
The 2022 Outbreaks of African Swine Fever Virus Demonstrate the First Report of Genotype II in Ghana
 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Next Generation Sequencing
2.2. Genome Assembly
2.3. Annotation of the Genome
2.4. Identification of Single-Nucleotide Polymorphisms (SNPs)
2.5. Genome Alignment
2.6. Protein Alignment
3. Results
3.1. Characteristics of Collected 2022 Outbreak Samples
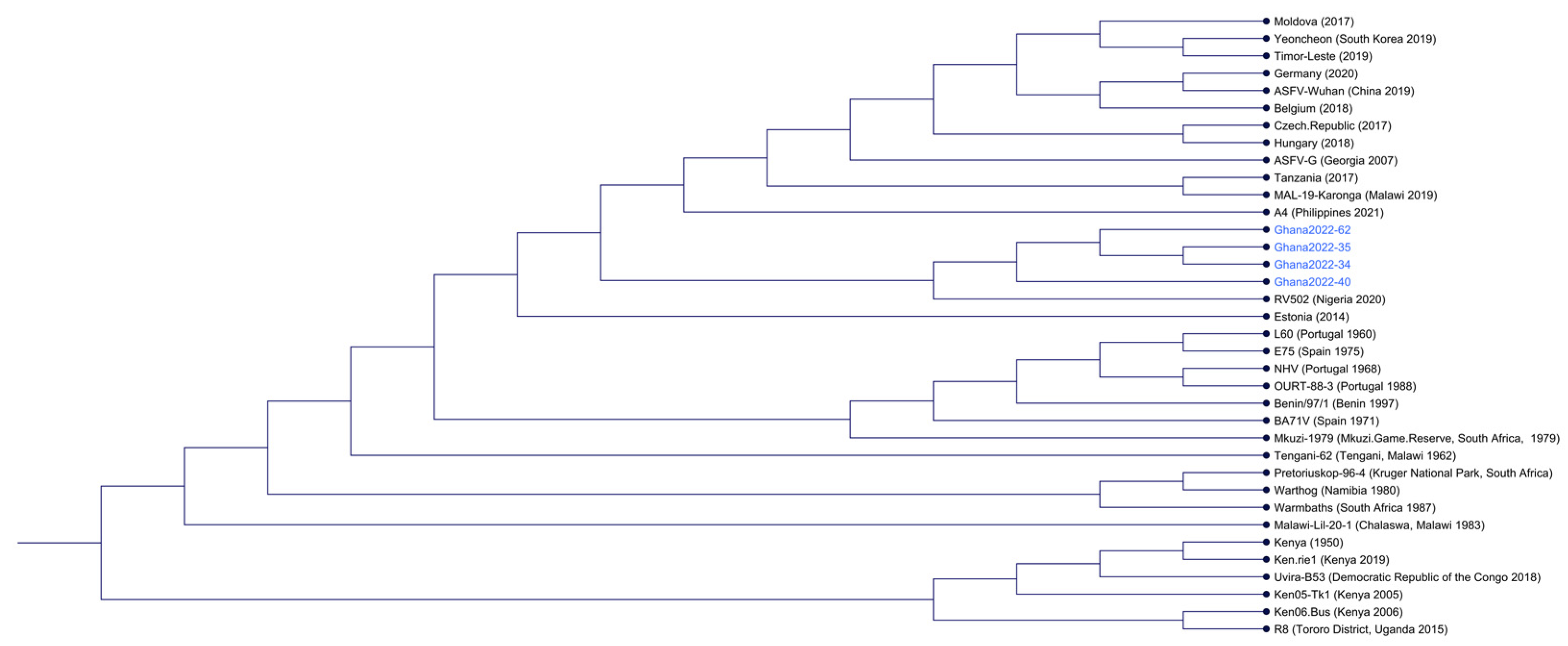
3.2. Genotyping of the ASFV Isolates
3.3. ASFV Full-Genome Alignments
3.4. Genetic Variation between Ghana 2022 Isolates
3.5. Analysis of a 6534 nt Deletion and Individual MGFs
3.6. Analysis of Ghana 2022 Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Montgomery, R.E. On a Form of Swine Fever Occurring in British East Africa (Kenya Colony). J. Comp. Pathol. Ther. 1921, 34, 159–191. [Google Scholar] [CrossRef]
- Chapman, D.A.; Darby, A.C.; Da Silva, M.; Upton, C.; Radford, A.D.; Dixon, L.K. Genomic analysis of highly virulent Georgia 2007/1 isolate of African swine fever virus. Emerg. Infect. Dis. 2011, 17, 599–605. [Google Scholar] [CrossRef]
- Ge, S.; Li, J.; Fan, X.; Liu, F.; Li, L.; Wang, Q.; Ren, W.; Bao, J.; Liu, C.; Wang, H.; et al. Molecular Characterization of African Swine Fever Virus, China, 2018. Emerg. Infect. Dis. 2018, 24, 2131–2133. [Google Scholar] [CrossRef]
- Ramirez-Medina, E.; O’Donnell, V.; Silva, E.; Espinoza, N.; Velazquez-Salinas, L.; Moran, K.; Daite, D.A.; Barrette, R.; Faburay, B.; Holland, R.; et al. Experimental Infection of Domestic Pigs with an African Swine Fever Virus Field Strain Isolated in 2021 from the Dominican Republic. Viruses 2022, 14, 1090. [Google Scholar] [CrossRef]
- Spinard, E.; O’Donnell, V.; Vuono, E.; Rai, A.; Davis, C.; Ramirez-Medina, E.; Espinoza, N.; Valladares, A.; Borca, M.V.; Gladue, D.P. Full genome sequence for the African swine fever virus outbreak in the Dominican Republic in 1980. Sci. Rep. 2023, 13, 1024. [Google Scholar] [CrossRef]
- Chen, W.Y.; Zhao, D.M.; He, X.J.; Liu, R.Q.; Wang, Z.L.; Zhang, X.F.; Li, F.; Shan, D.; Chen, H.F.; Zhang, J.W.; et al. A seven-gene-deleted African swine fever virus is safe and effective as a live attenuated vaccine in pigs. Sci. China Life Sci. 2020, 63, 623–634. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, L.; Zhang, H.; Li, Y.; Ge, X.; Guo, X.; Yu, K.; Yang, H. Interactome profile of the host cellular proteins and the nonstructural protein 2 of porcine reproductive and respiratory syndrome virus. PLoS ONE 2014, 9, e99176. [Google Scholar] [CrossRef]
- Liu, R.; Sun, Y.; Chai, Y.; Li, S.; Li, S.; Wang, L.; Su, J.; Yu, S.; Yan, J.; Gao, F.; et al. The structural basis of African swine fever virus pA104R binding to DNA and its inhibition by stilbene derivatives. Proc. Natl. Acad. Sci. USA 2020, 117, 11000–11009. [Google Scholar] [CrossRef]
