
First Report of Alphacoronavirus Circulating in Cavernicolous Bats from Portugal
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bokelmann, M.; Balkema-buschmann, A. Coronaviruses in Bats. Berl. Munch. Tierarztl. Wochenschr. 2021, 134, 1–16. [Google Scholar] [CrossRef]
- Domańska-Blicharz, K.; Miłek-Krupa, J.; Pikuła, A. Diversity of Coronaviruses in Wild Representatives of the Aves Class in Poland. Viruses 2021, 13, 1497. [Google Scholar] [CrossRef] [PubMed]
- Balboni, A.; Palladini, A.; Bogliani, G.; Battilani, M. Detection of a Virus Related to Betacoronaviruses in Italian Greater Horseshoe Bats. Epidemiol. Infect. 2011, 139, 216–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, A.; Kulcsar, K.; Misra, V.; Frieman, M.; Mossman, K. Bats and Coronaviruses. Viruses 2019, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Gloza-Rausch, F.; Ipsen, A.; Seebens, A.; Göttsche, M.; Panning, M.; Drexler, J.F.; Petersen, N.; Annan, A.; Grywna, K.; Müller, M.; et al. Detection and Prevalence Patterns of Group I Coronaviruses in Bats, Northern Germany. Emerg. Infect. Dis. 2008, 14, 626–631. [Google Scholar] [CrossRef]
- Mihindukulasuriya, K.A.; Wu, G.; St. Leger, J.; Nordhausen, R.W.; Wang, D. Identification of a Novel Coronavirus from a Beluga Whale by Using a Panviral Microarray. J. Virol. 2008, 82, 5084–5088. [Google Scholar] [CrossRef] [Green Version]
- Woo, P.C.Y.; Lau, S.K.P.; Lam, C.S.F.; Lau, C.C.Y.; Tsang, A.K.L.; Lau, J.H.N.; Bai, R.; Teng, J.L.L.; Tsang, C.C.C.; Wang, M.; et al. Discovery of Seven Novel Mammalian and Avian Coronaviruses in the Genus Deltacoronavirus Supports Bat Coronaviruses as the Gene Source of Alphacoronavirus and Betacoronavirus and Avian Coronaviruses as the Gene Source of Gammacoronavirus and Deltacoronavi. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [CrossRef] [Green Version]
- Woo, P.C.Y.; Lau, S.K.P.; Lam, C.S.F.; Tsang, A.K.L.; Hui, S.-W.; Fan, R.Y.Y.; Martelli, P.; Yuen, K.-Y. Discovery of a Novel Bottlenose Dolphin Coronavirus Reveals a Distinct Species of Marine Mammal Coronavirus in Gammacoronavirus. J. Virol. 2014, 88, 1318–1331. [Google Scholar] [CrossRef] [Green Version]
