1. Introduction
Peste des petits ruminants virus (PPRV) causes severe and highly contagious disease in both domestic and wild small ruminants, such as sheep and goats. PPRV belongs to the family
Paramyxoviridae of the genus
Morbillivirus, and its genome consists of negative-sense single-stranded RNA. The genome encodes six structural proteins, viz. the nucleocapsid protein (N), phosphoprotein (P), fusion protein (F), matrix protein (M), hemagglutinin-neuraminidase proteins (HN), and two non-structural proteins (C and V). In general, the viral proteins have been shown to be involved in the regulation of multiple cellular signaling pathways and work in sync to carry out functions that are critical for the propagation of the viruses in the host [
1]. A recent study has shown that Canine distemper virus (CDV) N protein induced complete autophagy and facilitated viral replication [
2]. Other studies have shown that the Heat shock protein 70K (HSP70K) interacts with the nucleoproteins (N) of CDV and measles morbillivirus [
3,
4].
In higher organisms, cell death can occur as a result of distinct processes, such as autophagy, apoptosis, and necrosis [
5]. Autophagy is defined as an evolutionarily conserved intracellular process leading to the development of the double-membrane structures called autophagosomes. The process is not only involved in the degradation and recycling of misfolded or damaged organelles in the lysosomes [
6,
7], but also provides nutrients to cells under stress [
8,
9]. One of the primary targets of RNA viruses is the proteome associated with the process of autophagy, which may suggest the significance of autophagy regulation throughout the progression of viral infection [
10]. Cellular protein phosphatidylinositiol-3-kinase complex-I is generated by phosphatidylinositol-3-phosphate (PI3P), which determines the sites for phagophore formation upon autophagy induction [
11,
12,
13]. The PI3-kinase can be classified into two complexes: complex-I comprises vacuolar protein sorting 34 (VPS34), vacuolar protein sorting 15 (VPS15), Beclin-1 (BECN1), and ATG14L, while complex-II consists of VPS34, VPS15, BECN1, and ultraviolet (UV) irradiation-resistance-associated gene (UVRAG). Autophagy is modulated at different levels by various upstream signaling pathways regulated by VPS34 kinase [
14]. The recruitment of PI3P-binding proteins occurs by PI3K kinase activity, subsequently leading to the conjugation of ATG8 family proteins, such as LC3, to phosphatidylethanolamine in phagophore membranes [
15,
16]. The autophagic flux or load can be correlated to the levels of lipid-conjugated microtubule-associated proteins 1A/1B light chain 3 or LC3 levels (LC3-II), with the level of LC3-II depicting the rates of LC3-II/autophagosome formation [
17]. LC3 protein localizes in autophagic structures in the cells undergoing autophagy, and it is a mammalian ortholog of yeast ATG8 gene. There are three forms of LC3, i.e., LC3A, LC3B, and LC3C, among which LC3B is the most widely used marker for autophagic processes. LC3B is further processed by different ATG-family proteins into two forms: LC3-I and LC3-II. The conversion of LC3-I to LC3-II is also an important hallmark of autophagy [
18]. Another important and widely used marker to study autophagic flux is the p62 protein, which is also called sequestosome 1 (SQSTM1). It binds to LC3 via a small LC3 interaction region (LIR). Its levels increase when autophagy in inhibited and decrease upon autophagy induction.
