Vaccines 2016, 15, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.S.; Webby, R.J. Traditional and new influenza vaccines. Clin. Microbiol. Rev. 2013, 26, 476–492. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.X.; Li, Z.R.; Zhang, P.J.; Han, J.H.; Di, H.Y.; Qin, J.Y.; Cong, Y.L. A Single Vaccination of Chimeric Bivalent Virus-Like Particle Vaccine Confers Protection Against H9N2 and H3N2 Avian Influenza in Commercial Broilers and Allows a Strategy of Differentiating Infected from Vaccinated Animals. Front. Immunol. 2022, 13, 902515. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Liu, Q.; Niu, S.; Huang, D.; Yan, D.; Teng, Q.; Li, X.; Beerens, N.; Forlenza, M.; de Jong, M.C.M.; et al. Efficacy of a recombinant turkey herpesvirus (H9) vaccine against H9N2 avian influenza virus in chickens with maternal-derived antibodies. Front. Microbiol. 2022, 13, 1107975. [Google Scholar] [CrossRef]
- Ren, W.; Pei, S.; Jiang, W.; Zhao, M.; Jiang, L.; Liu, H.; Yi, Y.; Hui, M.; Li, J. A replication-deficient H9N2 influenza virus carrying H5 hemagglutinin conferred protection against H9N2 and H5N1 influenza viruses in mice. Front. Microbiol. 2022, 13, 1042916. [Google Scholar] [CrossRef]
- Xu, S.; Zhang, B.; Yao, J.; Ruan, W.J.V. A new H9 influenza virus mRNA vaccine elicits robust protective immunity against infection. Vaccine 2023, 41, 2905–2913. [Google Scholar] [CrossRef]
- Boruah, J.L.H.; Venkatesh, G.; Nagarajan, S.; Senthilkumar, D.; Bhatia, S.; Tosh, C.; Kumar, M.; Rai, R.; Tripathi, S.; Shukla, S.; et al. Immunogenicity and cross-protective efficacy of recombinant H5HA1 protein of clade 2.3.2.1a highly pathogenic H5N1 avian influenza virus expressed in E. coli. Microb. Pathog. 2022, 168, 105605. [Google Scholar] [CrossRef]
- Pushko, P.; Tretyakova, I. Influenza Virus Like Particles (VLPs): Opportunities for H7N9 Vaccine Development. Viruses 2020, 12, 518. [Google Scholar] [CrossRef] [PubMed]
- Shirvani, E.; Paldurai, A.; Varghese, B.P.; Samal, S.K. Contributions of HA1 and HA2 Subunits of Highly Pathogenic Avian Influenza Virus in Induction of Neutralizing Antibodies and Protection in Chickens. Front. Microbiol. 2020, 11, 1085. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, J.; Zhang, J.; Ly, H. Advances in Development and Application of Influenza Vaccines. Front. Immunol. 2021, 12, 711997. [Google Scholar] [CrossRef] [PubMed]
- Phan, H.T.; Pham, V.T.; Ho, T.T.; Pham, N.B.; Chu, H.H.; Vu, T.H.; Abdelwhab, E.M.; Scheibner, D.; Mettenleiter, T.C.; Hanh, T.X.; et al. Immunization with Plant-Derived Multimeric H5 Hemagglutinins Protect Chicken against Highly Pathogenic Avian Influenza Virus H5N1. Vaccines 2020, 8, 593. [Google Scholar] [CrossRef] [PubMed]
