
BA.1/BA.5 Immunogenicity, Reactogenicity, and Disease Activity after COVID-19 Vaccination in Patients with ANCA-Associated Vasculitis: A Prospective Observational Cohort Study

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Anti-SARS-CoV-2 S1 and Nucleocapsid IgG Chemiluminescent Immunoassay
2.3. Detection of SARS-CoV-2-Specific Surrogate-Neutralizing Antibodies
2.4. Bead-Based Multiplex Assay for the Detection of Antibodies to Various SARS-CoV-2 Epitopes
2.5. Live-Virus Neutralization against the Omicron Subtypes BA.1 and BA.5
2.6. Monitoring of Reactogenicity and Disease Activity
2.7. Statistics
3. Results
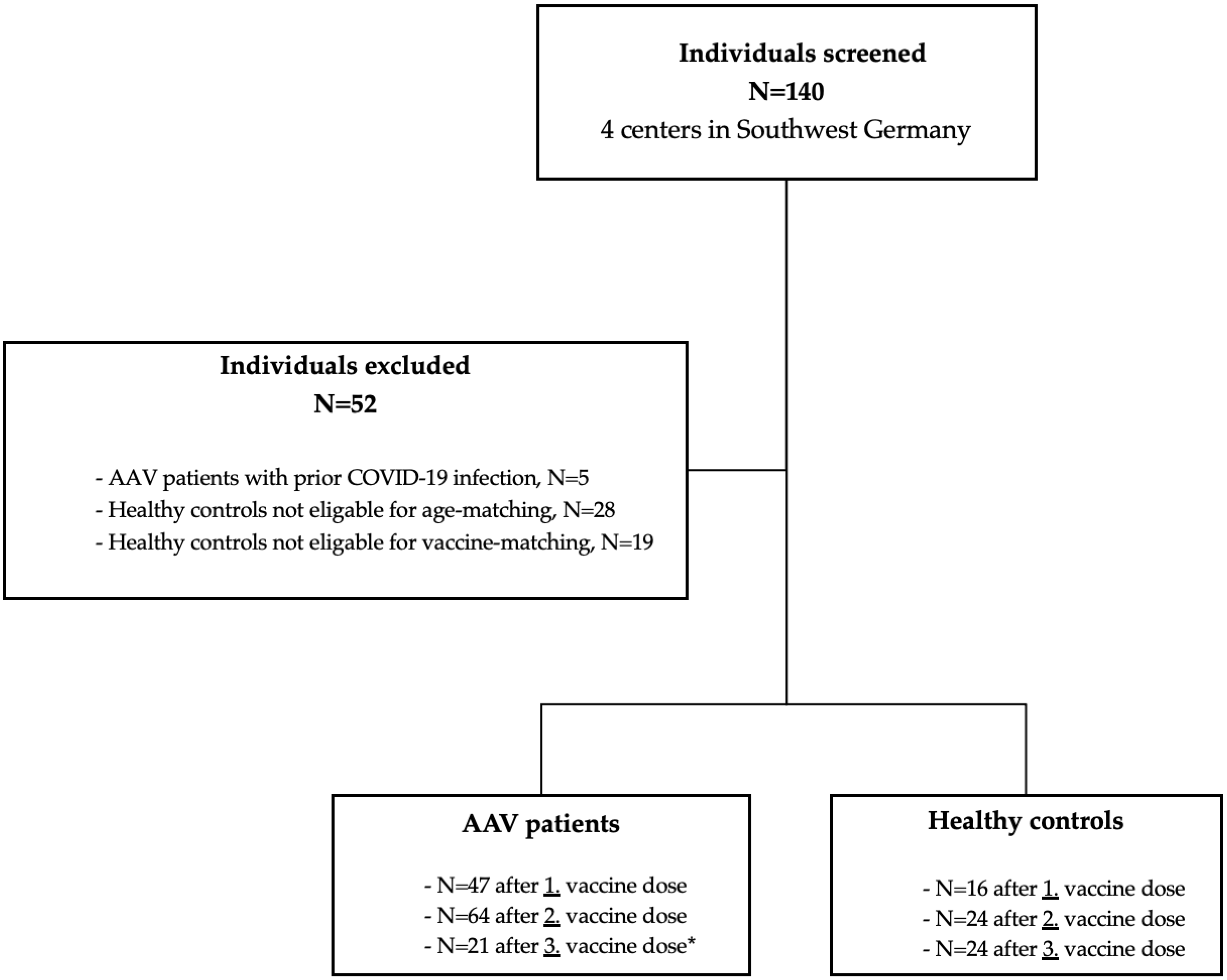
3.1. Study Population and Baseline Characteristics
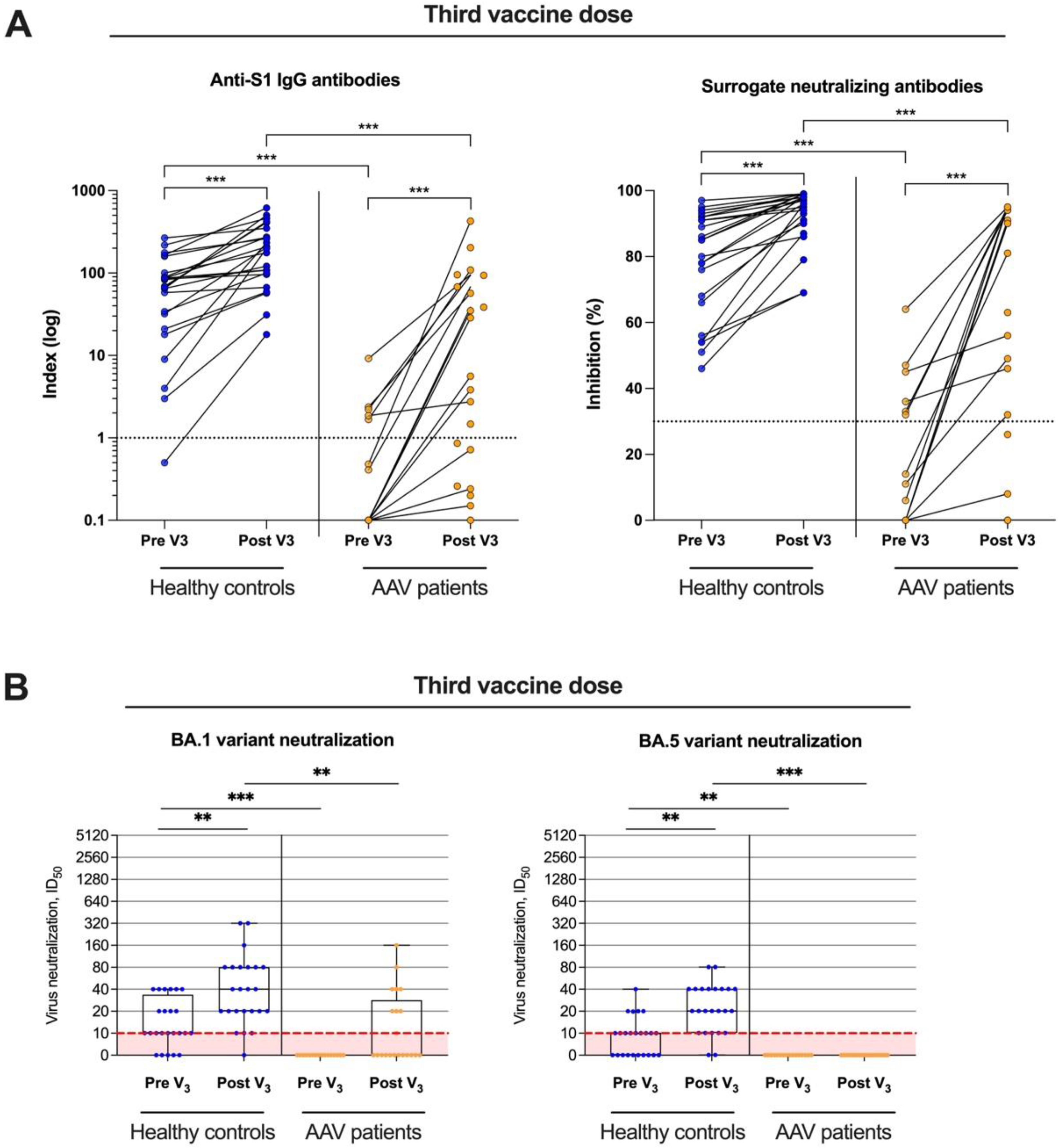
3.2. Anti-S1 IgG, Surrogate-Neutralizing Antibodies, and Antibodies to Various SARS-CoV-2 Epitopes in AAV Patients and Healthy Controls
3.3. Live-Virus Neutralization against the Omicron Subtypes BA.1 and BA.5 before and after Booster Vaccination
3.4. Longitudinal Monitoring of Reactogenicity and Disease Activity of ANCA-Associated Vasculitis after a First, Second, and Third SARS-CoV-2 Vaccine Dose
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Msemburi, W.; Karlinsky, A.; Knutson, V.; Aleshin-Guendel, S.; Chatterji, S.; Wakefield, J. The WHO Estimates of Excess Mortality Associated with the COVID-19 Pandemic. Nature 2023, 613, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Hassan, I.; Fernandes, G.; Mukaigawara, M.; Sridhar, D. Lessons from COVID-19 Must Be Learned before the next Outbreak. Nat. Med. 2023; ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, B.; Zhang, S.; Huang, N.; Zhao, T.; Lu, Q.; Cui, F. Differences in Incidence and Fatality of COVID-19 by SARS-CoV-2 Omicron Variant versus Delta Variant in Relation to Vaccine Coverage: A World-wide Review. J. Med. Virol. 2023, 95, e28118. [Google Scholar] [CrossRef] [PubMed]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing Antibody Levels Are Highly Predictive of Immune Protection from Symptomatic SARS-CoV-2 Infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Kang, M.; Zhao, N.; Zhuang, Y.; Li, S.; Song, T. Comprehensive Narrative Review of Real-World COVID-19 Vaccines: Viewpoints and Opportunities. Med. Rev. 2022, 2, 169–196. [Google Scholar] [CrossRef]
- Smits, P.D.; Gratzl, S.; Simonov, M.; Nachimuthu, S.K.; Cartwright, B.M.G.; Wang, M.D.; Baker, C.; Rodriguez, P.; Bogiages, M.; Althouse, B.M.; et al. Risk of COVID-19 Breakthrough Infection and Hospitalization in Individuals with Comorbidities. Vaccine 2023, 41, 2447–2455. [Google Scholar] [CrossRef]
