
Seed Transmission of Wheat Streak Mosaic Virus and Triticum Mosaic Virus in Differentially Resistant Wheat Cultivars
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Virus Isolates, and Seed Collection
2.2. Phylogenetic Analysis
2.3. Detection of WSMV and TriMV in Seeds
2.4. Evaluation of WSMV and TriMV Transmission from Seeds to Progeny Plant
2.5. Data Analysis
3. Results
3.1. Virus Accumulation in the Field
3.2. Phylogenetic Analysis
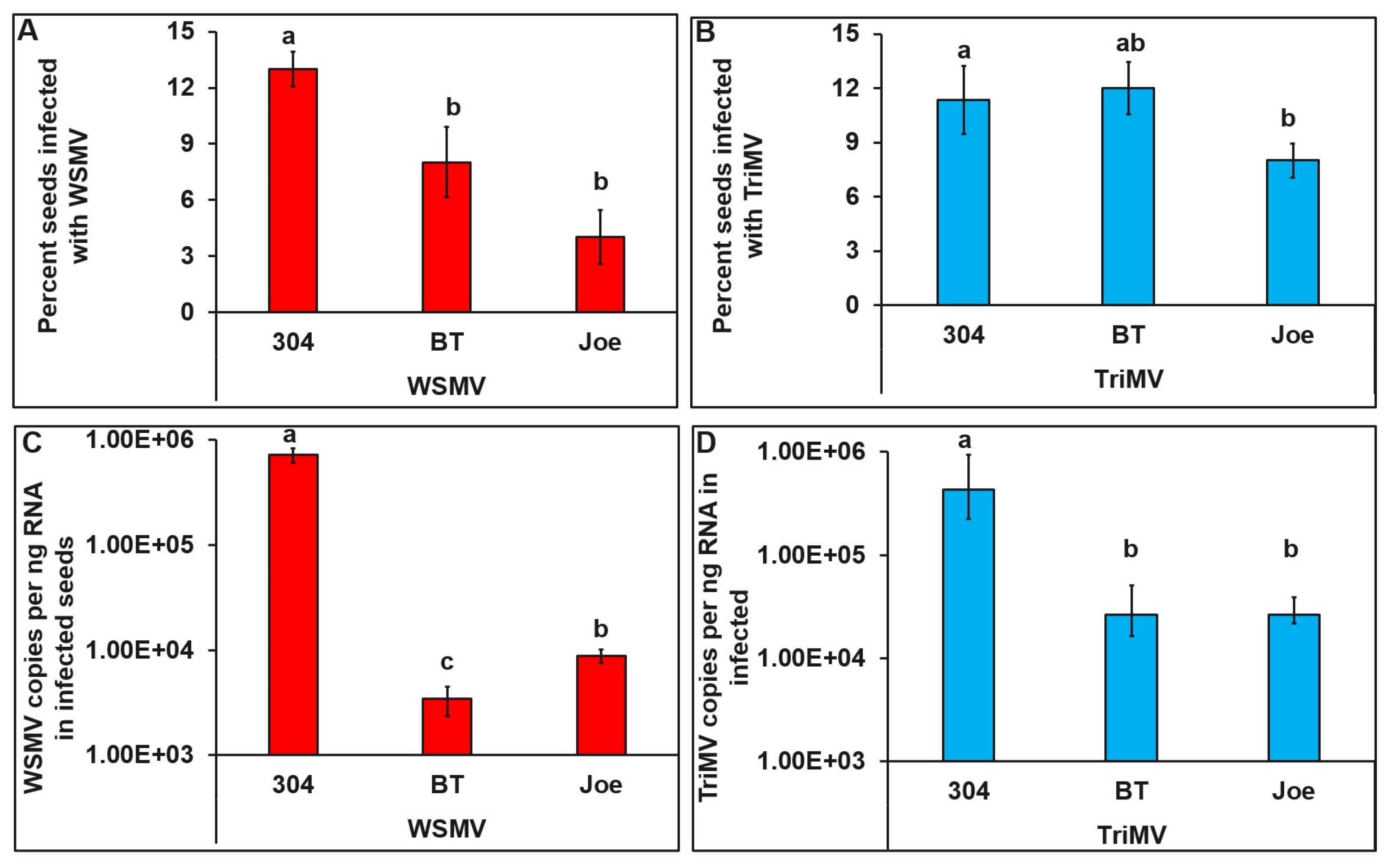
3.3. Detection of WSMV and TriMV in Field Collected Seeds
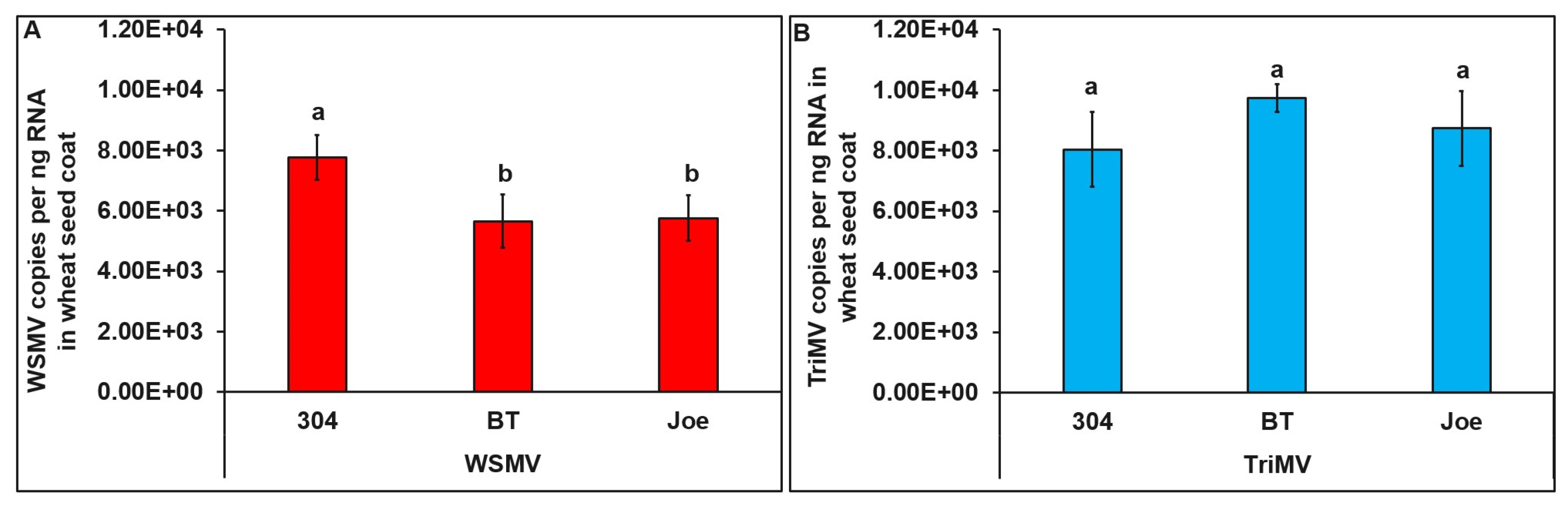
3.4. Distribution of WSMV and TriMV in Infected Seeds
3.5. Transmission of WSMV and TriMV from Seed to Progeny Wheat Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- U.S. Department of Agriculture National Agricultural Statistics Service. Quick Stats. 2017. Available online: https://quickstats.nass.usda.gov/ (accessed on 12 April 2023).
- Burrows, M.; Franc, G.; Rush, C.; Blunt, T.; Ito, D.; Kinzer, K.; Olson, J.; O’Mara, J.; Price, J.; Tande, C.; et al. Occurrence of Viruses in Wheat in the Great Plains Region, 2008. Plant Health Prog. 2009, 10, 14. [Google Scholar] [CrossRef]
- Fuentes-Bueno, I.; Price, J.A.; Rush, C.M.; Seifers, D.L.; Fellers, J.P. Triticum Mosaic Virus Isolates in the Southern Great Plains. Plant Dis. 2011, 95, 1516–1519. [Google Scholar] [CrossRef] [PubMed]
- Seifers, D.L.; Martin, T.J.; Harvey, T.L.; Fellers, J.P.; Michaud, J.P. Identification of the Wheat Curl Mite as the Vector of Triticum Mosaic Virus. Plant Dis. 2009, 93, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Slykhuis, J.T. Aceria Tulipae Keifer (Acarina: Eriophyidae) in Relation to the Spread of Wheat Streak Mosaic. Phytopathology 1955, 45, 116–128. [Google Scholar]
- Byamukama, E.; Wegulo, S.N.; Tatineni, S.; Hein, G.L.; Graybosch, R.A.; Baenziger, P.S.; French, R. Quantification of Yield Loss Caused by Triticum Mosaic Virus and Wheat Streak Mosaic Virus in Winter Wheat Under Field Conditions. Plant Dis. 2014, 98, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Byamukama, E.; Tatineni, S.; Hein, G.L.; Graybosch, R.A.; Baenziger, P.S.; French, R.; Wegulo, S.N. Effects of Single and Double Infections of Winter Wheat by Triticum Mosaic Virus and Wheat Streak Mosaic Virus on Yield Determinants. Plant Dis. 2012, 96, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Tatineni, S.; Graybosch, R.A.; Hein, G.L.; Wegulo, S.N.; French, R. Wheat Cultivar-Specific Disease Synergism and Alteration of Virus Accumulation During Co-Infection with Wheat Streak Mosaic Virus and Triticum Mosaic Virus. Phytopathology 2010, 100, 230–238. [Google Scholar] [CrossRef]