A stoichiometric subunit of PI3KC3 called Beclin-1-associated autophagy-related key regulator, or Barkor/ATG14(L), has been found to localize in autophagosomes and direct PI3KC3 to autophagosome. Several studies have described human ATG14 or Barkor protein as a potential functional mammalian ortholog of yeast ATG14. It has been shown to interact with Beclin-1 and the VPS34 class III PI3K complex to initiate autophagic vesicle nucleation [
19,
20,
21,
22]. Several reports have shown that many viruses modulate autophagy pathways of host cells in order to communicate with neighboring cells. For instance, the secretion, distribution, and transmission of infectious and non-infectious viral particles occur via the exosomal or secretory autophagy pathway [
17]. Viruses modulate the autophagy pathway, resulting either in autophagy induction or autophagy inhibition upon infection. The outcome depends on the host cell, virus type, and cellular/environmental conditions [
23]. Viruses code for proteins that can inhibit autophagy progression when autophagy is induced in infected cells as an anti-viral defense mechanism. In contrast, many viruses also have the ability to activate autophagy in host cells, allowing them to multiply and infect other cells [
24,
25]. Several reports have suggested that the autophagy pathway plays a crucial role in PPR virus replication in host cells, and thus, it might help in improving the efficiency of the production of live attenuated PPRV vaccines in a cell culture system by specifically targeting the autophagy pathway [
26]. Autophagy induction by PPRV has been shown to result in the inter-cellular transmission of virus particles via the exosomal pathway. The study showed that TSG101 plays an important role in gathering infectious PPRV RNA genomes into exosomes, thereby aiding in release of virus particles [
27]. The NECTIN 4 pathogen receptor has been revealed in another investigation to promote autophagy and PPRV viral replication. It has been shown that the expression levels of IRGM and HSPA1A were significantly increased following PPRV replication. However, the knockdown of these genes resulted in the impairment of a second wave of autophagy induced by PPRV [
28]. Another study showed that the nucleocapsid protein of PPRV can inhibit interferon production mediated by interaction with IRF3, thereby blocking its activation [
29]. Thus, the present study was undertaken with the aim to describe the role of
Peste des petits ruminants-virus-coded N protein with phosphatidylinositol-3-kinase complex-I in the induction of autophagy.
2. Materials and Methods
2.1. Cell Lines and Cell Culture
Human embryonic kidney (HEK-293 and HEK-293T) and African green monkey (Vero) cells were used. HEK-293, HEK-293T, and Vero cells were maintained in Dulbecco’s minimal essential medium (DMEM; GIBCO, Thermo Fisher Scientific, Waltham, MA, USA; Cat. No. 12430-047) supplemented with 10% fetal bovine serum (FBS; GIBCO, Thermo Fisher Scientific, Waltham, MA, USA; Cat. No. 10270-106) 100 µg/mL penicillin, 100 µg/mL streptomycin antibiotics (Pan Biotech, Aidenbach, Germany; Cat. No. P06-07300), and 1 µg/mL Amphotericin B (HiMedia, Maharashtra, India; Cat. No. A011) in a tissue culture flask with 75 square centimeter growth surface. All the cells were incubated and propagated under standard conditions (37 °C, 5% CO2, and 95% humidity).
2.2. Plasmids and DNA Transfections
The plasmid used to encode the Nucleocapsid (N) gene of PPRV Sungri/96 strain was a kind gift from the Indian Veterinary Research Institute (IVRI), Mukteshwar, Uttarakhand, India. The PPRV-N gene was amplified and sub-cloned into pCDNA-3X-flag vector, downstream of the CMV promoter. The pCR3.1-Beclin-1 (BECN1), pCAG-VPS34, pCAG-NT-VPS15, and pCAG-OSF 2X Strep/FL-ATG14L plasmid constructs were kind gifts from Prof. Beth Levine and Prof. Lyndaa Bennett, University of Texas South Western (UTSW) Medical Center, Texas, USA. Beclin-1 and ATG14L were amplified using specific primers and sub-cloned into eukaryotic HA-pCDNA expression vector. The primers used in the study have been listed in the
Supplementary Table S1. For the expression analysis and further experiments, the cell lines were transfected using 1 mg/mL polyethylenimine (PEI, Polyscience, Inc. Warrington, PA, USA; Cat. No. 23966). For transfections, 2–3 million healthy cells were cultured for 24 h in 100 mm dishes. The plasmids to be transfected were diluted in 1 mL of 150 mM NaCl and 80 µL of PEI solution. The mixture was incubated at room temperature for 15–20 min before adding in the culture dishes. The medium was replaced with fresh medium after 3–4 h of transfection.