- Yusof, M.Y.M.; Arnold, J.; Saleem, B.; Vandevelde, C.; Dass, S.; Savic, S.; Vital, E.M.; Emery, P. Breakthrough SARS-CoV-2 Infections and Prediction of Moderate-to-Severe Outcomes during Rituximab Therapy in Patients with Rheumatic and Musculoskeletal Diseases in the UK: A Single-Centre Cohort Study. Lancet Rheumatol. 2023, 5, e88–e98. [Google Scholar] [CrossRef]
- Montoya, A.; Wen, K.; Travers, J.L.; Rivera-Hernandez, M.; White, E.; Mor, V.; Berry, S.D. Resident Factors Associated with Breakthrough SARS-CoV-2 Infections. J. Am. Med. Dir. Assoc. 2023, 24, 901–905. [Google Scholar] [CrossRef]
- Tan, S.T.; Kwan, A.T.; Rodríguez-Barraquer, I.; Singer, B.J.; Park, H.J.; Lewnard, J.A.; Sears, D.; Lo, N.C. Infectiousness of SARS-CoV-2 Breakthrough Infections and Reinfections during the Omicron Wave. Nat. Med. 2023, 29, 358–365. [Google Scholar] [CrossRef]
- Benning, L.; Morath, C.; Bartenschlager, M.; Nusshag, C.; Kälble, F.; Buylaert, M.; Schaier, M.; Beimler, J.; Klein, K.; Grenz, J.; et al. Neutralization of SARS-CoV-2 Variants of Concern in Kidney Transplant Recipients after Standard COVID-19 Vaccination. Clin. J. Am. Soc. Nephrol. 2021, 17, 98–106. [Google Scholar] [CrossRef]
- Benning, L.; Morath, C.; Bartenschlager, M.; Reineke, M.; Töllner, M.; Nusshag, C.; Kälble, F.; Reichel, P.; Schaier, M.; Klein, K.; et al. Neutralizing Antibody Activity against the B.1.617.2 (Delta) Variant 8 Months after Two-Dose Vaccination with BNT162b2 in Health Care Workers. Clin. Microbiol. Infect. 2022, 28, 1024.e7–1024.e12. [Google Scholar] [CrossRef] [PubMed]
- Benning, L.; Klein, K.; Morath, C.; Bartenschlager, M.; Kim, H.; Buylaert, M.; Reineke, M.; Töllner, M.; Nusshag, C.; Kälble, F.; et al. Neutralizing Antibody Activity against the B.1.617.2 (Delta) Variant before and after a Third BNT162b2 Vaccine Dose in Hemodialysis Patients. Front. Immunol. 2022, 13, 840136. [Google Scholar] [CrossRef] [PubMed]
- Benning, L.; Morath, C.; Bartenschlager, M.; Kim, H.; Reineke, M.; Beimler, J.; Buylaert, M.; Nusshag, C.; Kälble, F.; Reichel, P.; et al. Neutralizing Antibody Response against the B.1.617.2 (Delta) and the B.1.1.529 (Omicron) Variants after a Third MRNA SARS-CoV-2 Vaccine Dose in Kidney Transplant Recipients. Am. J. Transplant. 2022, 22, 1873–1883. [Google Scholar] [CrossRef] [PubMed]
- Speer, C.; Benning, L.; Töllner, M.; Nusshag, C.; Kälble, F.; Reichel, P.; Schaier, M.; Bartenschlager, M.; Schnitzler, P.; Zeier, M.; et al. Neutralizing Antibody Response against Variants of Concern after Vaccination of Dialysis Patients with BNT162b2. Kidney Int. 2021, 100, 700–702. [Google Scholar] [CrossRef]
