
An Update on Current Antiviral Strategies to Combat Human Cytomegalovirus Infection



Abstract
:1. Introduction
2. Antiviral Drugs against HCMV
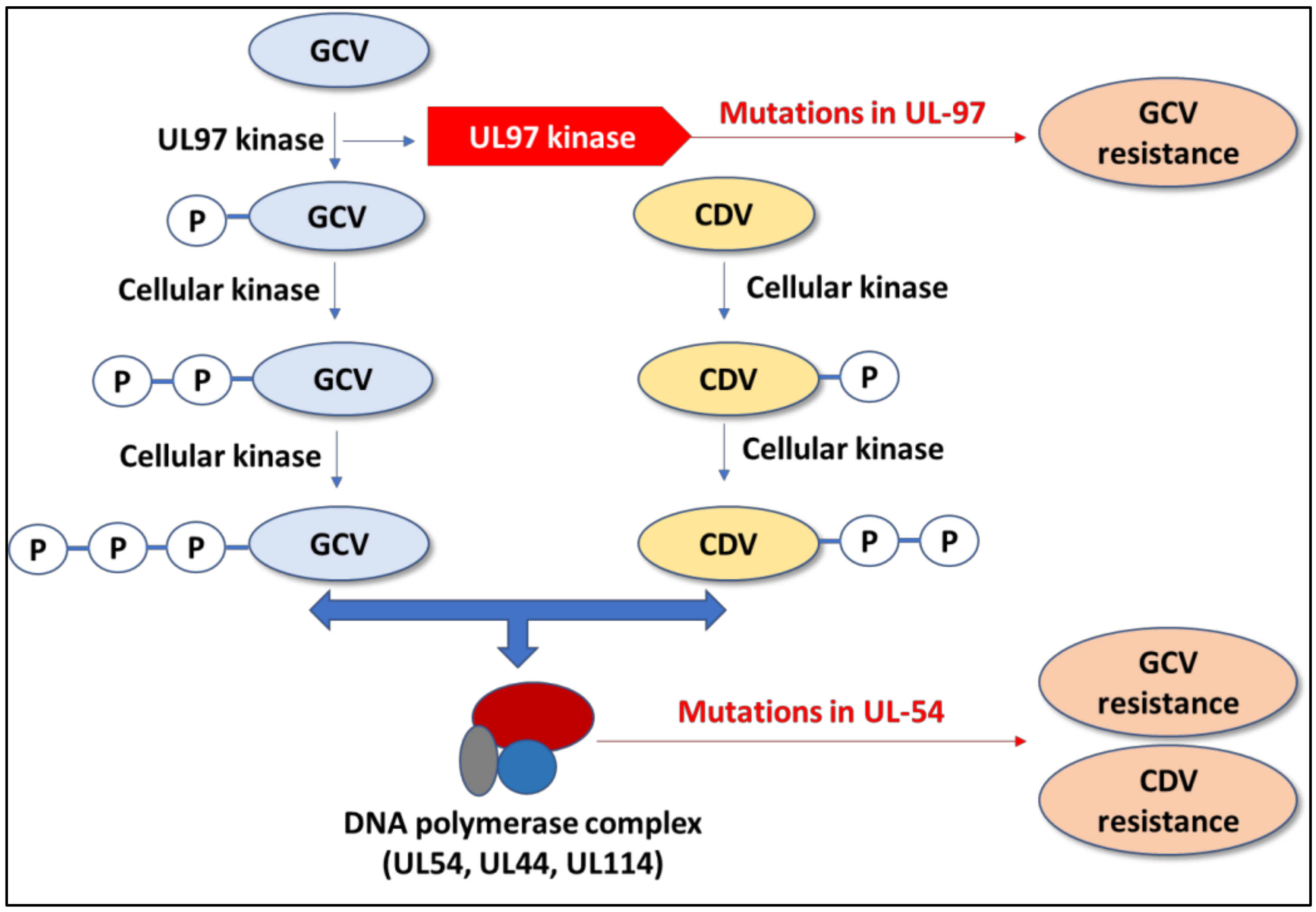
2.1. Ganciclovir and Valganciclovir
2.2. Cidofovir
2.3. Foscarnet
2.4. Letermovir
2.5. Maribavir
2.6. Repurposed Drugs
3. Cytomegalovirus Vaccines: Current Status and Future Prospects
4. Possible Alternative Strategies to Combat HCMV Infection
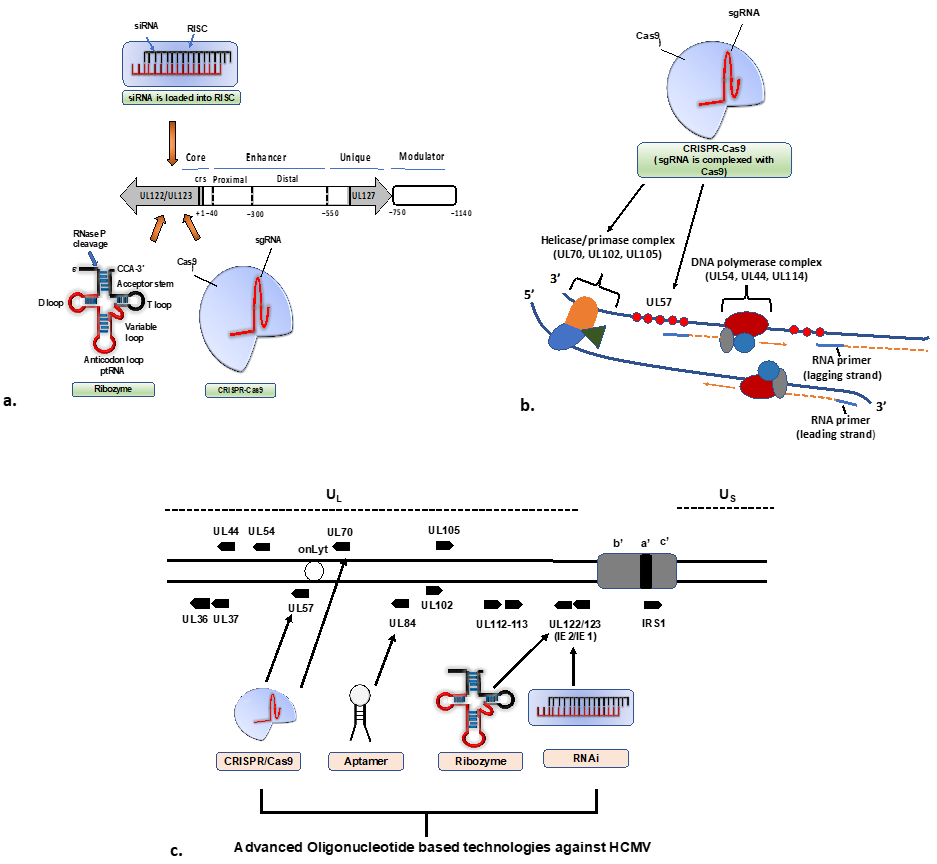
4.1. RNAi-Based Therapeutics against CMV
4.2. Ribozyme-Based Therapeutics
4.3. CRISPR/Cas9-Based Therapeutics
4.4. TALEN-Based Therapeutics
4.5. Aptamer-Based Therapeutics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gatherer, D.; Depledge, D.P.; Hartley, C.A.; Szpara, M.L.; Vaz, P.K.; Benkő, M.; Brandt, C.R.; Bryant, N.A.; Dastjerdi, A.; Doszpoly, A. ICTV virus taxonomy profile: Herpesviridae 2021. J. Gen. Virol. 2021, 102. [Google Scholar] [CrossRef] [PubMed]
- Goodrum, F. Human cytomegalovirus latency: Approaching the Gordian knot. Annu. Rev. Virol. 2016, 3, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fields, B.N. Fields’ Virology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 1. [Google Scholar]
- Boeckh, M.; Geballe, A.P. Cytomegalovirus: Pathogen, paradigm, and puzzle. J. Clin. Investig. 2011, 121, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Lancini, D.; Faddy, H.M.; Flower, R.; Hogan, C. Cytomegalovirus disease in immunocompetent adults. Med. J. Aust. 2014, 201, 578–580. [Google Scholar] [CrossRef] [Green Version]
- Al Mana, H.; Yassine, H.M.; Younes, N.N.; Al-Mohannadi, A.; Al-Sadeq, D.W.; Alhababi, D.; Nasser, E.A.; Nasrallah, G.K. The current status of cytomegalovirus (CMV) prevalence in the MENA region: A systematic review. Pathogens 2019, 8, 213. [Google Scholar] [CrossRef] [Green Version]
- Zuhair, M.; Smit, G.S.A.; Wallis, G.; Jabbar, F.; Smith, C.; Devleesschauwer, B.; Griffiths, P. Estimation of the worldwide seroprevalence of cytomegalovirus: A systematic review and meta-analysis. Rev. Med. Virol. 2019, 29, e2034. [Google Scholar] [CrossRef] [Green Version]
- Cannon, M.J.; Schmid, D.S.; Hyde, T.B. Review of cytomegalovirus seroprevalence and demographic characteristics associated with infection. Rev. Med. Virol. 2010, 20, 202–213. [Google Scholar] [CrossRef]