- Spinard, E.; Azzinaro, P.; Rai, A.; Espinoza, N.; Ramirez-Medina, E.; Valladares, A.; Borca, M.V.; Gladue, D.P. Complete Structural Predictions of the Proteome of African Swine Fever Virus Strain Georgia 2007. Microbiol. Resour. Announc. 2022, 11, e0088122. [Google Scholar] [CrossRef]
- Quembo, C.J.; Jori, F.; Vosloo, W.; Heath, L. Genetic characterization of African swine fever virus isolates from soft ticks at the wildlife/domestic interface in Mozambique and identification of a novel genotype. Transbound. Emerg. Dis. 2018, 65, 420–431. [Google Scholar] [CrossRef]
- Achenbach, J.E.; Gallardo, C.; Nieto-Pelegrín, E.; Rivera-Arroyo, B.; Degefa-Negi, T.; Arias, M.; Jenberie, S.; Mulisa, D.D.; Gizaw, D.; Gelaye, E.; et al. Identification of a New Genotype of African Swine Fever Virus in Domestic Pigs from Ethiopia. Transbound. Emerg. Dis. 2017, 64, 1393–1404. [Google Scholar] [CrossRef]
- Dixon, L.K.; Chapman, D.A.; Netherton, C.L.; Upton, C. African swine fever virus replication and genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef]
- Bastos, A.D.S.; Penrith, M.L.; Cruciere, C.; Edrich, J.L.; Hutchings, G.; Roger, F.; Couacy-Hymann, E.; Thomson, G.R. Genotyping field strains of African swine fever virus by partial p72 gene characterisation. Arch. Virol. 2003, 148, 693–706. [Google Scholar] [CrossRef]
- Qu, H.; Ge, S.; Zhang, Y.; Wu, X.; Wang, Z. A systematic review of genotypes and serogroups of African swine fever virus. Virus Genes 2022, 58, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Gladue, D.P.; Borca, M.V. Recombinant ASF Live Attenuated Virus Strains as Experimental Vaccine Candidates. Viruses 2022, 14, 878. [Google Scholar] [CrossRef] [PubMed]
- Odoom, T. African Swine Fever updates from Ghana. In Proceedings of the Regional Workshop for the International Networking of African Swine Fever Research in Asia, Tokyo, Japan, 26–27 February 2020. [Google Scholar]
- Borca, M.V.; Berggren, K.A.; Ramirez-Medina, E.; Vuono, E.A.; Gladue, D.P. CRISPR/Cas Gene Editing of a Large DNA Virus: African Swine Fever Virus. Bio-Protocol 2018, 8, e2978. [Google Scholar] [CrossRef]
- Borca, M.V.; Holinka, L.G.; Berggren, K.A.; Gladue, D.P. CRISPR-Cas9, a tool to efficiently increase the development of recombinant African swine fever viruses. Sci. Rep. 2018, 8, 3154. [Google Scholar] [CrossRef]
- O’Donnell, V.K.; Grau, F.R.; Mayr, G.A.; Sturgill Samayoa, T.L.; Dodd, K.A.; Barrette, R.W. Rapid Sequence-Based Characterization of African Swine Fever Virus by Use of the Oxford Nanopore MinION Sequence Sensing Device and a Companion Analysis Software Tool. J. Clin. Microbiol. 2019, 58, e01104-19. [Google Scholar] [CrossRef] [PubMed]
- Tcherepanov, V.; Ehlers, A.; Upton, C. Genome Annotation Transfer Utility (GATU): Rapid annotation of viral genomes using a closely related reference genome. BMC Genom. 2006, 7, 150. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ambagala, A.; Goonewardene, K.; Lamboo, L.; Goolia, M.; Erdelyan, C.; Fisher, M.; Handel, K.; Lung, O.L.V.; Blome, S.; King, J.; et al. Characterization of a Novel African Swine Fever Virus p72 Genotype II from Nigeria. Viruses 2023, 15, 915. [Google Scholar] [CrossRef] [PubMed]