- Strangfeld, A.; Schäfer, M.; Gianfrancesco, M.A.; Lawson-Tovey, S.; Liew, J.W.; Ljung, L.; Mateus, E.F.; Richez, C.; Santos, M.J.; Schmajuk, G.; et al. Factors Associated with COVID-19-Related Death in People with Rheumatic Diseases: Results from the COVID-19 Global Rheumatology Alliance Physician-Reported Registry. Ann. Rheum. Dis. 2021, 80, 930–942. [Google Scholar] [CrossRef]
- Boekel, L.; Steenhuis, M.; Hooijberg, F.; Besten, Y.R.; van Kempen, Z.L.E.; Kummer, L.Y.; van Dam, K.P.J.; Stalman, E.W.; Vogelzang, E.H.; Cristianawati, O.; et al. Antibody Development after COVID-19 Vaccination in Patients with Autoimmune Diseases in the Netherlands: A Substudy of Data from Two Prospective Cohort Studies. Lancet Rheumatol. 2021, 3, e778–e788. [Google Scholar] [CrossRef]
- Jena, A.; Mishra, S.; Deepak, P.; Kumar-M., P.; Sharma, A.; Patel, Y.I.; Kennedy, N.A.; Kim, A.H.J.; Sharma, V.; Sebastian, S. Response to SARS-CoV-2 Vaccination in Immune Mediated Inflammatory Diseases: Systematic Review and Meta-Analysis. Autoimmun. Rev. 2022, 21, 102927. [Google Scholar] [CrossRef]
- Prendecki, M.; Clarke, C.; Edwards, H.; McIntyre, S.; Mortimer, P.; Gleeson, S.; Martin, P.; Thomson, T.; Randell, P.; Shah, A.; et al. Humoral and T-Cell Responses to SARS-CoV-2 Vaccination in Patients Receiving Immunosuppression. Ann. Rheum. Dis. 2021, 80, 1322–1329. [Google Scholar] [CrossRef]
- Marty, P.K.; Keulen, V.P.V.; Erskine, C.L.; Shah, M.; Hummel, A.; Stachowitz, M.; Fatis, S.; Granger, D.; Block, M.S.; Duarte-García, A.; et al. Antigen Specific Humoral and Cellular Immunity Following SARS-CoV-2 Vaccination in ANCA-Associated Vasculitis Patients Receiving B-Cell Depleting Therapy. Front. Immunol. 2022, 13, 834981. [Google Scholar] [CrossRef]
- Xie, Y.; Liu, Y.; Liu, Y. The Flare of Rheumatic Disease after SARS-CoV-2 Vaccination: A Review. Front. Immunol. 2022, 13, 919979. [Google Scholar] [CrossRef]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.-C.; Tiu, C.; Hu, Z.; Chen, V.C.-W.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 Surrogate Virus Neutralization Test Based on Antibody-Mediated Blockage of ACE2-Spike Protein-Protein Interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- Bray, R.A.; Lee, J.-H.; Brescia, P.; Kumar, D.; Nong, T.; Shih, R.; Woodle, E.S.; Maltzman, J.S.; Gebel, H.M. Development and Validation of a Multiplex, Bead-Based Assay to Detect Antibodies Directed Against SARS-CoV-2 Proteins. Transplantation 2021, 105, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Kronbichler, A.; Geetha, D.; Smith, R.M.; Egan, A.C.; Bajema, I.M.; Schönermarck, U.; Mahr, A.; Anders, H.-J.; Bruchfeld, A.; Cid, M.C.; et al. The COVID-19 Pandemic and ANCA-Associated Vasculitis—Reports from the EUVAS Meeting and EUVAS Education Forum. Autoimmun. Rev. 2021, 20, 102986. [Google Scholar] [CrossRef]