- Grellet, E.; L’Hôte, I.; Goulet, A.; Imbert, I. Replication of the Coronavirus Genome: A Paradox among Positive-Strand RNA Viruses. J. Biol. Chem. 2022, 298, 101923. [Google Scholar] [CrossRef]
- Drzewnioková, P.; Festa, F.; Panzarin, V.; Lelli, D.; Moreno, A.; Zecchin, B.; De Benedictis, P.; Leopardi, S. Best Molecular Tools to Investigate Coronavirus Diversity in Mammals: A Comparison. Viruses 2021, 13, 1975. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Huang, Y.; Yuen, K.Y. Coronavirus Diversity, Phylogeny and Interspecies Jumping. Exp. Biol. Med. 2009, 234, 1117–1127. [Google Scholar] [CrossRef] [Green Version]
- Peiris, J.S.M.; Guan, Y.; Yuen, K.Y. Severe Acute Respiratory Syndrome. Nat. Med. 2004, 10, S88–S97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zumla, A.; Hui, D.S.; Perlman, S. Middle East Respiratory Syndrome. Lancet 2015, 386, 995–1007. [Google Scholar] [CrossRef] [Green Version]
- Platto, S.; Zhou, J.; Wang, Y.; Wang, H.; Carafoli, E. Biodiversity Loss and COVID-19 Pandemic: The Role of Bats in the Origin and the Spreading of the Disease. Biochem. Biophys. Res. Commun. 2021, 538, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Aravena, M.; McKee, C.; Gamble, A.; Lunn, T.; Morris, A.; Snedden, C.E.; Yinda, C.K.; Port, J.R.; Buchholz, D.W.; Yeo, Y.Y.; et al. Ecology, Evolution and Spillover of Coronaviruses from Bats; Springer: New York, NY, USA, 2022; Volume 20, ISBN 4157902100652. [Google Scholar]
- Voigt, C.C.; Kingston, T. Bats in the Anthropocene: Conservation of Bats in a Changing World; Springer: Cham, Switzerland; London, UK; New York, NY, USA, 2016; p. 600. ISBN 9783319252186. [Google Scholar]
- O’Shea, T.J.; Cryan, P.M.; Cunningham, A.A.; Fooks, A.R.; Hayman, D.T.S.; Luis, A.D.; Peel, A.J.; Plowright, R.K.; Wood, J.L.N. Bat Flight and Zoonotic Viruses. Emerg. Infect. Dis. 2014, 20, 741–745. [Google Scholar] [CrossRef] [Green Version]
- Ahn, M.; Cui, J.; Irving, A.T.; Wang, L.F. Unique Loss of the PYHIN Gene Family in Bats amongst Mammals: Implications for Inflammasome Sensing. Sci. Rep. 2016, 6, 21722. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Singh, M.P.; Goyal, K.; Tripti, P.; Ansari, M.I.; Obli Rajendran, V.; Dhama, K.; Malik, Y.S. Bats and Viruses: A Death-Defying Friendship. Virusdisease 2021, 32, 467–479. [Google Scholar] [CrossRef]