- Zani, L.; Forth, J.H.; Forth, L.; Nurmoja, I.; Leidenberger, S.; Henke, J.; Carlson, J.; Breidenstein, C.; Viltrop, A.; Hoper, D.; et al. Deletion at the 5′-end of Estonian ASFV strains associated with an attenuated phenotype. Sci. Rep. 2018, 8, 6510. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Q.H.; Zhu, Z.Y.; Wang, S.J.; Tu, S.Y.; Zhang, Y.Q.; Zou, Y.L.; Liu, Y.T.; Liu, C.J.; Ren, W.J.; et al. Tracing the Origin of Genotype II African Swine Fever Virus in China by Genomic Epidemiology Analysis. Transbound. Emerg. Dis. 2023, 2023, 4820809. [Google Scholar] [CrossRef]
- O’Donnell, V.; Holinka, L.G.; Gladue, D.P.; Sanford, B.; Krug, P.W.; Lu, X.Q.; Arzt, J.; Reese, B.; Carrillo, C.; Risatti, G.R.; et al. African Swine Fever Virus Georgia Isolate Harboring Deletions of MGF360 and MGF505 Genes Is Attenuated in Swine and Confers Protection against Challenge with Virulent Parental Virus. J. Virol. 2015, 89, 6048–6056. [Google Scholar] [CrossRef]
- Ramirez-Medina, E.; Vuono, E.; Pruitt, S.; Rai, A.; Silva, E.; Espinoza, N.; Zhu, J.; Velazquez-Salinas, L.; Borca, M.V.; Gladue, D.P. Development and In Vivo Evaluation of a MGF110-1L Deletion Mutant in African Swine Fever Strain Georgia. Viruses 2021, 13, 286. [Google Scholar] [CrossRef]
- Borca, M.V.; O’Donnell, V.; Holinka, L.G.; Sanford, B.; Azzinaro, P.A.; Risatti, G.R.; Gladue, D.P. Development of a fluorescent ASFV strain that retains the ability to cause disease in swine. Sci. Rep. 2017, 7, srep46747. [Google Scholar] [CrossRef]
- Ramirez-Medina, E.; Vuono, E.A.; Rai, A.; Pruitt, S.; Silva, E.; Velazquez-Salinas, L.; Zhu, J.; Gladue, D.P.; Borca, M.V. Evaluation in Swine of a Recombinant African Swine Fever Virus Lacking the MGF-360-1L Gene. Viruses 2020, 12, 1193. [Google Scholar] [CrossRef]
- Sanchez-Cordon, P.J.; Jabbar, T.; Berrezaie, M.; Chapman, D.; Reis, A.; Sastre, P.; Rueda, P.; Goatley, L.; Dixon, L.K. Evaluation of protection induced by immunisation of domestic pigs with deletion mutant African swine fever virus Benin Delta MGF by different doses and routes. Vaccine 2018, 36, 707–715. [Google Scholar] [CrossRef]
- Ramirez-Medina, E.; Vuono, E.A.; Velazquez-Salinas, L.; Silva, E.; Rai, A.; Pruitt, S.; Berggren, K.A.; Zhu, J.; Borca, M.V.; Gladue, D.P. The MGF360-16R ORF of African Swine Fever Virus Strain Georgia Encodes for a Nonessential Gene That Interacts with Host Proteins SERTAD3 and SDCBP. Viruses 2020, 12, 60. [Google Scholar] [CrossRef]
- Leitao, A.; Cartaxeiro, C.; Coelho, R.; Cruz, B.; Parkhouse, R.M.E.; Portugal, F.C.; Vigario, J.D.; Martins, C.L.V. The non-haemadsorbing African swine fever virus isolate ASFV/NH/P68 provides a model for defining the protective anti-virus immune response. J. Gen. Virol. 2001, 82, 513–523. [Google Scholar] [CrossRef]