- David, R.; Hanna, P.; Lee, K.; Ritchie, A. Relapsed ANCA Associated Vasculitis Following Oxford AstraZeneca ChAdOx1-S COVID-19 Vaccination: A Case Series of Two Patients. Nephrology 2022, 27, 109–110. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, K.; Yamaguchi, S.; Yasuda, I.; Yoshimoto, N.; Kojima, D.; Kaneko, K.; Aso, M.; Nagasaka, T.; Yoshida, E.; Uchiyama, K.; et al. Development of Alveolar Hemorrhage after Pfizer-BioNTech COVID-19 MRNA Vaccination in a Patient with Renal-Limited Anti-Neutrophil Cytoplasmic Antibody-Associated Vasculitis: A Case Report. Front. Med. 2022, 9, 874831. [Google Scholar] [CrossRef]
- Fillon, A.; Sautenet, B.; Barbet, C.; Moret, L.; Thillard, E.M.; Jonville-Béra, A.P.; Halimi, J.M. De Novo and Relapsing Necrotizing Vasculitis after COVID-19 Vaccination. Clin. Kidney J. 2021, 15, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xiong, Y.; Xu, G. New Insights of Antineutrophil Cytoplasmic Antibody-Associated Vasculitis from the Perspective of COVID-19 Vaccination. Clin. Exp. Immunol. 2023; ahead of print. [Google Scholar] [CrossRef]
- Carruthers, J.E.; Wells, J.; Gupta, A.; Kallon, D.; Cox, A.; Pina, N.; Yaqoob, M.M.; Rajakariar, R. Response to Vaccination Against SARS-CoV-2 in Patients with Antineutrophil Cytoplasmic Antibody-Associated Vasculitis with Renal Involvement. Front. Med. 2022, 8, 817845. [Google Scholar] [CrossRef]
- Furer, V.; Eviatar, T.; Zisman, D.; Peleg, H.; Braun-Moscovici, Y.; Balbir-Gurman, A.; Paran, D.; Levartovsky, D.; Zisapel, M.; Elalouf, O.; et al. Predictors of Immunogenic Response to the BNT162b2 MRNA COVID-19 Vaccination in Patients with Autoimmune Inflammatory Rheumatic Diseases Treated with Rituximab. Vaccines 2022, 10, 901. [Google Scholar] [CrossRef]
- Chiang, T.P.-Y.; Connolly, C.M.; Ruddy, J.A.; Boyarsky, B.J.; Alejo, J.L.; Werbel, W.A.; Massie, A.; Christopher-Stine, L.; Garonzik-Wang, J.; Segev, D.L.; et al. Antibody Response to the Janssen/Johnson & Johnson SARS-CoV-2 Vaccine in Patients with Rheumatic and Musculoskeletal Diseases. Ann. Rheum. Dis. 2021, 80, 1365–1366. [Google Scholar] [CrossRef]
- Speer, C.; Töllner, M.; Benning, L.; Klein, K.; Bartenschlager, M.; Nusshag, C.; Kälble, F.; Reichel, P.; Schnitzler, P.; Zeier, M.; et al. Third COVID-19 Vaccine Dose with BNT162b2 in Patients with ANCA-Associated Vasculitis. Ann. Rheum. Dis. 2022, 81, 593–595. [Google Scholar] [CrossRef]