- Kerth, G. Animal Sociality: Bat Colonies Are Founded by Relatives. Curr. Biol. 2008, 18, 740–742. [Google Scholar] [CrossRef] [Green Version]
- van Dam, A.; van Engelen, W.; Müller-Mahn, D.; Agha, S.; Junglen, S.; Borgemeister, C.; Bollig, M. Complexities of Multispecies Coexistence: Animal Diseases and Diverging Modes of Ordering at the Wildlife–Livestock Interface in Southern Africa. Environ. Plan. E Nat. Sp. 2023. [Google Scholar] [CrossRef]
- Johnson, C.K.; Hitchens, P.L.; Pandit, P.S.; Rushmore, J.; Evans, T.S.; Young, C.C.W.; Doyle, M.M. Global Shifts in Mammalian Population Trends Reveal Key Predictors of Virus Spillover Risk. Proc. R. Soc. B Biol. Sci. 2020, 287, 20192736. [Google Scholar] [CrossRef] [Green Version]
- Garrett, N.R.; Watkins, J.; Francis, C.; Simmons, N.B.; Ivanova, N.V.; Naaum, A.; Briscoe, A.; Drinkwater, R.; Clare, E.L. Out of Thin Air: Surveying Tropical Bat Roosts through Air Sampling of EDNA. PeerJ 2023, 11, e14772. [Google Scholar] [CrossRef] [PubMed]
- Cardiff, S.G.; Ratrimomanarivo, F.H.; Rembert, G.; Goodman, S.M. Hunting, Disturbance and Roost Persistence of Bats in Caves at Ankarana, Northern Madagascar. Afr. J. Ecol. 2009, 47, 640–649. [Google Scholar] [CrossRef]
- López-Roig, M.; Serra-Cobo, J. Impact of Human Disturbance, Density, and Environmental Conditions on the Survival Probabilities of Pipistrelle Bat (Pipistrellus pipistrellus). Popul. Ecol. 2014, 56, 471–480. [Google Scholar] [CrossRef]
- Speakman, J.R.; Webb, P.I.; Racey, P.A. Effects of Disturbance on the Energy Expenditure of Hibernating Bats. J. Appl. Ecol. 1991, 28, 1087–1104. [Google Scholar] [CrossRef]
- da Silva, P.G.; Nascimento, M.S.J.; Soares, R.R.G.; Sousa, S.I.V.; Mesquita, J.R. Airborne Spread of Infectious SARS-CoV-2: Moving Forward Using Lessons from SARS-CoV and MERS-CoV. Sci. Total Environ. 2021, 764, 142802. [Google Scholar] [CrossRef]
- Zhang, R.; Li, Y.; Zhang, A.L.; Wang, Y.; Molina, M.J. Identifying Airborne Transmission as the Dominant Route for the Spread of COVID-19. Proc. Natl. Acad. Sci. USA 2020, 117, 14857–14863. [Google Scholar] [CrossRef]
