
Bergamottin Inhibits PRRSV Replication by Blocking Viral Non-Structural Proteins Expression and Viral RNA Synthesis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses, and Bergamottin
2.2. Cytotoxicity Assay
2.3. Quantitative Real-Time Reverse-Transcription Polymerase Chain Reaction (RT-qPCR)
2.4. Western Blot
2.5. Expression Vector Construction and Transfection
2.6. Viral Binding, Entry, Replication, and Release Assays
2.7. Immunofluorescence Assay
2.8. Detection of Proinflammatory Cytokines and Interferon
2.9. Statistical Analysis
3. Results
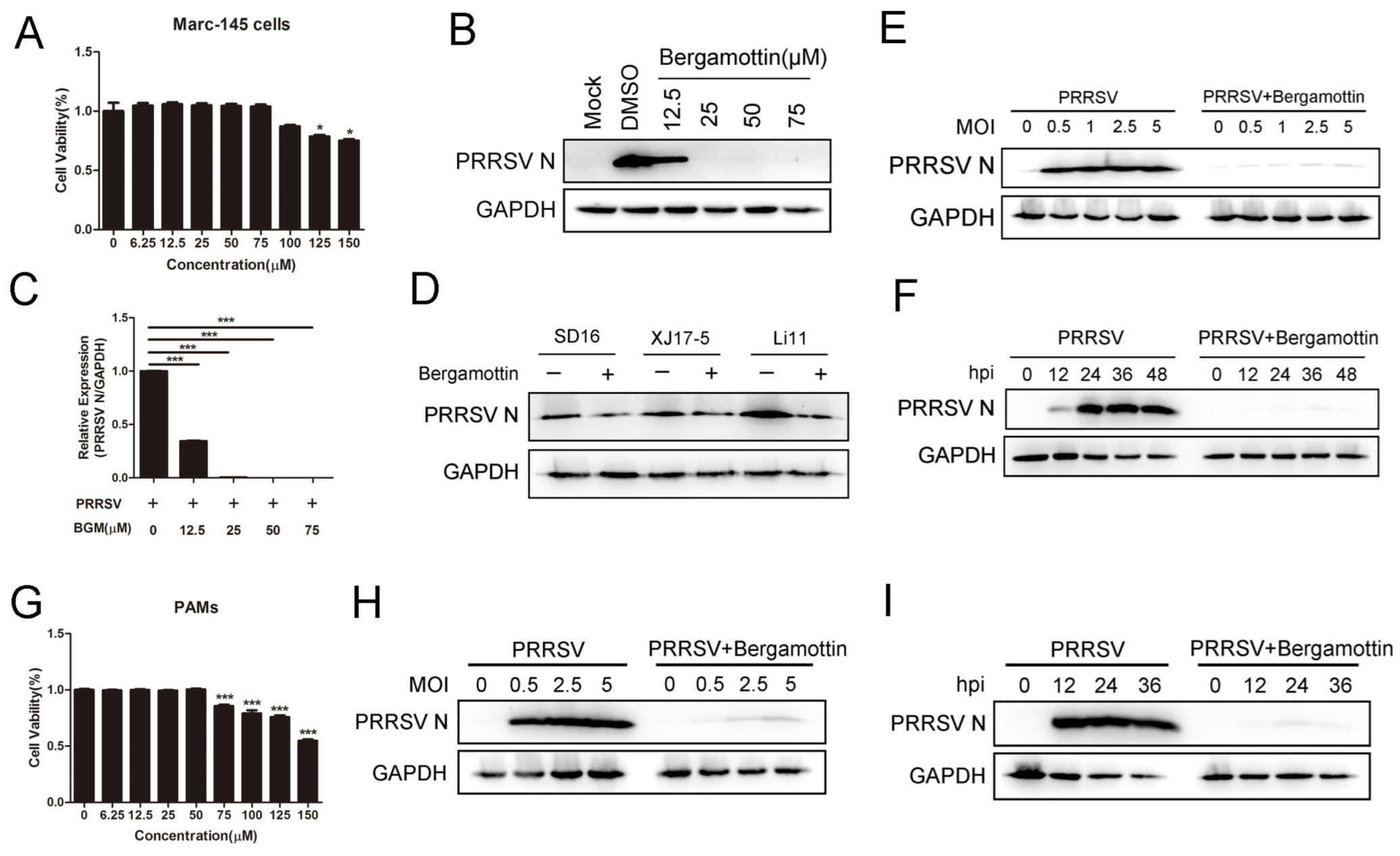
3.1. Bergamottin Inhibits PRRSV Replication
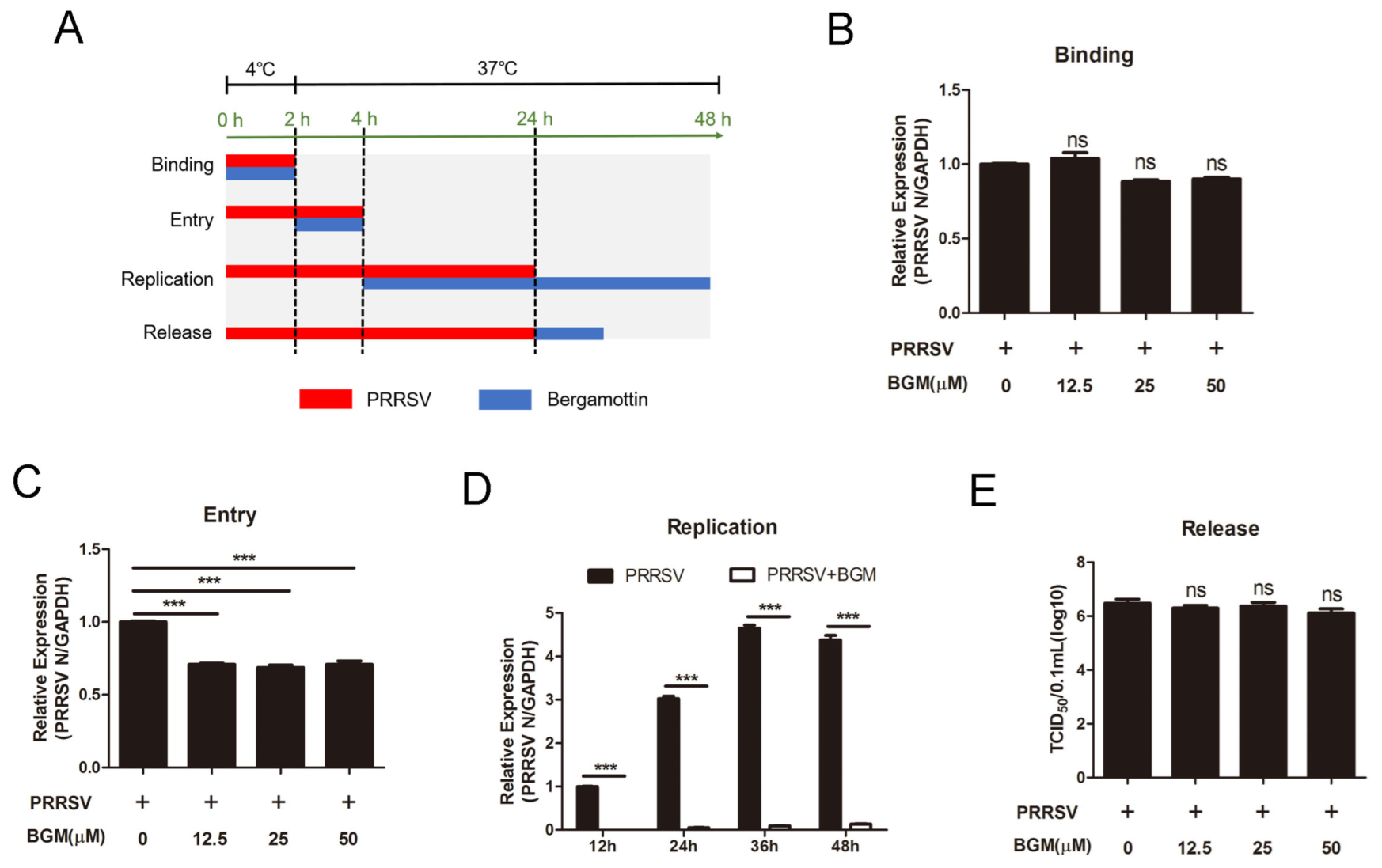
3.2. Bergamottin Blocks PRRSV Entry and Genome Replication but Had No Effects on Viral Binding or Release
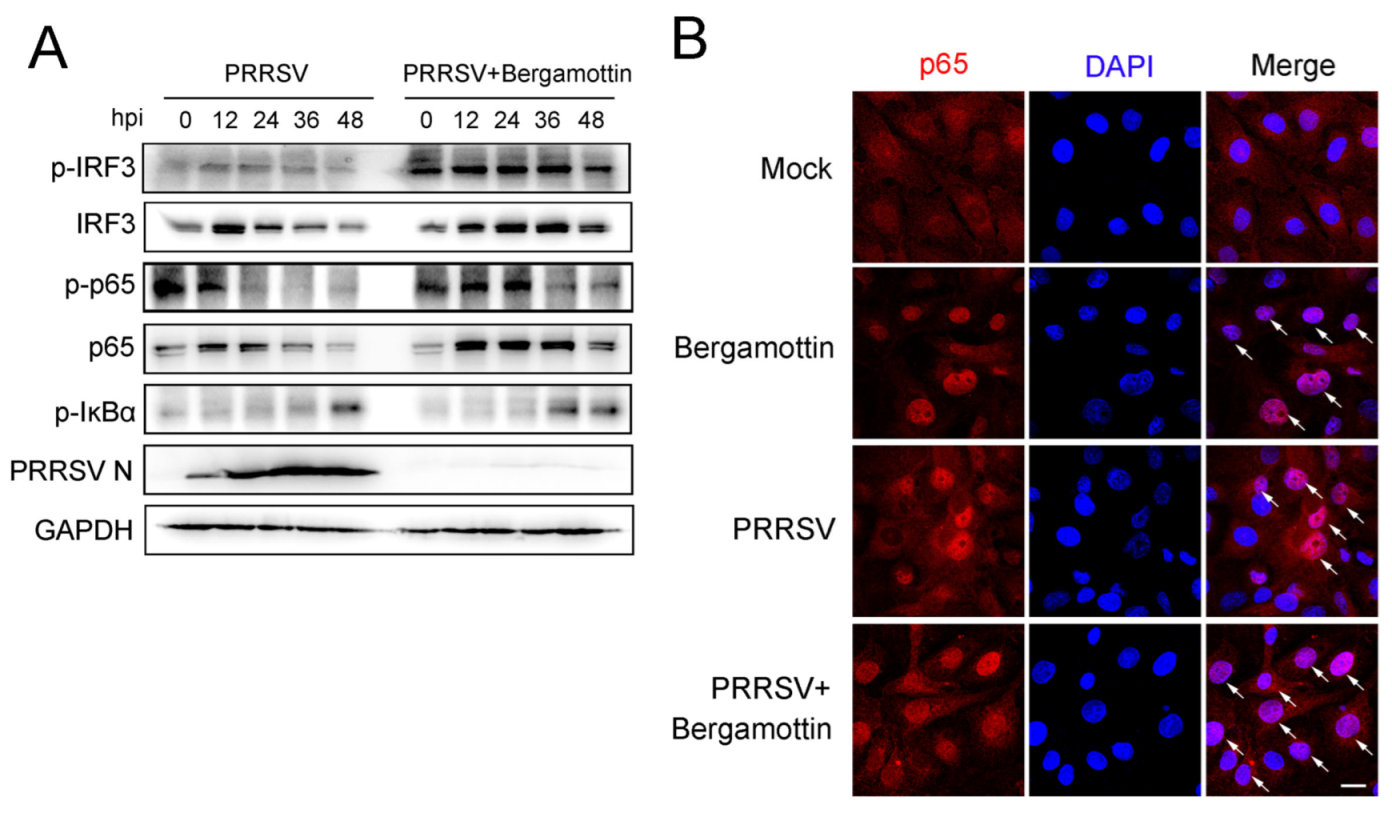
3.3. Bergamottin Activates IRF3 and NF-κB Signaling Pathway
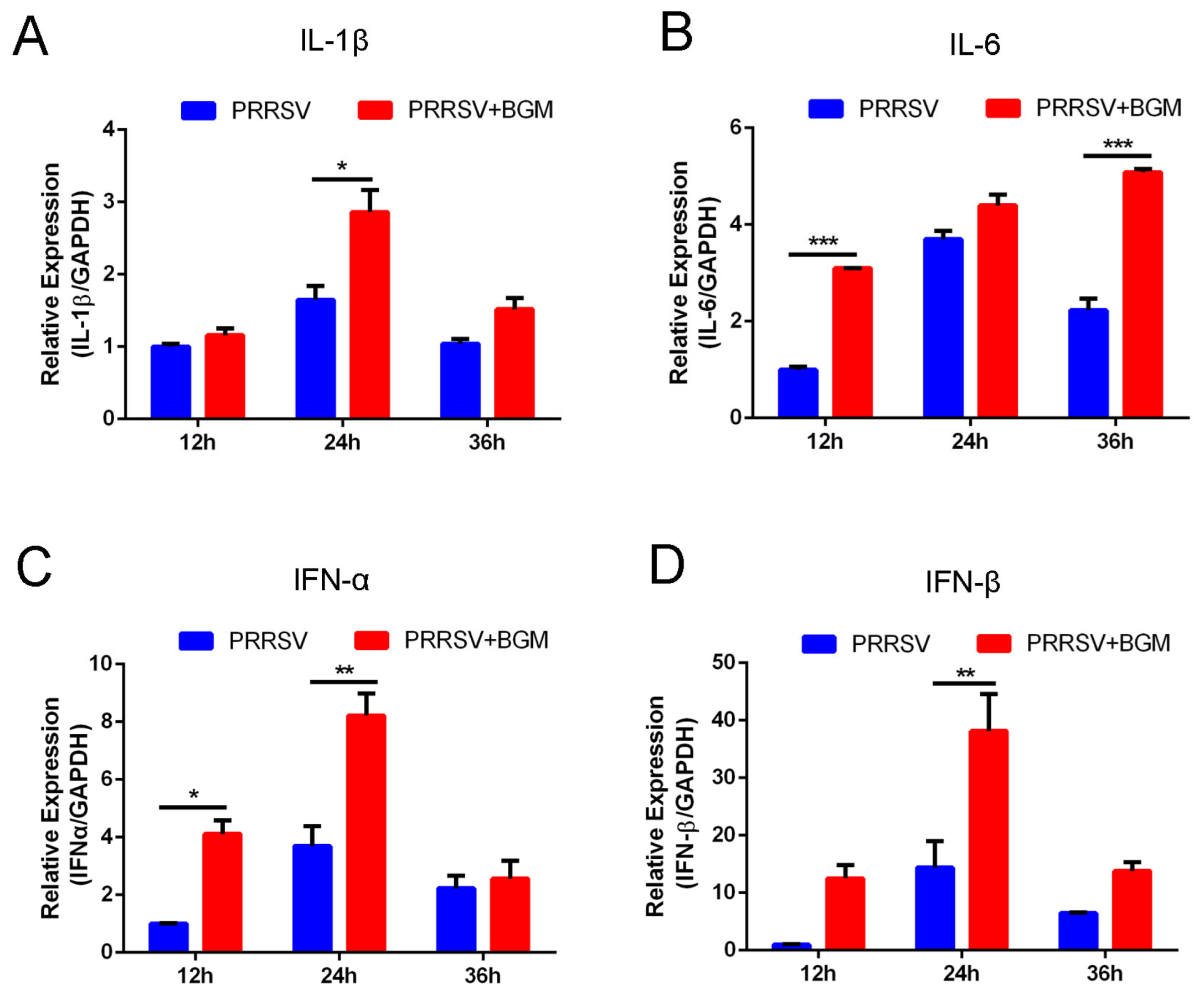
3.4. Bergamottin Promotes the Expression of Proinflammatory Cytokines and Type I Interferon
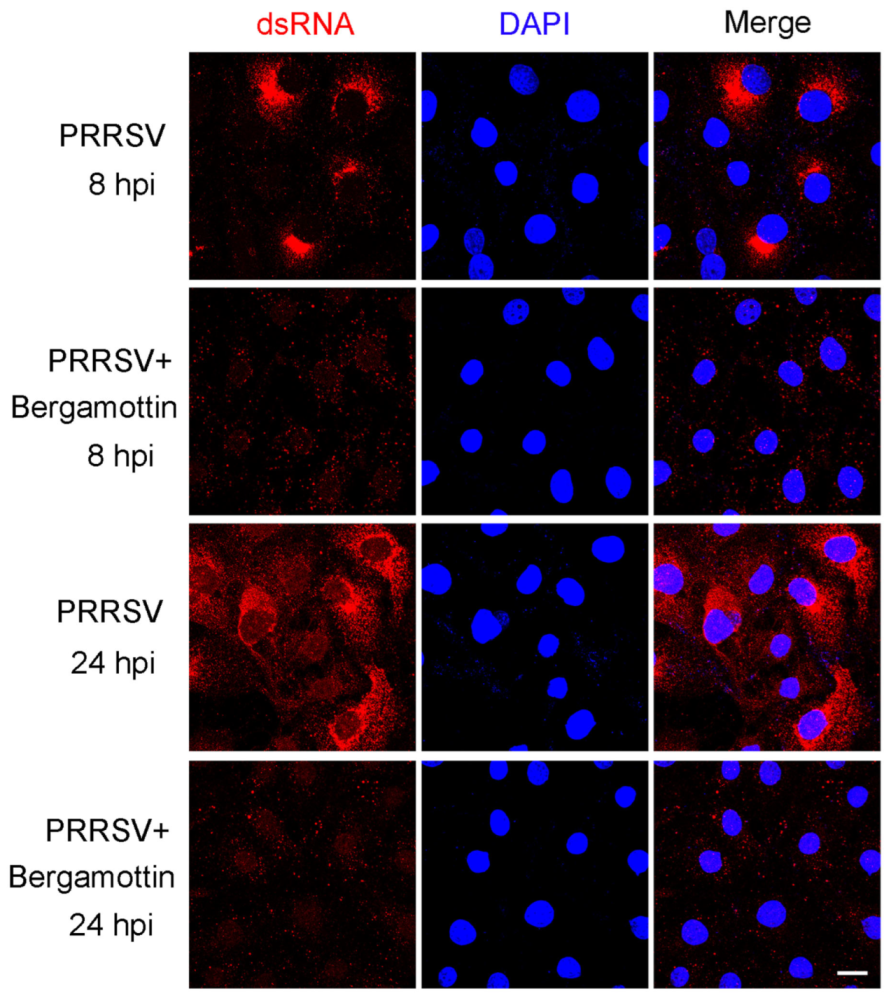
3.5. Bergamottin Inhibits Viral RNA Synthesis
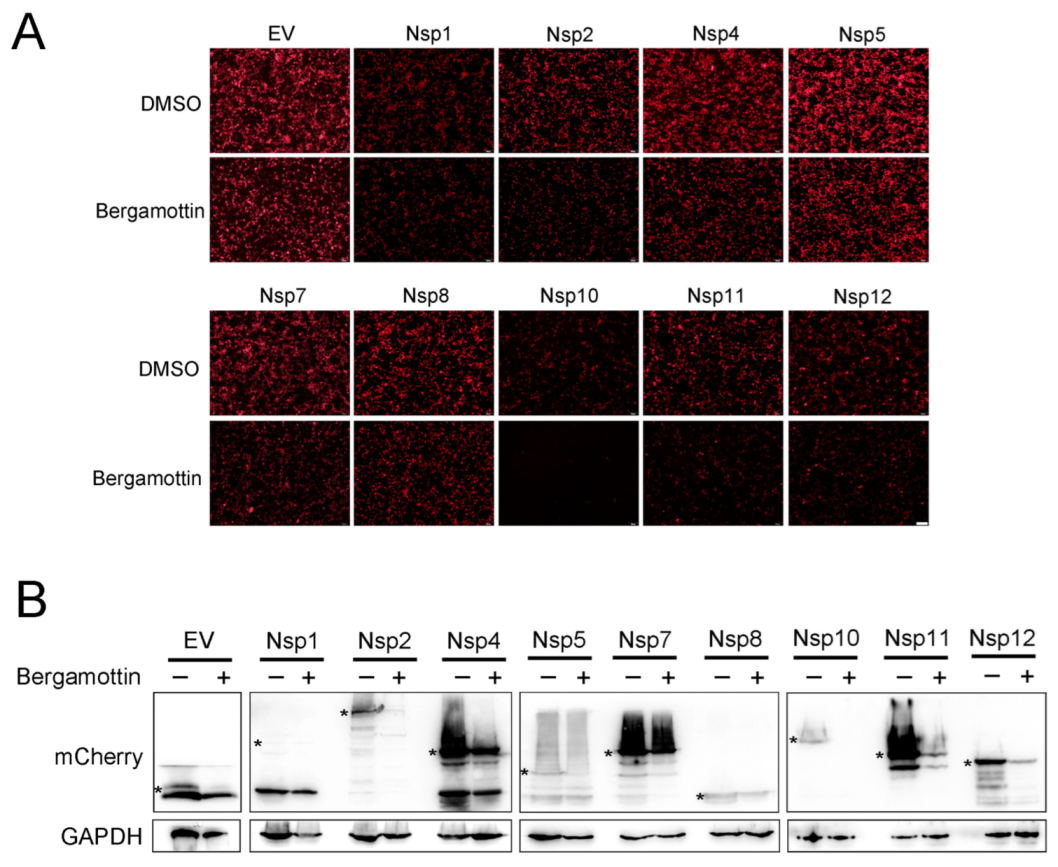
3.6. Bergamottin Declines Most Nsp Expression and Completely Inhibited Nsp10