- Rathakrishnan, A.; Connell, S.; Petrovan, V.; Moffat, K.; Goatley, L.C.; Jabbar, T.; Sánchez-Cordón, P.J.; Reis, A.L.; Dixon, L.K. Differential Effect of Deleting Members of African Swine Fever Virus Multigene Families 360 and 505 from the Genotype II Georgia 2007/1 Isolate on Virus Replication, Virulence, and Induction of Protection. J. Virol. 2022, 96, e01899-21. [Google Scholar] [CrossRef]
- Burrage, T.G.; Lu, Z.; Neilan, J.G.; Rock, D.L.; Zsak, L. African swine fever virus multigene family 360 genes affect virus replication and generalization of infection in Ornithodoros porcinus ticks. J. Virol. 2004, 78, 2445–2453. [Google Scholar] [CrossRef]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Gay, C.G.; Gladue, D.P. ASFV-G-∆I177L as an Effective Oral Nasal Vaccine against the Eurasia Strain of Africa Swine Fever. Viruses 2021, 13, 765. [Google Scholar] [CrossRef]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Holinka, L.G.; Velazquez-Salinas, L.; Zhu, J.; Gladue, D.P. Development of a Highly Effective African Swine Fever Virus Vaccine by Deletion of the I177L Gene Results in Sterile Immunity against the Current Epidemic Eurasia Strain. J. Virol. 2020, 94, e02017-19. [Google Scholar] [CrossRef]
- O’Donnell, V.; Risatti, G.R.; Holinka, L.G.; Krug, P.W.; Carlson, J.; Velazquez-Salinas, L.; Azzinaro, P.A.; Gladue, D.P.; Borca, M.V. Simultaneous Deletion of the 9GL and UK Genes from the African Swine Fever Virus Georgia 2007 Isolate Offers Increased Safety and Protection against Homologous Challenge. J. Virol. 2017, 91, e01760-16. [Google Scholar] [CrossRef] [PubMed]
- Gladue, D.P.; Ramirez-Medina, E.; Vuono, E.; Silva, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Borca, M.V. Deletion of the A137R Gene from the Pandemic Strain of African Swine Fever Virus Attenuates the Strain and Offers Protection against the Virulent Pandemic Virus. J. Virol. 2021, 95, e0113921. [Google Scholar] [CrossRef]
- Tran, X.H.; Le, T.T.P.; Nguyen, Q.H.; Do, T.T.; Nguyen, V.D.; Gay, C.G.; Borca, M.V.; Gladue, D.P. African swine fever virus vaccine candidate ASFV-G-DeltaI177L efficiently protects European and native pig breeds against circulating Vietnamese field strain. Transbound. Emerg. Dis. 2022, 69, e497–e504. [Google Scholar] [CrossRef] [PubMed]
- Osei-Amponsah, R.; Skinner, B.M.; Adjei, D.O.; Bauer, J.; Larson, G.; Affara, N.A.; Sargent, C.A. Origin and phylogenetic status of the local Ashanti Dwarf pig (ADP) of Ghana based on genetic analysis. BMC Genom. 2017, 18, 193. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sample | GenBank Accession | Location of Animal/Farm | Region | Date of Outbreak | Genotype p72/VP73 (B646L) | Genotype p54 (E183L) |
|---|---|---|---|---|---|---|
| 35 | OP479889 | AKUMADAN | Ashanti | 1/4/2022 | II | IIa |
| 62 | OP718535 | CENTRAL GONJA DISTRICT | Northern | 2/3/2022 | II | IIa |
| 34 | OP718533 | BUNSO | Eastern | 3/7/2022 | II | IIa |