- Egri, N.; Calderón, H.; Martinez, R.; Vazquez, M.; Gómez-Caverzaschi, V.; Pascal, M.; Araújo, O.; Juan, M.; González-Navarro, E.A.; Hernández-Rodríguez, J. Cellular and Humoral Responses after Second and Third SARS-CoV-2 Vaccinations in Patients with Autoimmune Diseases Treated with Rituximab: Specific T Cell Immunity Remains Longer and Plays a Protective Role against SARS-CoV-2 Reinfections. Front. Immunol. 2023, 14, 1146841. [Google Scholar] [CrossRef] [PubMed]
- Kant, S.; Azar, A.; Geetha, D. Antibody Response to COVID-19 Booster Vaccine in Rituximab-Treated Patients with Anti–Neutrophil Cytoplasmic Antibody–Associated Vasculitis. Kidney Int. 2022, 101, 414–415. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, F.; Mrdenovic, N.; Scheicht, D.; Pons-Kühnemann, J.; Scheibelhut, C.; Strunk, J. Humoral Immunogenicity of COVID-19 Vaccines in Patients with Inflammatory Rheumatic Diseases under Treatment with Rituximab: A Case–Control Study (COVID-19VacRTX). Rheumatology 2022, 61, keac036. [Google Scholar] [CrossRef] [PubMed]
- Stefanski, A.; Rincon-Arevalo, H.; Schrezenmeier, E.; Karberg, K.; Szelinski, F.; Ritter, J.; Jahrsdörfer, B.; Schrezenmeier, H.; Ludwig, C.; Sattler, A.; et al. B Cell Numbers Predict Humoral and Cellular Response Upon SARS–CoV-2 Vaccination among Patients Treated with Rituximab. Arthritis Rheumatol. 2022, 74, 934–947. [Google Scholar] [CrossRef]
- Töllner, M.; Speer, C.; Benning, L.; Bartenschlager, M.; Nusshag, C.; Morath, C.; Zeier, M.; Süsal, C.; Schnitzler, P.; Schmitt, W.; et al. Impaired Neutralizing Antibody Activity against B.1.617.2 (Delta) after Anti-SARS-CoV-2 Vaccination in Patients Receiving Anti-CD20 Therapy. J. Clin. Med. 2022, 11, 1739. [Google Scholar] [CrossRef]
- Kant, S.; Kronbichler, A.; Salas, A.; Bruchfeld, A.; Geetha, D. Timing of COVID-19 Vaccine in the Setting of Anti-CD20 Therapy: A Primer for Nephrologists. Kidney Int. Rep. 2021, 6, 1197–1199. [Google Scholar] [CrossRef]
- Smith, J.B.; Gonzales, E.G.; Li, B.H.; Langer-Gould, A. Analysis of Rituximab Use, Time Between Rituximab and SARS-CoV-2 Vaccination, and COVID-19 Hospitalization or Death in Patients With Multiple Sclerosis. JAMA Netw. Open. 2022, 5, e2248664. [Google Scholar] [CrossRef]
- Troldborg, A.; Thomsen, M.K.; Bartels, L.E.; Andersen, J.B.; Vils, S.R.; Mistegaard, C.E.; Johannsen, A.D.; Hermansen, M.-L.F.; Mikkelsen, S.; Erikstrup, C.; et al. Time since Rituximab Treatment Is Essential for Developing a Humoral Response to COVID-19 MRNA Vaccines in Patients with Rheumatic Diseases. J. Rheumatol. 2022, 49, 644–649. [Google Scholar] [CrossRef]
- Khoury, D.S.; Schlub, T.E.; Cromer, D.; Steain, M.; Fong, Y.; Gilbert, P.B.; Subbarao, K.; Triccas, J.A.; Kent, S.J.; Davenport, M.P. Correlates of Protection, Thresholds of Protection, and Immunobridging among Persons with SARS-CoV-2 Infection. Emerg. Infect. Dis. 2023, 29, 381–388. [Google Scholar] [CrossRef]