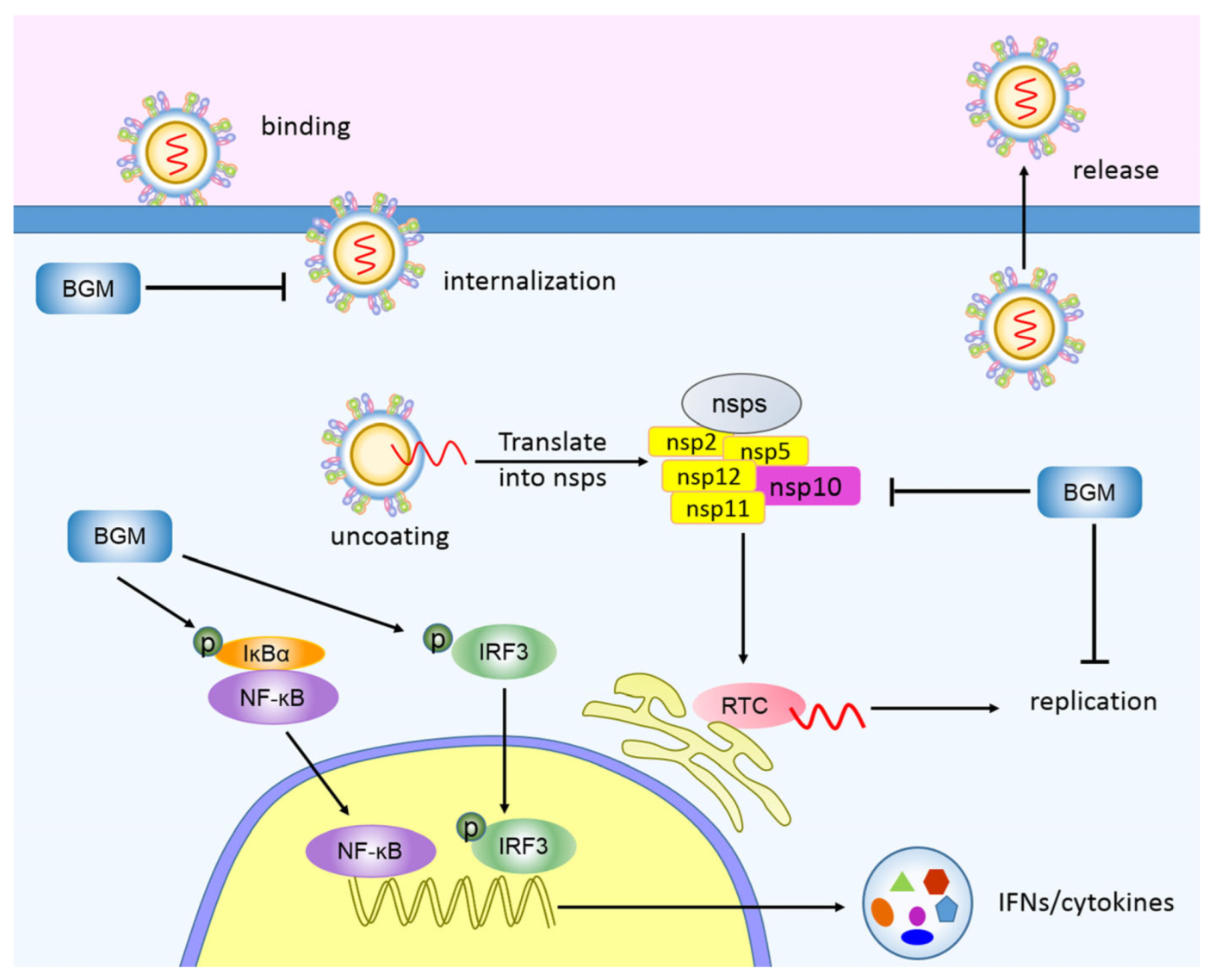
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lunney, J.K.; Benfield, D.A.; Rowland, R.R. Porcine reproductive and respiratory syndrome virus: An update on an emerging and re-emerging viral disease of swine. Virus Res. 2010, 154, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Zhou, L.; Ge, X.; Guo, X.; Yang, H. Pathogenesis and control of the Chinese highly pathogenic porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 2017, 209, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.G.; Dee, S.A. Porcine reproductive and respiratory syndrome virus. Theriogenology 2006, 66, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Music, N.; Gagnon, C.A. The role of porcine reproductive and respiratory syndrome (PRRS) virus structural and non-structural proteins in virus pathogenesis. Anim. Health Res. Rev. 2010, 11, 135–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montaner-Tarbes, S.; Del Portillo, H.A.; Montoya, M.; Fraile, L. Key Gaps in the Knowledge of the Porcine Respiratory Reproductive Syndrome Virus (PRRSV). Front. Vet. Sci. 2019, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef]