- Wilkinson, D.A.; Joffrin, L.; Lebarbenchon, C.; Mavingui, P. Analysis of Partial Sequences of the RNA-Dependent RNA Polymerase Gene as a Tool for Genus and Subgenus Classification of Coronaviruses. J. Gen. Virol. 2021, 101, 1261–1269. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Zhang, D.; Kan, X.; Huss, S.E.; Jiang, L.; Chen, L.Q.; Hu, Y. Using Phylogenetic Analysis to Investigate Eukaryotic Gene Origin. J. Vis. Exp. 2018, 2018, e56684. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v4: Recent Updates and New Developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Amengual, B.; López-Roig, M.; Serra-Cobo, J. First Record of Seasonal over Sea Migration of Miniopterus schreibersii and Myotis capaccinii between Balearic Islands (Spain). Acta Chiropterol. 2007, 9, 319–322. [Google Scholar] [CrossRef]
- Wong, S.; Lau, S.; Woo, P.; Yuen, K.Y. Bats as a Continuing Source of Emerging Infections in Humans. Rev. Med. Virol. 2007, 17, 67–91. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.S.; do Canto Olegário, J.; Weber, M.N.; da Silva, M.S.; Canova, R.; Sauthier, J.T.; Baumbach, L.F.; Witt, A.A.; Varela, A.P.M.; Mayer, F.Q.; et al. Detection of Coronavirus in Vampire Bats (Desmodus rotundus) in Southern Brazil. Transbound. Emerg. Dis. 2022, 69, 2384–2389. [Google Scholar] [CrossRef] [PubMed]
- Bonny, T.; Driver, J.; Paisie, T.; Salemi, M.; Morris, J.; Shender, L.; Smith, L.; Enloe, C.; Oxenrider, K.; Gore, J.; et al. Detection of Alphacoronavirus VRNA in the Feces of Brazilian Free-Tailed Bats (Tadarida brasiliensis) from a Colony in Florida, USA. Diseases 2017, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Zhang, A.; Sun, J.L.; Liu, H.; Hu, J.; Xu, L.X. Study of SARS Transmission via Liquid Droplets in Air. J. Biomech. Eng. 2005, 127, 32–38. [Google Scholar] [CrossRef]
- Xiao, W.; Wang, M.; Wei, W.; Wang, J.; Zhao, J.; Yi, B.; Li, J. [Detection of SARS-CoV and RNA on aerosol samples from SARS-patients admitted to hospital]. Zhonghua Liu Xing Bing Xue Za Zhi 2004, 25, 882–885. [Google Scholar]
- Pyankov, O.V.; Bodnev, S.A.; Pyankova, O.G.; Agranovski, I.E. Survival of Aerosolized Coronavirus in the Ambient Air. J. Aerosol Sci. 2018, 115, 158–163. [Google Scholar] [CrossRef]