- Piñana, J.L.; Martino, R.; Vazquez, L.; López-Corral, L.; Pérez, A.; Chorão, P.; Avendaño-Pita, A.; Pascual, M.-J.; Sánchez-Salinas, A.; Sanz-Linares, G.; et al. SARS-CoV-2-Reactive Antibody Waning, Booster Effect and Breakthrough SARS-CoV-2 Infection in Hematopoietic Stem Cell Transplant and Cell Therapy Recipients at One Year after Vaccination. Bone Marrow Transplant. 2023, 58, 567–580. [Google Scholar] [CrossRef]
- Sanghavi, D.K.; Bhakta, S.; Wadei, H.M.; Bosch, W.; Cowart, J.B.; Carter, R.E.; Shah, S.Z.; Pollock, B.D.; Neville, M.R.; Oman, S.P.; et al. Low Antispike Antibody Levels Correlate with Poor Outcomes in COVID-19 Breakthrough Hospitalizations. J. Intern. Med. 2022, 292, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Hadjadj, J.; Planas, D.; Ouedrani, A.; Buffier, S.; Delage, L.; Nguyen, Y.; Bruel, T.; Stolzenberg, M.-C.; Staropoli, I.; Ermak, N.; et al. Immunogenicity of BNT162b2 Vaccine against the Alpha and Delta Variants in Immunocompromised Patients with Systemic Inflammatory Diseases. Ann. Rheum. Dis. 2022, 81, 720–728. [Google Scholar] [CrossRef] [PubMed]
- Davis-Gardner, M.E.; Lai, L.; Wali, B.; Samaha, H.; Solis, D.; Lee, M.; Porter-Morrison, A.; Hentenaar, I.T.; Yamamoto, F.; Godbole, S.; et al. Neutralization against BA.2.75.2, BQ.1.1, and XBB from MRNA Bivalent Booster. N. Engl. J. Med. 2022, 388, 183–185. [Google Scholar] [CrossRef] [PubMed]
- Offit, P.A. Bivalent Covid-19 Vaccines—A Cautionary Tale. N. Engl. J. Med. 2023, 388, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Zhang, J.; Schiavon, C.R.; He, M.; Chen, L.; Shen, H.; Zhang, Y.; Yin, Q.; Cho, Y.; Andrade, L.; et al. SARS-CoV-2 Spike Protein Impairs Endothelial Function via Downregulation of ACE 2. Circ. Res. 2021, 128, 1323–1326. [Google Scholar] [CrossRef]
- Dolci, M.; Signorini, L.; D’Alessandro, S.; Perego, F.; Parapini, S.; Sommariva, M.; Taramelli, D.; Ferrante, P.; Basilico, N.; Delbue, S. In Vitro SARS-CoV-2 Infection of Microvascular Endothelial Cells: Effect on Pro-Inflammatory Cytokine and Chemokine Release. Int. J. Mol. Sci. 2022, 23, 4063. [Google Scholar] [CrossRef]
- Perico, L.; Benigni, A.; Remuzzi, G. SARS-CoV-2 and the Spike Protein in Endotheliopathy. Trends Microbiol. 2023; ahead of print. [Google Scholar] [CrossRef]
- Perico, L.; Morigi, M.; Galbusera, M.; Pezzotta, A.; Gastoldi, S.; Imberti, B.; Perna, A.; Ruggenenti, P.; Donadelli, R.; Benigni, A.; et al. SARS-CoV-2 Spike Protein 1 Activates Microvascular Endothelial Cells and Complement System Leading to Platelet Aggregation. Front. Immunol. 2022, 13, 827146. [Google Scholar] [CrossRef]