- Fang, Y.; Snijder, E.J. The PRRSV replicase: Exploring the multifunctionality of an intriguing set of nonstructural proteins. Virus Res. 2010, 154, 61–76. [Google Scholar] [CrossRef]
- Yun, S.I.; Lee, Y.M. Overview: Replication of porcine reproductive and respiratory syndrome virus. J. Microbiol. 2013, 51, 711–723. [Google Scholar] [CrossRef]
- Yuan, S.; Zhang, N.; Xu, L.; Zhou, L.; Ge, X.; Guo, X.; Yang, H. Induction of Apoptosis by the Nonstructural Protein 4 and 10 of Porcine Reproductive and Respiratory Syndrome Virus. PLoS ONE 2016, 11, e0156518. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhou, L.; Zhang, J.; Ge, X.; Zhou, R.; Zheng, H.; Geng, G.; Guo, X.; Yang, H. Nsp9 and Nsp10 contribute to the fatal virulence of highly pathogenic porcine reproductive and respiratory syndrome virus emerging in China. PLoS Pathog. 2014, 10, e1004216. [Google Scholar] [CrossRef]
- Huang, C.; Zhang, Q.; Feng, W.H. Regulation and evasion of antiviral immune responses by porcine reproductive and respiratory syndrome virus. Virus Res. 2015, 202, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.L.X.; Pattnaik, A.K.; Osorio, F.A. Strategies to broaden the cross-protective