- Song, S.J.; Shin, G.I.; Noh, J.; Lee, J.; Kim, D.H.; Ryu, G.; Ahn, G.; Jeon, H.; Diao, H.P.; Park, Y.; et al. Plant-based, adjuvant-free, potent multivalent vaccines for avian influenza virus via Lactococcus surface display. J. Integr. Plant Biol. 2021, 63, 1505–1520. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Xie, B.; Gao, T.; Cen, Q.; Ren, Y. Yeast display platform technology to prepare oral vaccine against lethal H7N9 virus challenge in mice. Microb. Cell Fact. 2020, 19, 53. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, Q.; Peng, P.; Li, R.; Li, J.; Wang, X.; Gu, M.; Hu, Z.; Hu, S.; Liu, X.; et al. Baculovirus-derived influenza virus-like particle confers complete protection against lethal H7N9 avian influenza virus challenge in chickens and mice. Vet. Microbiol. 2022, 264, 109306. [Google Scholar] [CrossRef] [PubMed]
- Tihanyi, B.; Nyitray, L. Recent advances in CHO cell line development for recombinant protein production. Drug Discov. Today. Technol. 2020, 38, 25–34. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Cao, N.; Zhu, W.; Chen, Y.; Tan, L.; Zhou, P.; Cao, Z.; Ke, C.; Li, Y.; Wu, J.; Qi, W.; et al. Avian influenza A (H5N1) virus antibodies in pigs and residents of swine farms, southern China. J. Clin. Virol. 2013, 58, 647–651. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, D.; Xu, B.; Liang, W.; Sui, J.; Chen, Y.; Yang, H.; Chen, H.; Wei, P.; Qiao, C. Immune efficacy of an adenoviral vector-based swine influenza vaccine against antigenically distinct H1N1 strains in mice. Antivir. Res. 2017, 147, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Lupiani, B.; Reddy, S.M. The history of avian influenza. Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Gao, X.; Liu, B.; Li, Y.; Liu, W.; Lu, J.; Liu, C.; Xue, R.; Li, X. Mouse adaptation of the H9N2 avian influenza virus causes the downregulation of genes related to innate immune responses and ubiquitin-mediated proteolysis in mice. Med. Microbiol. Immunol. 2020, 209, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Jan, J.T.; Dionne, B.; Butler, M.; Huang, M.H.; Wu, C.Y.; Wong, C.H.; Wu, S.C. Different immunity elicited by recombinant H5N1 hemagglutinin proteins containing pauci-mannose, high-mannose, or complex type N-glycans. PLoS ONE 2013, 8, e66719. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.; Liu, W.C.; Chen, I.C.; Liu, C.C.; Huang, M.H.; Jan, J.T.; Wu, S.C. Recombinant hemagglutinin produced from Chinese Hamster Ovary (CHO) stable cell clones and a PELC/CpG combination adjuvant for H7N9 subunit vaccine development. Vaccine 2019, 37, 6933–6941. [Google Scholar] [CrossRef]
- Dai, L.; Gao, L.; Tao, L.; Hadinegoro, S.R.; Erkin, M.; Ying, Z.; He, P.; Girsang, R.T.; Vergara, H.; Akram, J.; et al. Efficacy and Safety of the RBD-Dimer-Based Covid-19 Vaccine ZF2001 in Adults. N. Engl. J. Med. 2022, 386, 2097–2111. [Google Scholar] [CrossRef]
- Nagy, A.; Lee, J.; Mena, I.; Henningson, J.; Li, Y.; Ma, J.; Duff, M.; Li, Y.; Lang, Y.; Yang, J.; et al. Recombinant Newcastle disease virus expressing H9 HA protects chickens against heterologous avian influenza H9N2 virus challenge. Vaccine 2016, 34, 2537–2545. [Google Scholar] [CrossRef]
- Kim, J.I.; Park, M.S. N-linked glycosylation in the hemagglutinin of influenza A viruses. Yonsei Med. J. 2012, 53, 886–893. [Google Scholar] [CrossRef]
- Tate, M.D.; Job, E.R.; Deng, Y.M.; Gunalan, V.; Maurer-Stroh, S.; Reading, P.C. Playing hide and seek: How glycosylation of the influenza virus hemagglutinin can modulate the immune response to infection. Viruses 2014, 6, 1294–1316. [Google Scholar] [CrossRef]