2.3. Immunoblotting Analysis
Whole cell lysates were prepared by harvesting the transfected cells and extracting the protein using RIPA lysis buffer (100 mM Tris-Cl, pH 7.5, 1 mM EDTA, 10% glycerol, 1% NP-40, and 150 mM NaCl) containing protease inhibitors (1 µg/mL pepstatin A (Cat. No. P5318), 1 µg/mL leupeptin, 1 µg/mL aprotinin (Cat. No. A1153), and 100 µM PMSF (Cat. No. P-7626) (Sigma Aldrich, St. Louis, MO, USA). Then, cell lysates were obtained by centrifugation at 14,000× g for 15 min at 4 °C. The protein concentration was estimated using Bradford’s reagent (Biorad; Cat No. 5000006), and approximately 80–100 µg of protein was separated on 12% SDS-PAGE gels. The protein bands were transferred onto polyvinylidene fluoride (PVDF) (mdi; Cat. No. SVFX8302XXXX101) membrane at 100 volts for 45–50 min, followed by blocking with 0.5% fat-free milk for 1 h at room temperature. The membrane was then washed with 1X TBST buffer thrice and incubated overnight with primary antibody (1:1000 dilution) at 4 °C. The next day, the membrane was thoroughly washed with 1X TBST thrice for 5 min each and incubated with fluorescently tagged Alexa-fluor goat anti-mice or goat anti rabbit secondary antibodies (1:10,000 dilution) for 1–2 h at room temperature. The fluorescence signal was measured using a Typhoon scanner (GE, Pittsburg, PA, USA) or FLA scanner (Fuji, Minato, Japan).
2.4. Antibodies and Reagents
As PPRV-N was sub-cloned into pCDNA-3X-flag-tagged eukaryotic expression vector, anti-flag monoclonal antibody was used in this study. The following primary antibodies were used in this study: anti-FLAG (Sigma-Aldrich, St. Louis, MO, USA; Cat. No. F3165), anti-HA (Cell Signaling Technology, Danvers, MA, USA; Cat. No. C29F4), anti-GAPDH (Cell Signaling Technology; Cat. No. 14C10), anti-GST (Thermo scientific, Waltham, MA, USA; Cat. No. 8-326), anti-PI3KC3 or anti-VPS34 (Cell Signaling Technology; Cat. No. 4263), anti-PI3KR4 or anti-VPS15 (Cell Signaling Technology; Cat. No. 14580), anti-ATG14L (Cell Signaling Technology; Cat. No. 96752), anti-LC3B (Affinity biosciences, Cincinnati, OH, USA; Cat. No. AF4650), and anti-SQSTM1/p62 (Affinity biosciences; Cat. No. AF5384). The following secondary antibodies were used in this study: Alexa Fluor goat anti-mice IgG 488 (Invitrogen, Carlsbad, CA, USA; Cat. No. A11001), Alexa Fluor goat anti-rabbit IgG 488 (Invitrogen; Cat. No. A11034), Alexa Fluor chicken anti-rabbit IgG 594 (Invitrogen; Cat. No. A21201). The following chemicals and reagents were used in this study: Nonidet P-40 (HiMedia, Maharashtra, India; Cat. No. RM2352), paraformaldehyde (Sigma-Aldrich; Cat. No. F8775), glutaraldehyde (Sigma-Aldrich; Cat. No. G6257), poly-L-lysine (Sigma-Aldrich; Cat. No. P4707), rapamycin (MedChem Express, Monmouth Junction, NJ, USA; Cat. No. HY-10219), chloroquine (Sigma-Aldrich; Cat. No. C6628), wortmannin (MedChem Express; Cat. No. HY-10197), and lysotracker red DND99 (Invitrogen; Cat. No. 7528).