efficacy of vaccines against porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 2017, 206, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Du, T.; Nan, Y.; Xiao, S.; Zhao, Q.; Zhou, E.M. Antiviral Strategies against PRRSV Infection. Trends Microbiol. 2017, 25, 968–979. [Google Scholar] [CrossRef] [PubMed]
- Villalaín, J. Bergamottin: Location, aggregation and interaction with the plasma membrane. J. Biomol. Struct. Dyn. 2023, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.H.; Arfuso, F.; Sethi, G.; Ahn, K.S. Pharmacological Utilization of Bergamottin, Derived from Grapefruits, in Cancer Prevention and Therapy. Int. J. Mol. Sci. 2018, 19, 4048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Liao, N. Bergamottin Induces DNA Damage and Inhibits Malignant Progression in Melanoma by Modulating miR-145/Cyclin D1 Axis. OncoTargets Ther. 2021, 14, 3769–3781. [Google Scholar] [CrossRef]
- Kim, S.M.; Lee, J.H.; Sethi, G.; Kim, C.; Baek, S.H.; Nam, D.; Chung, W.S.; Kim, S.H.; Shim, B.S.; Ahn, K.S. Bergamottin, a natural furanocoumarin obtained from grapefruit juice induces chemosensitization and apoptosis through the inhibition of STAT3 signaling pathway in tumor cells. Cancer Lett. 2014, 354, 153–163. [Google Scholar] [CrossRef]