- Chakravarti, A.; Kashyap, B.; Matlani, M. Cytomegalovirus infection: An Indian perspective. Indian J. Med. Microbiol. 2009, 27, 3–11. [Google Scholar] [CrossRef]
- Manicklal, S.; Emery, V.C.; Lazzarotto, T.; Boppana, S.B.; Gupta, R.K. The “silent” global burden of congenital cytomegalovirus. Clin. Microbiol. Rev. 2013, 26, 86–102. [Google Scholar] [CrossRef] [Green Version]
- Landolfo, S.; Gariglio, M.; Gribaudo, G.; Lembo, D. The human cytomegalovirus. Pharmacol. Ther. 2003, 98, 269–297. [Google Scholar] [CrossRef]
- Mancebo, F.J.; Parras-Moltó, M.; García-Ríos, E.; Pérez-Romero, P. Deciphering the Potential Coding of Human Cytomegalovirus: New Predicted Transmembrane Proteome. Int. J. Mol. Sci. 2022, 23, 2768. [Google Scholar] [CrossRef]
- Stern-Ginossar, N.; Weisburd, B.; Michalski, A.; Le, V.T.K.; Hein, M.Y.; Huang, S.-X.; Ma, M.; Shen, B.; Qian, S.-B.; Hengel, H. Decoding human cytomegalovirus. Science 2012, 338, 1088–1093. [Google Scholar] [CrossRef] [Green Version]
- Varnum, S.M.; Streblow, D.N.; Monroe, M.E.; Smith, P.; Auberry, K.J.; Paša-Tolić, L.; Wang, D.; Camp, D.G.; Rodland, K.; Wiley, S. Identification of proteins in human cytomegalovirus (HCMV) particles: The HCMV proteome. Virol. J. 2004, 78, 10960–10966. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yu, J.; Liu, Z. MicroRNAs expressed by human cytomegalovirus. Virol. J. 2020, 17, 34. [Google Scholar] [CrossRef] [Green Version]
- Murphy, E.; Shenk, T.E. Human cytomegalovirus genome. Hum. Cytomegal. 2008, 325, 1–19. [Google Scholar]
- Yu, X.; Jih, J.; Jiang, J.; Zhou, Z.H. Atomic structure of the human cytomegalovirus capsid with its securing tegument layer of pp150. Science 2017, 356. [Google Scholar] [CrossRef] [Green Version]
- Schottstedt, V.; Blümel, J.; Burger, R.; Drosten, C.; Gröner, A.; Gürtler, L.; Heiden, M.; Hildebrandt, M.; Jansen, B.; Montag-Lessing, T. Human cytomegalovirus (HCMV)–revised. Transfus. Med. Hemother. 2010, 37, 365. [Google Scholar]
- Schauflinger, M.; Villinger, C.; Mertens, T.; Walther, P.; von Einem, J. Analysis of human cytomegalovirus secondary envelopment by advanced electron microscopy. Cell. Microbiol. 2013, 15, 305–314. [Google Scholar] [CrossRef]