2.5. GST-Tagged Protein Preparation
Only the PPRV-N-pGEX4T3 and pGEX-4T3 vector were transformed into Escherichia coli BL-21-competent cells. Then, they were plated on LB agar plates with ampicillin antibiotic and incubated at 37 °C overnight. A primary culture was propagated from a single colony in approximately 3 mL of Luria-Bertani (LB) broth medium supplemented with ampicillin antibiotic and incubated at 37 °C overnight at 200 rpm in a shaker incubator. The next day, 1% of the primary culture was added to 250 mL of LB medium with ampicillin and incubated at 37 °C with shaking at 200 rpm. The bacterial culture was incubated until the OD600 value reached 0.7, induced with 1 mM of final concentration of isopropyl β-D-1-thiogalactopyranoside (IPTG) (AMRESCO, Cat. No. 0487), and incubated further for 4 h at 37 °C with shaking at 200 rpm for protein over-production. After that, the bacterial cells were pelleted by centrifugation at 3000× g for 10 min at 4 °C, and the supernatant was discarded. The bacterial pellet was then washed twice with 10 mL of ice-cold STE buffer (100 mM NaCl, 10 mM Tris pH 7.5, and 1 mM EDTA) and centrifuged at 3000× g for 10 min at 4 °C. Next, the pellet was resuspended in 1.5 mL of ice-cold NETN buffer (100 mM NaCl, 20 mM Tris pH 8.0, 1 mM EDTA, and 0.5% NP-40) supplemented with protease inhibitors and incubated on ice for 15 min. Then, 75 µL of 1 M Dithiothreitol (DTT; Sigma-Aldrich; Cat. No. D9778) and 900 µL of 10% N-Lauryl sarcosine sodium salt (sarkosyl; Sigma-Aldrich; Cat. No. 137-16-6) prepared in STE buffer was added to the cell suspension, and the cells were lysed by 2 sonication cycles for 4 min each (10 secs ON/OFF pulse). The cell debris was pelleted by centrifugation at 10,000× g for 10 min at 4 °C after sonication. The supernatant was collected in a fresh tube, and 10% Triton X-100 prepared in STE and glutathione sepharose beads (GE Healthcare, Chicago, IL, USA; Cat. No. 17-0756-01) were added. The samples were then nutated at 4 °C overnight. The following day, the beads were centrifuged at 3000× g for 3 min at 4 °C. The beads were washed thrice with NETN buffer supplemented with protease inhibitors and resuspended in 100 μL of the same buffer. To check the expression of GST-tagged proteins, 10 μL of the beads were added to 10 μL of 4X SDS loading dye, and the sample was boiled for 5 min. The samples were then resolved by SDS polyacrylamide gel electrophoresis (SDS-PAGE) for the confirmation of protein expression. Only the concentration of pGEX-4T3 vector and GST-tagged PPRV-N protein was estimated according to the BSA protein standards.
2.6. GST Pulldown Assay
HEK-293T cells were used for the over-expression of PPRV-N or BECN1, or VPS34 or VPS15 or ATG14L. Lysates were prepared by transfecting approx. 10 × 106 HEK-293T cells with PPRV-N or BECN1, or VPS34 or VPS15 or ATG14L expression plasmids and cultured for 48 h. The cells were then harvested, and protein lysate was prepared. Approx. 10% of the protein lysate was taken out from the total lysate, to be used as an input control. To the lysate, GST-tagged protein beads were added, and it was pre-cleared for 2 h at 4 °C in binding buffer supplemented with protease inhibitors. The lysate was then incubated with GST-tagged fusion protein beads. The following day, the beads were washed with the binding buffer (0.5% NP40, 20 mM Tris, 1 mM EDTA, 100 mM NaCl, 80% glycerol, and 5 mM DTT) supplemented with protease inhibitors (1 μg/mL pepstatin, 1 μg/mL aprotinin, 1 μg/mL leupeptin, and 100 μM PMSF) to remove any unbound proteins. The beads were then mixed with 4X SDS dye to extract and denature the bound proteins. The samples were loaded on to a 12% SDS-PAGE gel to resolve the protein bands, followed by Western blotting and probing using a tag-specific primary antibody.
2.7. Co-Immunoprecipitation (co-IP) Assay
HEK-293T cells were co-transfected with PPRV-N and BECN1 or VPS34, or VPS15, or ATG14L expression plasmids for 48 h. The transfected cells were harvested, and lysates were prepared using RIPA lysis buffer supplemented with protease inhibitors. From the total lysate, 5% of the lysate was taken out to be used as the input control. For each sample, 450 µL of lysate was incubated with 8 µg of primary antibody on nutation at 4 °C overnight. The next day, 50 µL of protein A (Thermo Scientific; Cat. No. 10-1141) and G beads (Thermo Scientific; Cat. No. 10-1242) were added in a ratio of 3:1, and the samples were kept again on nutation at 4 °C for 4 h. The beads were then centrifuged at 3000× g and washed thrice with RIPA (IP) buffer supplemented with protease inhibitors. The beads were then finally resuspended in 50 µL of IP buffer, and samples were prepared for SDS-PAGE by adding SDS loading dye. The samples were resolved on SDS-PAGE followed by immunoblotting.