- Vetrichelvan, O.; Gorjala, P.; Goodman, O., Jr.; Mitra, R. Bergamottin a CYP3A inhibitor found in grapefruit juice inhibits prostate cancer cell growth by downregulating androgen receptor signaling and promoting G0/G1 cell cycle block and apoptosis. PLoS ONE 2021, 16, e0257984. [Google Scholar] [CrossRef]
- Wang, X.; Tian, Y.; Liang, X.; Yin, C.; Huai, Y.; Zhao, Y.; Huang, Q.; Chu, X.; Wang, W.; Qian, A. Bergamottin promotes osteoblast differentiation and bone formation via activating the Wnt/β-catenin signaling pathway. Food Funct. 2022, 13, 2913–2924. [Google Scholar] [CrossRef]
- Jung, Y.Y.; Ko, J.H.; Um, J.Y.; Sethi, G.; Ahn, K.S. A Novel Role of Bergamottin in Attenuating Cancer Associated Cachexia by Diverse Molecular Mechanisms. Cancers 2021, 13, 1347. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, S.; Lin, T. Bergamottin exerts anticancer effects on human colon cancer cells via induction of apoptosis, G2/M cell cycle arrest and deactivation of the Ras/Raf/ERK signalling pathway. Arch. Med. Sci. AMS 2022, 18, 1572–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Zhang, Y.; Zeb, A.; Wu, Y.; Cheng, L. Bergamottin and PAP-1 Induced ACE2 Degradation to Alleviate Infection of SARS-CoV-2. Int. J. Mol. Sci. 2022, 23, 12565. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Liu, Y.; Cao, J.; Dong, S.; Hou, Y.; Yu, Y.; Zhang, Q.; Zhang, Y.; Jia, X.; Zhang, B.; et al. Bergamottin, a bioactive component of bergamot, inhibits SARS-CoV-2 infection in golden Syrian hamsters. Antivir. Res. 2022, 204, 105365. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, J.; Cao, J.; Zhang, G.; Jia, X.; Wang, P.; Xiao, G.; Wang, W. Screening of Botanical Drugs against Lassa Virus Entry. J. Virol. 2021, 95, e02429-20. [Google Scholar] [CrossRef]
- Zhao, Y.; Song, Z.; Bai, J.; Liu, X.; Nauwynck, H.; Jiang, P. ZAP, a CCCH-Type Zinc Finger Protein inhibits PRRSV replication and interacts with viral Nsp9. J. Virol. 2019, 93, e00001-19. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Zhou, Y.; Tong, G. Mechanisms of suppression of interferon production by porcine reproductive and respiratory syndrome virus. Acta Virol. 2012, 56, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sha, H.; Zhang, H.; Chen, Y.; Huang, L.; Zhao, M.; Wang, N. Research Progress on the NSP9 Protein of Porcine Reproductive and Respiratory Syndrome Virus. Front. Vet. Sci. 2022, 9, 872205. [Google Scholar] [CrossRef]