- Jarrot, P.-A.; Kaplanski, G. Pathogenesis of ANCA-Associated Vasculitis: An Update. Autoimmun. Rev. 2016, 15, 704–713. [Google Scholar] [CrossRef]
- Nakazawa, D.; Masuda, S.; Tomaru, U.; Ishizu, A. Pathogenesis and Therapeutic Interventions for ANCA-Associated Vasculitis. Nat. Rev. Rheumatol. 2019, 15, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Hakroush, S.; Tampe, B. Case Report: ANCA-Associated Vasculitis Presenting with Rhabdomyolysis and Pauci-Immune Crescentic Glomerulonephritis after Pfizer-BioNTech COVID-19 MRNA Vaccination. Front. Immunol. 2021, 12, 762006. [Google Scholar] [CrossRef] [PubMed]
- Simoncelli, E.; Conticini, E.; Colafrancesco, S.; Gattamelata, A.; Spinelli, F.R.; Garufi, C.; Truglia, S.; Grazzini, S.; Giardina, F.; Izzo, R.; et al. Multicentre Case-Control Study Evaluating the Safety of Anti-SARS-CoV-2 Vaccines in a Cohort of Patients with Systemic Vasculitis. Clin. Exp. Rheumatol. 2023, 41, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Krasselt, M.; Wagner, U.; Nguyen, P.; Pietsch, C.; Boldt, A.; Baerwald, C.; Pierer, M.; Seifert, O. Humoral and Cellular Response to COVID-19 Vaccination in Patients with Autoimmune Inflammatory Rheumatic Diseases under Real-Life Conditions. Rheumatology 2022, 61, SI180–SI188. [Google Scholar] [CrossRef] [PubMed]
- Röltgen, K.; Nielsen, S.C.A.; Silva, O.; Younes, S.F.; Zaslavsky, M.; Costales, C.; Yang, F.; Wirz, O.F.; Solis, D.; Hoh, R.A.; et al. Immune Imprinting, Breadth of Variant Recognition, and Germinal Center Response in Human SARS-CoV-2 Infection and Vaccination. Cell 2022, 185, 1025–1040.e14. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AAV Patients (N = 64) | Healthy Controls (N = 24) | p | |
|---|---|---|---|
| Age, median (IQR), y | 68 (55–72) | 61 (59–64) | 0.58 |
| Female sex, N (%) | 34 (53) | 14 (58) | 0.81 |
| Diagnosis, N (%) | |||
| Granulomatosis with polyangiitis | 47 (73) | - | - |
| Microscopic polyangiitis | 15 (23) | - | - |
| Eosinophile granulomatosis with polyangiitis | 2 (3) | - | - |
| Time since initial diagnosis, median (IQR), y | 8 (5–14) | - | - |
| ANCA ELISA, N (%) | |||
| PR3 | 44 (69) | - | - |
| MPO | 12 (19) | - | - |
| Double positive | 8 (13) | - | - |
| Organ involvement, N (%) * | |||
| Ears, nose, throat | 26 (43) | - | - |
| Kidney | 49 (82) | - | - |
| Lung | 45 (75) | - | - |
| Others | 25 (39) | - | - |
| Induction therapy, N (%) | |||
| CYC ± steroids | 38 (59) | - | - |
| Rituximab ± steroids | 26 (41) | - | - |
| Maintenance therapy, N (%) | |||
| ±steroids | 14 (22) | - | - |
| Azathioprine or MMF ± steroids | 22 (34) | - | - |
| Rituximab | 28 (44) | - | - |
| Time since last Rituximab dose, | 4 (2–7) | - | - |