| 40 | OP718534 | SUHYEN | Eastern | 3/11/2022 | II | IIa |
| Gene | Ghana2022-34 | Ghana2022-35 | Ghana2022-40 | Ghana2022-62 |
|---|---|---|---|---|
| B117L | Y98F | Y98F | Y98F | Y98F |
| B602L | V13I | V13I | V13I | V13I |
| C315R | no mutation | no mutation | Q30H | no mutation |
| C475L | no mutation | no mutation | no mutation | Q148H |
| EP402R | na | L300V, P313L | na | na |
| EP424R | N155S | N155S | N155S | N155S |
| F334L | S282G | S282G | S282G | S282G |
| H339R | Q319R | Q319R | Q319R | Q319R |
| K145R | no mutation | no mutation | Y116R | no mutation |
| KP360L | F19L, H41Y, K47E | F19L, H41Y, K47E | F19L, H41Y, K47E | F19L, H41Y, K47E |
| MGF_110-1L | 54 AA extension to the C-terminus | 54 AA extension to the C-terminus | 54 AA extension to the C-terminus | 54 AA extension to the C-terminus |
| MGF_110-7L | Y112R | Y112R | Y112R | Y112R |
| MGF_360-13L | G177D, NINQAMLTSVQYYNIGNIFFCID 262-284 ATSTKLCLLQYNIITSVIYFSV* | G177D, NINQAMLTSVQYYNIGNIFFCID 262-284 ATSTKLCLLQYNIITSVIYFSV* | G177D, NINQAMLTSVQYYN 262-276 QHQPSYAYFSTIL* | G177D, NINQAMLTSVQYYN 262-276 QHQPSYAYFSTIL* |
| MGF_360-14L | P351L | P351L | P351L | P351L |
| MGF_360-16R | D92*, split into MGF_360-16Ra and MGF_360-16Rb | D92*, split into MGF_360-16Ra and MGF_360-16Rb | D92*, split into MGF_360-16Ra and MGF_360-16Rb | D92*, split into MGF_360-16Ra and MGF_360-16Rb |
| MGF_360-18R | no mutation | no mutation | no mutation | Split into MGF 360-18Ra and MGF 360-18Rb |
| MGF_360-6L | LHKKILEPSE 341-350 RFTTNPLSS* | LHKKILEPSE 341-350 RFTTNPLSS* | na | LHKKILEPSE 341-350 RFTTNPLSS* |
| MGF_505-11L | S231L | S231L | S231L, I280V | S231L, I280V |
| MGF_505-1R | I450R | I450R | I450R | I450R |
| MGF_505-2R | T355A | T355A | T355A | T355A |
| MGF_505-5R | G477S | G477S | G477S | G477S |
| NP868R | A9T, A589V | A9T, A589V | A9T, A589V | A9T, A589V |
| X69R | L16P | L16P | no mutation | no mutation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spinard, E.; Rai, A.; Osei-Bonsu, J.; O’Donnell, V.; Ababio, P.T.; Tawiah-Yingar, D.; Arthur, D.; Baah, D.; Ramirez-Medina, E.; Espinoza, N.; et al. The 2022 Outbreaks of African Swine Fever Virus Demonstrate the First Report of Genotype II in Ghana. Viruses 2023, 15, 1722. https://doi.org/10.3390/v15081722
Spinard E, Rai A, Osei-Bonsu J, O’Donnell V, Ababio PT, Tawiah-Yingar D, Arthur D, Baah D, Ramirez-Medina E, Espinoza N, et al. The 2022 Outbreaks of African Swine Fever Virus Demonstrate the First Report of Genotype II in Ghana. Viruses. 2023; 15(8):1722. https://doi.org/10.3390/v15081722
Chicago/Turabian StyleSpinard, Edward, Ayushi Rai, Jehadi Osei-Bonsu, Vivian O’Donnell, Patrick T. Ababio, Daniel Tawiah-Yingar, Daniel Arthur, Daniel Baah, Elizabeth Ramirez-Medina, Nallely Espinoza, and et al. 2023. "The 2022 Outbreaks of African Swine Fever Virus Demonstrate the First Report of Genotype II in Ghana" Viruses 15, no. 8: 1722. https://doi.org/10.3390/v15081722