2.8. Confocal Immunofluorescence Microscopy
Coverslips were treated with poly-L-lysine to be used in the experiment. Each treated coverslip was placed into each well of a 6-well plate. The HEK-293T cells were seeded in the 6-well plates with coverslips. After the cells were adhered to the coverslips, they were co-transfected with the expression plasmids of PPRV-N and BECN1 or VPS34, or VPS15 or ATG14L and incubated for 24 h. The cells were then washed gently with 1X PBS buffer three times after removing the medium. To fix the cells, 1 mL of methanol/acetone (1:1) was added to each well and incubated at −20 °C for 15 min. The cells were then air-dried and blocked with 1% fish skin gelatin (AMRESCO, Solon, OH, USA; Cat. No. M319) prepared in PBS. The coverslips were washed three times with 1X PBS buffer and incubated with specific primary antibodies at 4 °C overnight in a moist chamber. The next day, the coverslips were again washed with 1X PBS buffer three times and incubated with fluorescently labelled secondary antibodies for 2 h at room temperature in dark. Finally, the coverslips were washed again three times, and the cells were treated with DAPI (4′, 6-diamino-2-phenylindole; Sigma-Aldrich; Cat. No. D9542) for 30 s. The unbound DAPI was removed by washing three times with 1X PBS buffer. The coverslips were mounted onto the clean glass slides using fluoromount and visualized under a confocal microscope (Leica TCS SP8 system).
2.9. Quantitative Real-Time PCR (qPCR)
The total RNAs were extracted from the HEK-293 cells using an RNA isolation kit (Invitrogen, Carlsbad, CA, USA) as per the manufacturer’s protocol. The cDNAs were synthesized using a SuperScript First-Strand Reverse Transcription Kit (Agilent technologies, CA, USA) as per the manufacturer’s protocol. The primer pairs listed in the
Supplementary Table S2 were used in RT-PCR for the amplification of TFEB, E2F1, BECN1, FOXO, GATA4, and ZKSCAN genes. Normalization was performed using GAPDH as an internal control. The experiments were performed independently in triplicates, and all experiments were carried out with at least three repeats.
2.10. Transmission Electron Microscopy
The visualization and evaluation of autophagosomes by transmission electron microscopy was performed at Sophisticated Analytical Instrumentation Facility (SAIF), AIIMS, New Delhi, India. In brief, the HEK-293T cells were mock-transfected or transfected with PPRV-N expression plasmid for 48 h. Rapamycin (200 nM)-treated HEK-293T cells were used as positive control. The cells were then harvested and centrifuged at 1000× g for 3 min. The cell pellet was washed with 0.1 M phosphate buffer pH 7.4. The cells were then fixed with 2% paraformaldehyde and 2.5% glutaraldehyde in 0.1 M phosphate buffer at 4 °C for 4–6 h. Next, the cells were washed and postfixed in 1% osmium tetroxide (OSO4) for 1 h at 4 °C. The samples were dehydrated in acetone, infiltrated and embedded in araldite CY212 (TAAB, Berkshire, UK). Ultrathin sections (1 µm) were cut using an ultramicrotome (Leica Ultracut UC7, Vienna, Austria) and mounted and adhered onto clean glass slides. Later, they were stained with aqueous toluidine blue. The sections were observed under the light microscope to check the quality of the tissue fixation. For electron microscope examination, thin sections of grey-silver color interference (70–80 nm) were cut and mounted onto 300 mesh-copper grids. The sections were stained with aqueous uranyl acetate and alkaline lead citrate for 10 min in each. They were washed gently with distilled water and observed under a Tecnai G2 20 high resolution transmission electron microscope (Fei Company, Eindhoven, The Netherlands) at an operating voltage of 200 kV.
2.11. Autophagic Flux Assay
For the autophagic flux assay, the HEK-293T cells were seeded in 6-well culture dishes, and cells were either transfected or mock transfected with PPRV-N expression plasmid, or treated with 200 nM of rapamycin for 48 h. The transfections or mock transfections with PPRV-N expression plasmid or rapamycin treatment was completed in two sets. One set of cells was used as control without chloroquine treatment, and the second set of cells was treated with chloroquine (20 µM) for 2 h. After that, cells were harvested and lysed using RIPA lysis buffer, and 100 µg of total protein lysate was resolved on SDS-PAGE, followed by immunoblotting using the appropriate primary antibodies to monitor autophagic flux. For the detection of the inhibition of autophagy, the HEK-293T cells were pretreated with 200 nM of wortmannin for 6 h, followed by transfection with PPRV-N expression plasmid and further incubation for 48 h at standard culture conditions. Later, the cells were harvested and lysed, and resolved on SDS-PAGE, followed by Western blotting with specific antibodies for autophagy.