- Böhm, E.; Seyfried, B.K.; Dockal, M.; Graninger, M.; Hasslacher, M.; Neurath, M.; Konetschny, C.; Matthiessen, P.; Mitterer, A.; Scheiflinger, F. Differences in N-glycosylation of recombinant human coagulation factor VII derived from BHK, CHO, and HEK293 cells. BMC Biotechnol. 2015, 15, 87. [Google Scholar] [CrossRef]
- Croset, A.; Delafosse, L.; Gaudry, J.P.; Arod, C.; Glez, L.; Losberger, C.; Begue, D.; Krstanovic, A.; Robert, F.; Vilbois, F.; et al. Differences in the glycosylation of recombinant proteins expressed in HEK and CHO cells. J. Biotechnol. 2012, 161, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.Q.; Yan, Z.Q.; Zeng, F.G.; Liao, C.T.; Zhou, Q.F.; Qin, J.P.; Xie, Q.M.; Bi, Y.Z.; Chen, F. Isolation and phylogenetic analysis of hemagglutinin gene of H9N2 influenza viruses from chickens in South China from 2012 to 2013. J. Vet. Sci. 2015, 16, 317–324. [Google Scholar] [CrossRef]
- Margine, I.; Palese, P.; Krammer, F. Expression of functional recombinant hemagglutinin and neuraminidase proteins from the novel H7N9 influenza virus using the baculovirus expression system. JoVE 2013, 81, e51112. [Google Scholar] [CrossRef]
- Lee, D.H.; Park, J.K.; Lee, Y.N.; Song, J.M.; Kang, S.M.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. H9N2 avian influenza virus-like particle vaccine provides protective immunity and a strategy for the differentiation of infected from vaccinated animals. Vaccine 2011, 29, 4003–4007. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Cai, Y.; Jiang, Y.; He, X.; Wei, Y.; Yu, Y.; Tian, X. Vaccine adjuvants: Mechanisms and platforms. Signal Transduct. Target. Ther. 2023, 8, 283. [Google Scholar] [CrossRef]
- Bi, Y.; Li, J.; Shi, W. The time is now: A call to contain H9N2 avian influenza viruses. Lancet Microbe 2022, 3, e804–e805. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| PBS | 12 μg | 25 μg | 50 μg | 100 μg | |
|---|---|---|---|---|---|
| Orpharyngeal swabs | 7/7 | 4/7 | 6/7 | 0/7 | 0/7 |
| Cloacal swabs | 4/7 | 0/7 | 0/7 | 0/7 | 0/7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, S.; Nie, Z.; Che, Y.; Shu, J.; Wu, S.; He, Y.; Wu, Y.; Qian, H.; Feng, H.; Zhang, Q. The Chinese Hamster Ovary Cell-Based H9 HA Subunit Avian Influenza Vaccine Provides Complete Protection against the H9N2 Virus Challenge in Chickens. Viruses 2024, 16, 163. https://doi.org/10.3390/v16010163
Zhu S, Nie Z, Che Y, Shu J, Wu S, He Y, Wu Y, Qian H, Feng H, Zhang Q. The Chinese Hamster Ovary Cell-Based H9 HA Subunit Avian Influenza Vaccine Provides Complete Protection against the H9N2 Virus Challenge in Chickens. Viruses. 2024; 16(1):163. https://doi.org/10.3390/v16010163
Chicago/Turabian StyleZhu, Shunfan, Zhenyu Nie, Ying Che, Jianhong Shu, Sufang Wu, Yulong He, Youqiang Wu, Hong Qian, Huapeng Feng, and Qiang Zhang. 2024. "The Chinese Hamster Ovary Cell-Based H9 HA Subunit Avian Influenza Vaccine Provides Complete Protection against the H9N2 Virus Challenge in Chickens" Viruses 16, no. 1: 163. https://doi.org/10.3390/v16010163