- Li, R.; Qiao, S.; Zhang, G. Reappraising host cellular factors involved in attachment and entry to develop antiviral strategies against porcine reproductive and respiratory syndrome virus. Front. Microbiol. 2022, 13, 975610. [Google Scholar] [CrossRef]
- Zhang, Q.; Yoo, D. PRRS virus receptors and their role for pathogenesis. Vet. Microbiol. 2015, 177, 229–241. [Google Scholar] [CrossRef]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479–480, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Yoo, D. Engineering the PRRS virus genome: Updates and perspectives. Vet. Microbiol. 2014, 174, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.X.; Song, N.; Chen, L.Z.; Cheng, S.P.; Wu, H.; Wen, Y.J. Non-structural protein 2 of the porcine reproductive and respiratory syndrome (PRRS) virus: A crucial protein in viral pathogenesis, immunity and diagnosis. Res. Vet. Sci. 2013, 95, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Liu, L.; Wu, W.; Armstrong, S.D.; Xia, D.; Nan, H.; Hiscox, J.A.; Chen, H. Determination of the interactome of non-structural protein12 from highly pathogenic porcine reproductive and respiratory syndrome virus with host cellular proteins using high throughput proteomics and identification of HSP70 as a cellular factor for virus replication. J. Proteom. 2016, 146, 58–69. [Google Scholar] [CrossRef]
- Chen, X.X.; Qiao, S.; Li, R.; Wang, J.; Li, X.; Zhang, G. Evasion strategies of porcine reproductive and respiratory syndrome virus. Front. Microbiol. 2023, 14, 1140449. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Z.; Xu, Y.; Chen, L.; Zhang, M.; Li, X. Bergamottin Inhibits PRRSV Replication by Blocking Viral Non-Structural Proteins Expression and Viral RNA Synthesis. Viruses 2023, 15, 1367. https://doi.org/10.3390/v15061367
Zhu Z, Xu Y, Chen L, Zhang M, Li X. Bergamottin Inhibits PRRSV Replication by Blocking Viral Non-Structural Proteins Expression and Viral RNA Synthesis. Viruses. 2023; 15(6):1367. https://doi.org/10.3390/v15061367
Chicago/Turabian StyleZhu, Zhenbang, Yuqian Xu, Lulu Chen, Meng Zhang, and Xiangdong Li. 2023. "Bergamottin Inhibits PRRSV Replication by Blocking Viral Non-Structural Proteins Expression and Viral RNA Synthesis" Viruses 15, no. 6: 1367. https://doi.org/10.3390/v15061367