2.12. Lysosome Staining
The HEK-293T cells were plated onto clean poly-L-lysine-treated coverslips placed in 6-well culture dishes. The cells were transfected or mock transfected with PPRV-N expression plasmid and culture for 48 h. The cells were gently washed with 1X PBS buffer and fixed with 3.2% of paraformaldehyde for 30 min at room temperature. For the lysosome staining, the cells were washed again with 1X PBS buffer and incubated with 100 nM lysotracker red DND-99 dye in DMEM medium for 30 min at 37 °C. Finally, the cells were counter-stained with DAPI after washing with 1X PBS buffer, mounted onto a clean glass slide, and finally visualized under a Carl Ziess Apotome 2 Axio Observer microscope (Göttingen, Germany) for lysosome detection. A total of three microscopic fields were observed for each sample, and the average red color intensity of lysotracker red dye was plotted on the graph. The experiments were performed independently in duplicates.
2.13. Imaging and Statistical Analysis
Confocal microscopy was performed using a Leica TCS SP8 confocal microscope, and Leica application suite advanced fluorescence software was used for the visualization of immune-stained cells and their quantification. The protein bands were quantified using ImageJ software (NIH). All the experiments were performed independently at least in duplicates or triplicates. Statistical analysis and the plotting of graph were performed using Microsoft Excel (MS office student 2016). The significance was determined by paired Student’s t-test. Values of *** p < 0.0005, ** p < 0.005, and * p < 0.05 were considered statistically significant.
4. Discussion
Peste des petits ruminants virus causes a highly contagious disease of great economic importance. The disease is considered to be a potential threat to small ruminants around the globe [
40]. Upon infection of the intracellular pathogens, host cells can induce autophagy as an innate immune response to control the spread and transmission of the disease. Several studies have shown that autophagy can play an important role in both the protective function for host cell survival under stress, and also in the cellular defense mechanism against the pathogen [
41,
42]. VPS34 protein is considered to be the core subunit of the class III PI3K-complex-I, and its main function includes the activation of the autophagy pathway. VPS15 is believed to anchor VPS34 to the membrane through N-terminal myristylation and acts as a positive regulator of autophagy [
43,