- Bryan, B.; Paetzold, L.; Workneh, F.; Rush, C.M. Incidence of Mite-Vectored Viruses of Wheat in the Texas High Plains and Interactions With Their Host and Vector. Plant Dis. 2019, 103, 2996–3001. [Google Scholar] [CrossRef]
- Stenger, D.C.; Hall, J.S.; Choi, I.-R.; French, R. Phylogenetic Relationships within the Family Potyviridae: Wheat Streak Mosaic Virus and Brome Streak Mosaic Virus Are Not Members of the Genus Rymovirus. Phytopathology 1998, 88, 782–787. [Google Scholar] [CrossRef]
- Fellers, J.P.; Seifers, D.; Ryba-White, M.; Joe Martin, T. The Complete Genome Sequence of Triticum Mosaic Virus, a New Wheat-Infecting Virus of the High Plains. Arch. Virol. 2009, 154, 1511–1515. [Google Scholar] [CrossRef]
- Tatineni, S.; Ziems, A.D.; Wegulo, S.N.; French, R. Triticum Mosaic Virus: A Distinct Member of the Family Potyviridae with an Unusually Long Leader Sequence. Phytopathology 2009, 99, 943–950. [Google Scholar] [CrossRef] [PubMed]
- McKinney, H.H. Mosaic Diseases of Wheat and Related Cereals; US Department of Agriculture Circular: Washington DC, USA, 1937; pp. 1–23.
- Seifers, D.L.; Martin, T.J.; Harvey, T.L.; Fellers, J.P.; Stack, J.P.; Ryba-White, M.; Haber, S.; Krokhin, O.; Spicer, V.; Lovat, N.; et al. Triticum Mosaic Virus: A New Virus Isolated from Wheat in Kansas. Plant Dis. 2008, 92, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Bockus, W.; Bowden, R.; Hunger, R.; Morrill, W.; Murray, T.; Smiley, R. (Eds.) Gary Hein Wheat Curl Mite. In Compendium of Wheat Diseases and Pests; American Phytopathological Society: St Paul, MN, USA, 2010; pp. 132–133. [Google Scholar]
- Graybosch, R.A.; Peterson, C.J.; Baenziger, P.S.; Baltensperger, D.D.; Nelson, L.A.; Jin, Y.; Kolmer, J.; Seabourn, B.; French, R.; Hein, G.; et al. Registration of ‘Mace’ Hard Red Winter Wheat. J. Plant Regist. 2009, 3, 51–56. [Google Scholar] [CrossRef]
- Lu, H.; Price, J.; Devkota, R.; Rush, C.; Rudd, J. A Dominant Gene for Resistance to Wheat Streak Mosaic Virus in Winter Wheat Line CO960293-2. Crop Sci. 2011, 51, 5–12. [Google Scholar] [CrossRef]