- Mach, M.; Kropff, B.; Dal Monte, P.; Britt, W. Complex formation by human cytomegalovirus glycoproteins M (gpUL100) and N (gpUL73). Virol. J. 2000, 74, 11881–11892. [Google Scholar] [CrossRef] [Green Version]
- Shimamura, M.; Mach, M.; Britt, W.J. Human cytomegalovirus infection elicits a glycoprotein M (gM)/gN-specific virus-neutralizing antibody response. Virol. J. 2006, 80, 4591–4600. [Google Scholar] [CrossRef] [Green Version]
- Kropff, B.; Burkhardt, C.; Schott, J.; Nentwich, J.; Fisch, T.; Britt, W.; Mach, M. Glycoprotein N of human cytomegalovirus protects the virus from neutralizing antibodies. PLoS Pathog. 2012, 8, e1002999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Shenk, T. Human cytomegalovirus virion protein complex required for epithelial and endothelial cell tropism. Proc. Natl. Acad. Sci. USA 2005, 102, 18153–18158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciferri, C.; Chandramouli, S.; Leitner, A.; Donnarumma, D.; Cianfrocco, M.A.; Gerrein, R.; Friedrich, K.; Aggarwal, Y.; Palladino, G.; Aebersold, R. Antigenic characterization of the HCMV gH/gL/gO and pentamer cell entry complexes reveals binding sites for potently neutralizing human antibodies. PLoS Pathog. 2015, 11, e1005230. [Google Scholar] [CrossRef] [PubMed]
- Sinzger, C.; Digel, M.; Jahn, G. Cytomegalovirus cell tropism. Hum. Cytomegal. 2008, 325, 63–83. [Google Scholar]
- Gugliesi, F.; Coscia, A.; Griffante, G.; Galitska, G.; Pasquero, S.; Albano, C.; Biolatti, M. Where do we stand after decades of studying human cytomegalovirus? Microorganisms 2020, 8, 685. [Google Scholar] [CrossRef]
- Freeman, R., Jr. The ‘indirect’effects of cytomegalovirus infection. Am. J. Transplant. 2009, 9, 2453–2458. [Google Scholar] [CrossRef]
- Osman, N.M.; Sayed, N.M.; Abdel-Rahman, S.M.; Hamza, S.A. The impact of cytomegalovirus infection on mechanically ventilated patients in the respiratory and geriatric intensive care units. Egypt. J. Chest Dis. Tuberc. 2014, 63, 239–245. [Google Scholar] [CrossRef] [Green Version]
- Lanzieri, T.M.; Dollard, S.C.; Josephson, C.D.; Schmid, D.S.; Bialek, S.R. Breast milk–acquired cytomegalovirus infection and disease in VLBW and premature infants. Pediatrics 2013, 131, e1937–e1945. [Google Scholar] [CrossRef] [Green Version]