44]. Beclin-1 is another regulatory subunit of this complex that plays an important role in binding to other modulators of autophagy. ATG14L is another regulatory protein present at the peripheral ER and forms puncta during autophagosome formation [
22,
45].
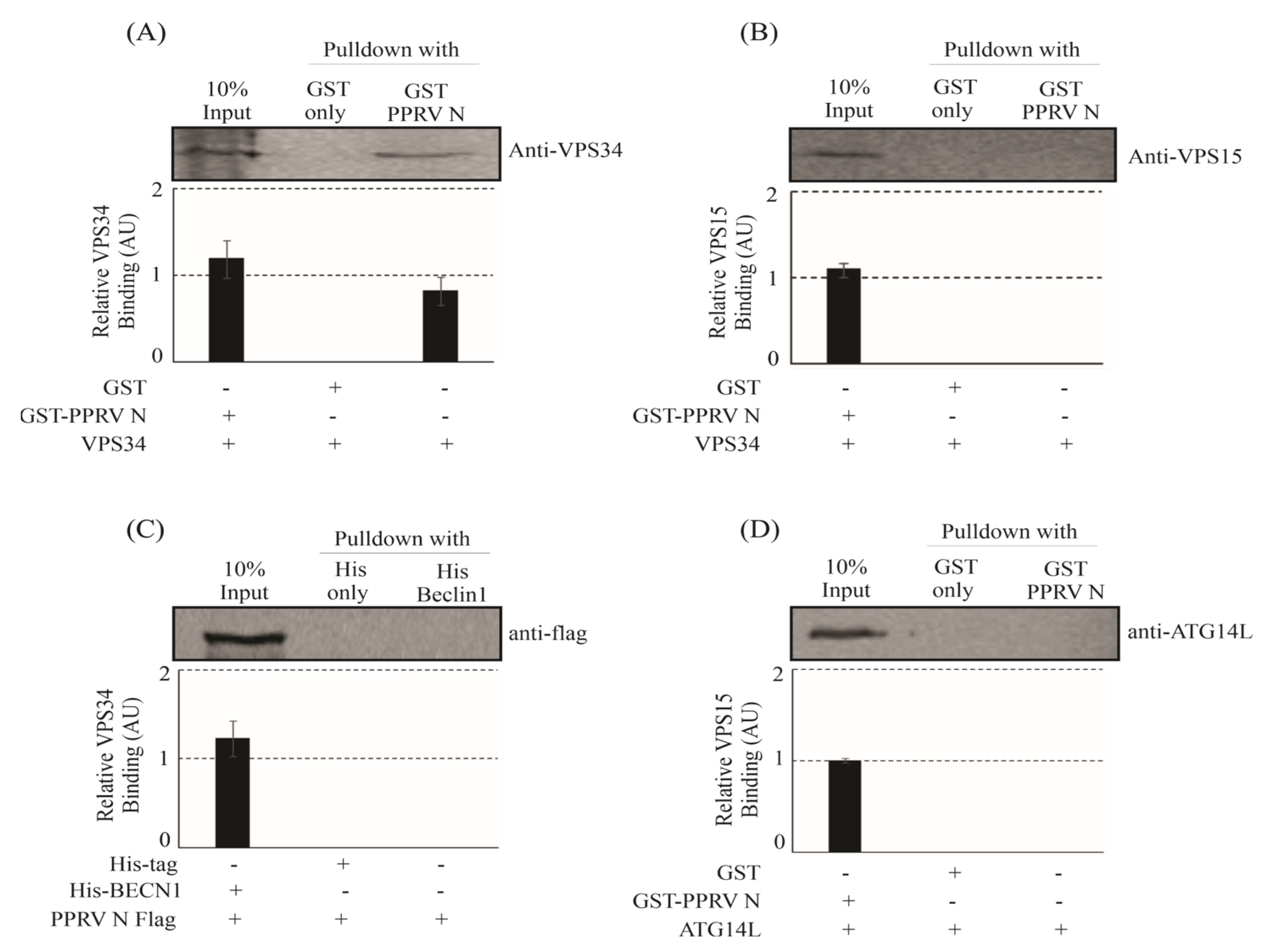
Our study has now shown that PPRV-coded structural protein N can interact with the components of phosphatidylinositol-3-kinase complex-I in vitro (
Figure 1) as well as in vivo (
Figure 2). Previous reports have shown that ATG14L/Barkor interacts with Beclin-1 and VPS34, and together, they play an important role in the initiation of the autophagy process [
19,
20,
21,
22]. Beclin-1 is known as an autophagy marker and is known to interact with various proteins of viruses. Earlier, studies have also shown that the Nef protein of HIV interacts directly with Beclin-1, thereby inhibiting the autophagy pathway, which was further confirmed by assessing the mRNA expression levels [
46]. A study had shown that the NS2 protein of human respiratory syncytial virus induces autophagy through the modulation and stabilization of Beclin-1 protein by impairing ISGylation to overcome autophagy inhibition [
47]. Studies had also shown that HCV non-structural protein NS4B interacts and forms a complex with Rab5 and VPS34 that leads to autophagy induction [
48]. The HCV non-structural protein 5A interacts with ATG14L and ATG14L-Beclin1-VPS34-VPS15 complex, which is necessary for autophagosome formation, in a RACK1-dependent manner. The interaction is important for the formation of HCV double-membrane vesicle and viral replication [
49]. Another study showed nuclear receptor binding factor 2 (Nrbf2) interacts with ATG14L-containing Beclin-1-VPS34 complex and plays an important role in autophagic regulation [
50]. VPS34 acts as a catalytic subunit, which exists in a heterodimeric structure with the regulatory subunit, VPS15 [
51]. Therefore, VPS34 is considered to be the core of the PI3K complex-I, and the interaction of PPRV-N with VPS34 signifies its probable contribution in the initiation of the autophagy pathway. Our study now shows the evidence for a stable interaction between PPRV-N and PI3kinase complex-I protein components, and that this interaction may be simply involved in the initiation of autophagosome biosynthesis. Thus, the interaction of PPRV-N with individual components of phosphatidylinositol-3-kinase complex-I specifies the critical role of PPRV-N in initiating the formation of autophagosomes and hence autophagy.