- Kim, S.H.; Chang, S.Y.; Sung, M.; Park, J.H.; Bin Kim, H.; Lee, H.; Choi, J.P.; Choi, W.S.; Min, J.Y. Extensive Viable Middle East Respiratory Syndrome (MERS) Coronavirus Contamination in Air and Surrounding Environment in MERS Isolation Wards. Clin. Infect. Dis. 2016, 63, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Azhar, E.I.; Hashem, A.M.; El-Kafrawy, S.A.; Sohrab, S.S.; Aburizaiza, A.S.; Farraj, S.A.; Hassan, A.M.; Al-Saeed, M.S.; Jamjoom, G.A.; Madani, T.A. Detection of the Middle East Respiratory Syndrome Coronavirus Genome in an Air Sample Originating from a Camel Barn Owned by an Infected Patient. MBio 2014, 5, e01450-14. [Google Scholar] [CrossRef] [Green Version]
- da Silva, P.G.; Gonçalves, J.; Nascimento, M.S.; Sousa, S.I.V.; Mesquita, J.R. Detection of SARS-CoV-2 in the Indoor and Outdoor Areas of Urban Public Transport Systems of Three Major Cities of Portugal in 2021. Int. J. Environ. Res. Public Health 2022, 19, 5955. [Google Scholar]
- Lednicky, J.; Lauzard, M.; Fan, Z.H.; Jutla, A.; Tilly, T.; Gangwar, M.; Usmani, M.; Shankar, S.N.; Mohamed, K.; Eiguren-Fernandez, A.; et al. Viable SARS-CoV-2 in the Air of a Hospital Room with COVID-19 Patients. Int. J. Infect. Dis. 2020, 100, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Morawska, L.; Milton, D.K. It Is Time to Address Airborne Transmission of COVID-19. Clin. Infect. Dis. 2020, 71, 2311–2313. [Google Scholar] [CrossRef] [PubMed]
- WHO Team. WHO Coronavirus Disease (COVID-19): How Is It Transmitted? World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Silva, P.G.; Branco, P.T.B.S.; Soares, R.R.G.; Mesquita, J.R.; Sousa, S.I.V. SARS-CoV-2 Air Sampling: A Systematic Review on the Methodologies for Detection and Infectivity. Indoor Air 2022, 32, e13083. [Google Scholar] [CrossRef]
- da Silva, P.G.; Mesquita, J.R.; de São José Nascimento, M.; Ferreira, V.A.M. Viral, Host and Environmental Factors That Favor Anthropozoonotic Spillover of Coronaviruses: An Opinionated Review, Focusing on SARS-CoV, MERS-CoV and SARS-CoV-2. Sci. Total Environ. 2021, 750, 141483. [Google Scholar] [CrossRef] [PubMed]
- Breshears, L.E.; Nguyen, B.T.; Mata Robles, S.; Wu, L.; Yoon, J.Y. Biosensor Detection of Airborne Respiratory Viruses Such as SARS-CoV-2. SLAS Technol. 2022, 27, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Fan, H.; Lan, T.; Yang, X.L.; Shi, W.F.; Zhang, W.; Zhu, Y.; Zhang, Y.W.; Xie, Q.M.; Mani, S.; et al. Fatal Swine Acute Diarrhoea Syndrome Caused by an HKU2-Related Coronavirus of Bat Origin. Nature 2018, 556, 255–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