- Haley, S.D.; Martin, T.J.; Quick, J.S.; Seifers, D.L.; Stromberger, J.A.; Clayshulte, S.R.; Clifford, B.L.; Peairs, F.B.; Rudolph, J.B.; Johnson, J.J.; et al. Registration of CO960293-2 Wheat Germplasm Resistant to Wheat Streak Mosaic Virus and Russian Wheat Aphid. Crop Sci. 2002, 42, 1381–1382. [Google Scholar] [CrossRef]
- Haley, S.D.; Johnson, J.J.; Peairs, F.B.; Stromberger, J.A.; Heaton, E.E.; Seifert, S.A.; Kottke, R.A.; Rudolph, J.B.; Martin, T.J.; Bai, G.; et al. Registration of ‘Snowmass’ Wheat. J. Plant Regist. 2011, 5, 87–90. [Google Scholar] [CrossRef]
- Seifers, D.L.; Martin, T.J.; Harvey, T.L.; Haber, S. Temperature-Sensitive Wheat Streak Mosaic Virus Resistance Identified in KS03HW12 Wheat. Plant Dis. 2007, 91, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Tatineni, S.; Wosula, E.N.; Bartels, M.; Hein, G.L.; Graybosch, R.A. Temperature-Dependent Wsm1 and Wsm2 Gene-Specific Blockage of Viral Long-Distance Transport Provides Resistance to Wheat Streak Mosaic Virus and Triticum Mosaic Virus in Wheat. Mol. Plant-Microbe Interact. 2016, 29, 724–738. [Google Scholar] [CrossRef]
- Seifers, D.L. Temperature Sensitivity and Efficacy of Wheat Streak Mosaic Virus Resistance Derived from Agropyron Intermedium. Plant Dis. 1995, 79, 1104. [Google Scholar] [CrossRef]
- Triebe, B.; Mukai, Y.; Dhaliwal, H.S.; Martin, T.J.; Gill, B.S. Identification of Alien Chromatin Specifying Resistance to Wheat Streak Mosaic and Greenbug in Wheat Germ Plasm by C-Banding and in Situ Hybridization. Theor. Appl. Genet. 1991, 81, 381–389. [Google Scholar] [CrossRef]
- Liu, W.; Seifers, D.L.; Qi, L.L.; Friebe, B.; Gill, B.S. A Compensating Wheat-Thinopyrum Intermedium Robertsonian Translocation Conferring Resistance to Wheat Streak Mosaic Virus and Triticum Mosaic Virus. Crop Sci. 2011, 51, 2382–2390. [Google Scholar] [CrossRef]
- Sastry, K.S. Seed-Borne Plant Virus Diseases; Springer: Delhi, India, 2013; ISBN 978-81-322-0812-9. [Google Scholar]
- Jones, R.A.C.; Coutts, B.A.; Mackie, A.E.; Dwyer, G.I. Seed Transmission of Wheat Streak Mosaic Virus Shown Unequivocally in Wheat. Plant Dis. 2005, 89, 1048–1050. [Google Scholar] [CrossRef] [PubMed]
- Coutts, B.A.; Banovic, M.; Kehoe, M.A.; Severtson, D.L.; Jones, R.A.C. Epidemiology of Wheat Streak Mosaic Virus in Wheat in a Mediterranean-Type Environment. Eur. J. Plant Pathol. 2014, 140, 797–813. [Google Scholar] [CrossRef]