We also investigated the sub-cellular localization of these different components of phosphatidyl-3-kinase complex-I in cells expressing PPRV-N viral protein. Previous research has demonstrated that in RPE-1 cells, ectopically expressed mCherry-Nrbf2 and Atg14L-EGFP predominantly colocalized on punctate structures under both nutrient-rich and serum-starved conditions [
50]. The Rab7 protein, which is essential for transport between early and late endosomes, colocalizes with p150/VPS15 on punctate perinuclear vesicles. In order to form a complex on late endosomes, it also colocalizes with hVPS34 [
52]. Our findings showed that the PPRV-N protein and the host proteins involved in autophagy, largely colocalizes in the cytoplasm without having any remarkable effect on their intracellular localization. The presence of PPRV-N protein does not cause any change in the expression and localization of the host proteins. The fact that PPRV-N and PI3-kinase complex proteins exist stably inside the same sub-cellular compartment could be important for their stable interaction, which would be critical for the role of these interactions in the modulation of the autophagy pathway.
The process of autophagy is very much regulated and involves the modulation of the transcription of several transcriptional factors. The first observation related to the role of transcriptional regulation in autophagy induction was made in yeast by Kirisako group, where they reported that nitrogen starvation in yeast can trigger autophagy by up-regulating the Apg8p autophagy gene, which is also a homolog of mammalian LC3 [
53]. In past, several studies have reported the up-regulation or down-regulation of several autophagy-related genes, which correlate with the induction of autophagy. Our study also showed that the transcription of several autophagy inducers, like TFEB, E2F1, BECN1, and FOXO, increased significantly in the presence of PPRV-N protein; while the transcription of autophagic repressors, like ZKSCAN and GATA, were either reduced or remained unaffected. Therefore, the real time data implies that PPRV-N ectopic expression can modulate the transcription of genes involved in the induction and repression of autophagy.
For monitoring the autophagic flux in the cultured cells, LC3 has been used widely as an autophagosome marker. In our study, there was a significant increase in the accumulation of LC3-II and a simultaneous reduction in the levels of SQSTM1/p62 in PPRV-N-expressing or rapamycin-treated cells, both in the presence and absence of chloroquine. These findings are correlate with the significant contribution of PPRV-N in triggering the formation of autophagosomes mediated by phsophotidylinositol-3-kinase complex-I, thereby leading to enhanced autophagic activity. In contrast, low basal levels of LC3-II and SQSTM1/p62 were detected in the mock transfected HEK-293T cells, indicating the basal level of autophagy. Our study further showed that the initiation of autophagy in PPRV-N-expressing cells was blocked after treatment with wortmannin, as indicated by reduced accumulation of LC3-II, resulting in reduced autophagic flux. On the other hand, there were no significant changes in the mock transfected or untreated cells, implying the direct role of PPRV-coded N protein in the induction of autophagy. Additionally, lysotracker dyes can also be employed as markers to visualize lysosomes and the autophagic flux. The number of red-colored puncta were found to be increasing with the increasing expression of PPRV-N protein in the presence of chloroquine, which suggests that the effect of PPRV-N on punctate formation is dose-dependent. The cells with larger number of red puncta are indicative of increased autophagic activity, thereby further confirming the role of PPRV-N in the induction of the autophagy process.
Autophagosomes are defined as double-membrane structures and can be visualized at the ultrastructural level using transmission electron microscopy (TEM). A study had employed TEM to show that Dengue Virus-2 can induce autophagosome formation, as shown by ultra-structure images [
54]. Some studies have shown that PPR virus infection induces two distinct waves of autophagy, which have been confirmed via the ultrastructural analysis of EECs infected with UV-irradiated PPRV or PPRV using TEM [
28]. Our study investigates the presence of autophagosomes in PPRV-N-expressing HEK-293T cells and also in rapamycin-treated cells. Many double-membrane autophagosomes and single-membrane autolysosomes were found within these PPRV-N-expressing and rapamycin-treated cells. Completely or partially degraded organelles were seen in the cells with active autophagy. Therefore, this confirms that PPRV-N promotes autophagy in cultured cells. Our study therefore provides evidence for the critical role of PPRV-N protein in the induction of autophagy, which is likely to be mediated by phosphatidylinositol-3-kinase complex-I (
Figure 8).

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