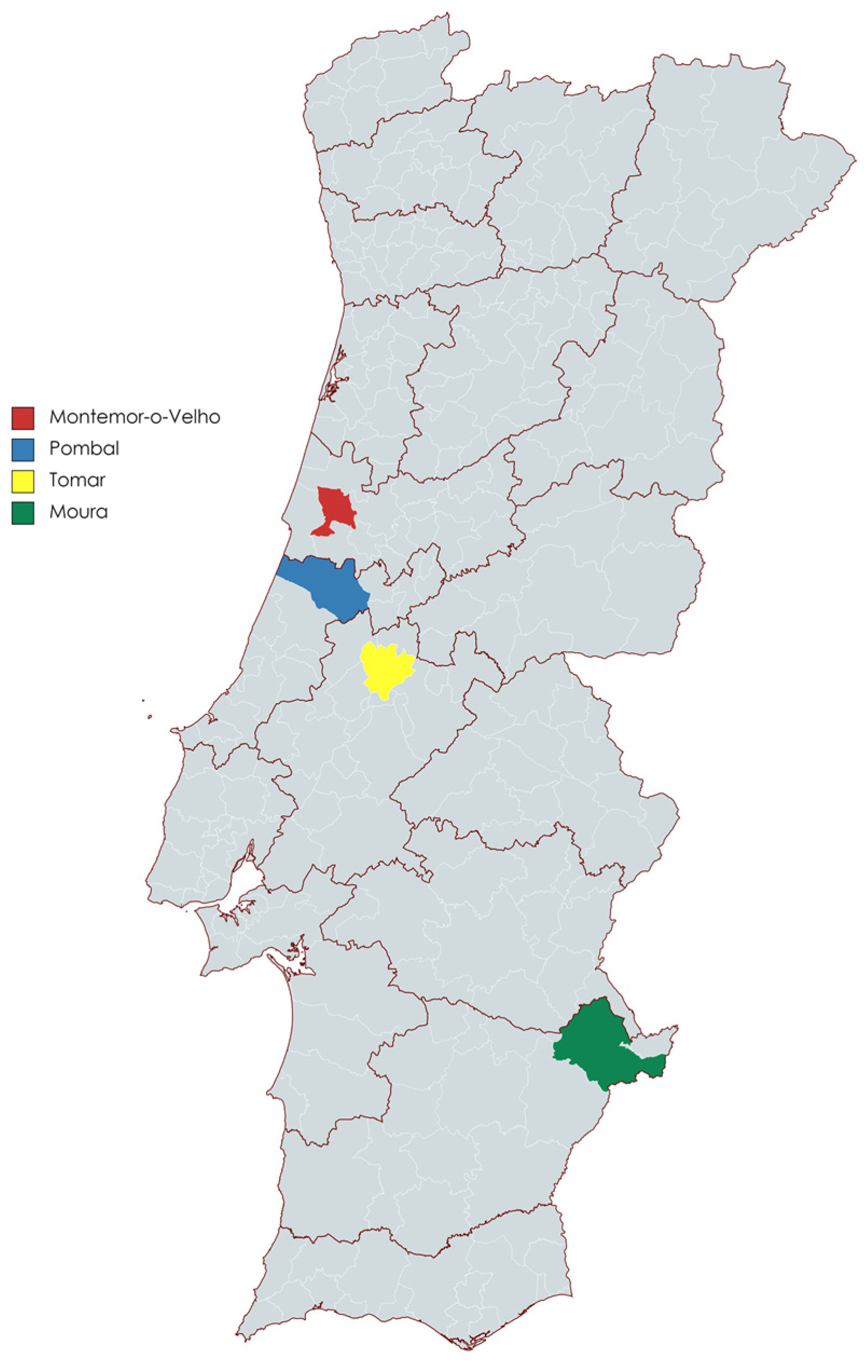
| Location | Species | Number of Individuals Found in Each Location | Matrices Collected | ||
|---|---|---|---|---|---|
| Stool Samples | Anal Swab | Buccal Swab | |||
| Pombal | Myotis myotis | 1 | 0 | 1 | 1 |
| Tomar | Miniopterus schreibersii | 7 | 1 | 7 | 7 |
| Rhinolophus mehelyi | 2 | 1 | 2 | 2 | |
| Myotis myotis | 5 | 3 | 5 | 5 | |
| Moura | Miniopterus schreibersii | 7 | 2 | 7 | 7 |
| Rhinolophus mehelyi | 6 | 1 | 6 | 6 | |
| Myotis myotis | 9 | 4 | 9 | 9 | |
| Rhinolophus ferrumequinum | 5 | 2 | 5 | 5 | |
| Total | 42 | 14 | 42 | 42 | |
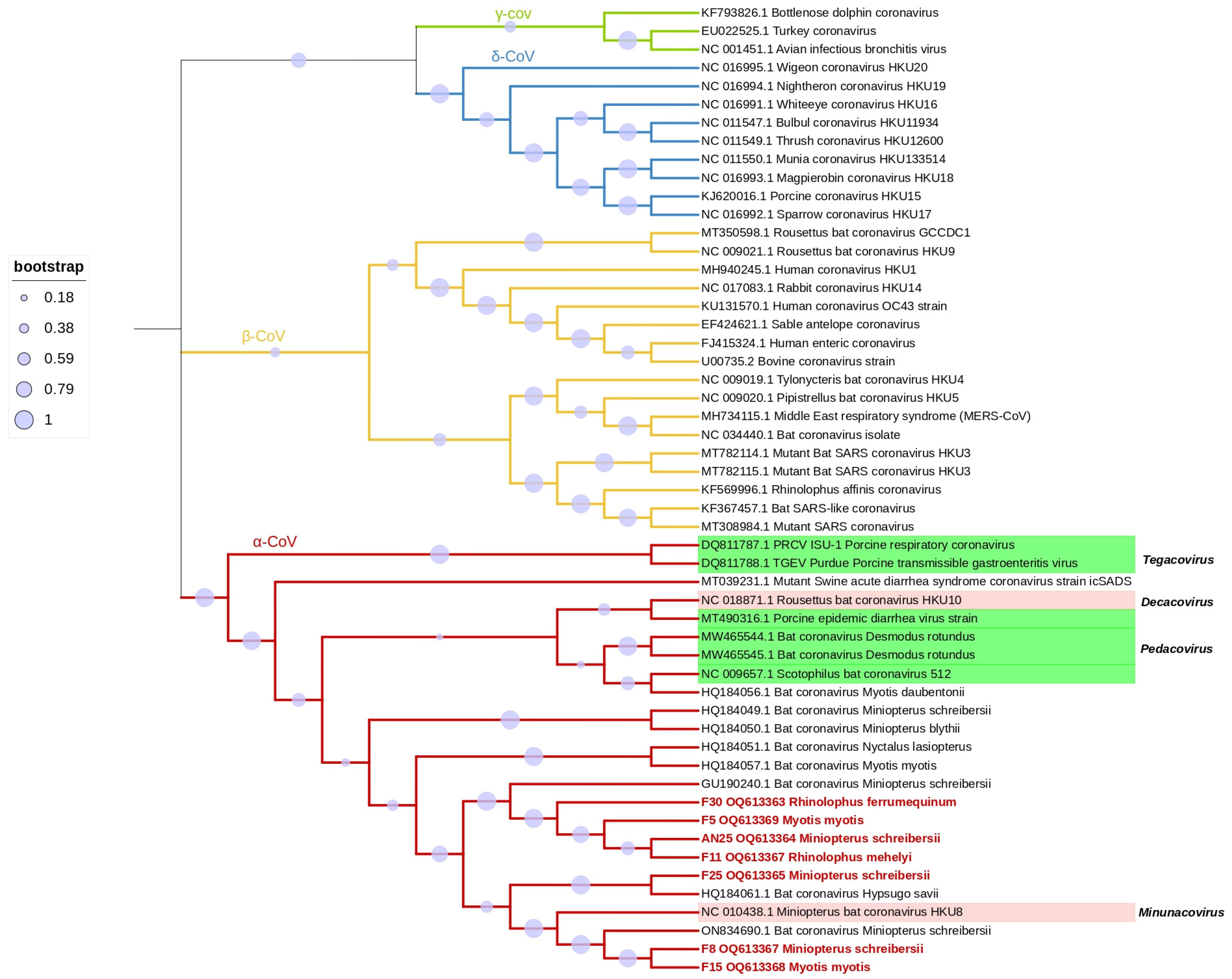
| Collection Site | Sample ID | Host Species | Acession Number | Shared Identity |
|---|---|---|---|---|
| Tomar | F5 | Myotis myotis | OQ613369 | Miniopterus schreibersii-Bulgaria-GU190240.1 |
| F8 | Miniopterus schreibersii | OQ613367 | Miniopterus schreibersii-Italy-ON834690.1 | |
| F11 | Rhinolophus mehelyi | OQ613367 | Miniopterus schreibersii-Bulgaria-GU190240.1 | |
| F15 | Myotis. Myotis | OQ613368 | Miniopterus schreibersii-Bulgaria-GU190240.1 | |
| Moura | F25 | Miniopterus schreibersii | OQ613365 | Hypsugo savii-Spain-HQ184061.1 |
| AN25 | Miniopterus schreibersii | OQ613364 | Miniopterus schreibersii-Bulgaria-GU190240.1 | |
| F30 | Rhinolophus ferrumequinum | OQ613363 | Miniopterus schreibersii-Bulgaria-GU190240.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hemnani, M.; Silva, P.G.d.; Thompson, G.; Poeta, P.; Rebelo, H.; Mesquita, J.R. First Report of Alphacoronavirus Circulating in Cavernicolous Bats from Portugal. Viruses 2023, 15, 1521. https://doi.org/10.3390/v15071521
Hemnani M, Silva PGd, Thompson G, Poeta P, Rebelo H, Mesquita JR. First Report of Alphacoronavirus Circulating in Cavernicolous Bats from Portugal. Viruses. 2023; 15(7):1521. https://doi.org/10.3390/v15071521
Chicago/Turabian StyleHemnani, Mahima, Priscilla Gomes da Silva, Gertrude Thompson, Patricia Poeta, Hugo Rebelo, and João R. Mesquita. 2023. "First Report of Alphacoronavirus Circulating in Cavernicolous Bats from Portugal" Viruses 15, no. 7: 1521. https://doi.org/10.3390/v15071521