- Lanoiselet, V.M.; Hind-Lanoiselet, T.L.; Murray, G.M. Studies on the Seed Transmission of Wheat Streak Mosaic Virus. Australas. Plant Pathol. 2008, 37, 584. [Google Scholar] [CrossRef]
- Drummond, A.J.; Ashton, B.; Buxton, S.; Cheung, M.; Cooper, A.; Duran, C.; Field, M.; Heled, J.; Kearse, M.; Markowitz, S.; et al. Geneious, version Pro v. 8.1.9, Biomatters: Auckland, New Zealand, 2014.
- Schliep, K.P. Phangorn: Phylogenetic Analysis in R. Bioinformatics 2011, 27, 592–593. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v4: Recent Updates and New Developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 2nd ed.; Sage Publications: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- Lenth, R.V.; Bolker, B.; Buerkner, P.; Giné-Vázquez, I.; Herve, M.; Jung, M.; Love, J.; Miguez, F.; Riebl, H.; Singmann, H. Estimated Marginal Means, Aka Least-Squares Means [R Package Emmeans Version 1.7.0]. Am. Stat. 2023, 34, 216–222. [Google Scholar]
- Sowell, A. Bryn Swearingen Wheat Sector at a Glance; USDA ERS: Washington DC, USA, 2022. [Google Scholar]
- Pagán, I. Transmission through Seeds: The Unknown Life of Plant Viruses. PLoS Pathog. 2022, 18, e1010707. [Google Scholar] [CrossRef]
- Oliveira-Hofman, C.; Wegulo, S.N.; Tatineni, S.; Hein, G.L. Impact of Wheat Streak Mosaic Virus and Triticum Mosaic Virus Coinfection of Wheat on Transmission Rates by Wheat Curl Mites. Plant Dis. 2015, 99, 1170–1174. [Google Scholar] [CrossRef]
- Torre, C.; Agüero, J.; Gómez-Aix, C.; Aranda, M.A. Comparison of DAS-ELISA and QRT-PCR for the Detection of Cucurbit Viruses in Seeds. Ann. Appl. Biol. 2020, 176, 158–169. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gautam, S.; Chinnaiah, S.; Herron, B.; Workneh, F.; Rush, C.M.; Gadhave, K.R. Seed Transmission of Wheat Streak Mosaic Virus and Triticum Mosaic Virus in Differentially Resistant Wheat Cultivars. Viruses 2023, 15, 1774. https://doi.org/10.3390/v15081774
Gautam S, Chinnaiah S, Herron B, Workneh F, Rush CM, Gadhave KR. Seed Transmission of Wheat Streak Mosaic Virus and Triticum Mosaic Virus in Differentially Resistant Wheat Cultivars. Viruses. 2023; 15(8):1774. https://doi.org/10.3390/v15081774
Chicago/Turabian StyleGautam, Saurabh, Senthilraja Chinnaiah, Benjamin Herron, Fekede Workneh, Charles M. Rush, and Kiran R. Gadhave. 2023. "Seed Transmission of Wheat Streak Mosaic Virus and Triticum Mosaic Virus in Differentially Resistant Wheat Cultivars" Viruses 15, no. 8: 1774. https://doi.org/10.3390/v15081774