| median (IQR), y | |||
| Vaccination regimen, N (%) | |||
| Homologous BNT162b2 | 49 (77) | 19 (79) | 0.99 |
| Homologous mRNA-1273 | 4 (6) | 0 (0) | 0.57 |
| Heterologous ChAdOx1/BNT162b2 | 6 (9) | 2 (8) | 0.99 |
| Heterologous ChAdOx1/mRNA-1273 | 2 (3) | 0 (0) | 0.99 |
| Homologous ChAdOx1 | 3 (5) | 3 (13) | 0.34 |
| AAV Patients (N = 21) | Healthy Controls (N = 24) | p | |
|---|---|---|---|
| Age, median (IQR), y | 71 (59–74) | 61 (59–64) | 0.16 |
| Female sex, N (%) | 7 (33) | 14 (58) | 0.14 |
| Diagnosis, N (%) | |||
| Granulomatosis with polyangiitis | 13 (62) | - | - |
| Microscopic polyangiitis | 8 (38) | - | - |
| Time since initial diagnosis, median (IQR), y | 4 (1-10) | - | - |
| ANCA ELISA, N (%) | |||
| PR3 | 11 (52) | - | - |
| MPO | 8 (38) | - | - |
| Double positive | 2 (10) | - | - |
| Organ involvement, N (%) | |||
| Ears, nose, throat | 4 (19) | - | - |
| Kidney | 18 (86) | - | - |
| Lung | 10 (48) | - | - |
| Others | 4 (19) | - | - |
| Induction therapy, N (%) | |||
| CYC ± steroids | 18 (86) | - | - |
| Rituximab ± steroids | 3 (14) | - | - |
| Maintenance therapy, N (%) | |||
| ±steroids | 4 (19) | - | - |
| Azathioprine or MMF ± steroids | 9 (43) | - | - |
| Rituximab | 8 (38) | - | - |
| Time since last Rituximab dose, | 3 (1–8) | - | - |
| median (IQR), y | |||
| Homologous mRNA vaccination, N (%) | 19 (90) | 19 (79) | 0.42 |
| Heterologous ChAdOx1/2x mRNA, N (%) | 2 (10) | 5 (21) | 0.42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Speer, C.; Töllner, M.; Benning, L.; Bartenschlager, M.; Kim, H.; Nusshag, C.; Kälble, F.; Reineke, M.; Reichel, P.; Schnitzler, P.; et al. BA.1/BA.5 Immunogenicity, Reactogenicity, and Disease Activity after COVID-19 Vaccination in Patients with ANCA-Associated Vasculitis: A Prospective Observational Cohort Study. Viruses 2023, 15, 1778. https://doi.org/10.3390/v15081778
Speer C, Töllner M, Benning L, Bartenschlager M, Kim H, Nusshag C, Kälble F, Reineke M, Reichel P, Schnitzler P, et al. BA.1/BA.5 Immunogenicity, Reactogenicity, and Disease Activity after COVID-19 Vaccination in Patients with ANCA-Associated Vasculitis: A Prospective Observational Cohort Study. Viruses. 2023; 15(8):1778. https://doi.org/10.3390/v15081778
Chicago/Turabian StyleSpeer, Claudius, Maximilian Töllner, Louise Benning, Marie Bartenschlager, Heeyoung Kim, Christian Nusshag, Florian Kälble, Marvin Reineke, Paula Reichel, Paul Schnitzler, and et al. 2023. "BA.1/BA.5 Immunogenicity, Reactogenicity, and Disease Activity after COVID-19 Vaccination in Patients with ANCA-Associated Vasculitis: A Prospective Observational Cohort Study" Viruses 15, no. 8: 1778. https://doi.org/10.3390/v15081778