- Sivasai, K.; Mohanakumar, T.; Phelan, D.; Martin, S.; Anstey, M.; Brennan, D. Cytomegalovirus immune globulin intravenous (human) administration modulates immune response to alloantigens in sensitized renal transplant candidates. Clin. Exp. Immunol. 2000, 119, 559–565. [Google Scholar] [CrossRef]
- Santhanakrishnan, K.; Yonan, N.; Callan, P.; Karimi, E.; Al-Aloul, M.; Venkateswaran, R. The use of CMVIg rescue therapy in cardiothoracic transplantation: A single-center experience over 6 years (2011–2017). Clin. Transplant. 2019, 33, e13655. [Google Scholar] [CrossRef]
- Ishida, J.H.; Patel, A.; Mehta, A.K.; Gatault, P.; McBride, J.M.; Burgess, T.; Derby, M.A.; Snydman, D.R.; Emu, B.; Feierbach, B. Phase 2 randomized, double-blind, placebo-controlled trial of RG7667, a combination monoclonal antibody, for prevention of cytomegalovirus infection in high-risk kidney transplant recipients. Antimicrob. Agents Chemother. 2017, 61, e01794-16. [Google Scholar] [CrossRef] [Green Version]
- Erice, A.; Jordan, M.C.; Chace, B.A.; Fletcher, C.; Chinnock, B.J.; Balfour, H.H. Ganciclovir treatment of cytomegalovirus disease in transplant recipients and other immunocompromised hosts. JAMA 1987, 257, 3082–3087. [Google Scholar] [CrossRef]
- Wolf, D.G.; Smith, I.L.; Lee, D.J.; Freeman, W.R.; Flores-Aguilar, M.; Spector, S.A. Mutations in human cytomegalovirus UL97 gene confer clinical resistance to ganciclovir and can be detected directly in patient plasma. J. Clin. Investig. 1995, 95, 257–263. [Google Scholar] [CrossRef]
- Cheeran, M.C.-J.; Lokensgard, J.R.; Schleiss, M.R. Neuropathogenesis of congenital cytomegalovirus infection: Disease mechanisms and prospects for intervention. Clin. Microbiol. Rev. 2009, 22, 99–126. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Qiu, A.; Wang, J.; Pan, Z. Comparative Effects of Valganciclovir and Ganciclovir on the Congenital Cytomegalovirus Infection and Hearing Loss: A Randomized Controlled Trial. Iran. J. Pediatr. 2022, 32, e118874. [Google Scholar] [CrossRef]
- Kimberlin, D.W.; Jester, P.M.; Sánchez, P.J.; Ahmed, A.; Arav-Boger, R.; Michaels, M.G.; Ashouri, N.; Englund, J.A.; Estrada, B.; Jacobs, R.F. Valganciclovir for symptomatic congenital cytomegalovirus disease. N. Engl. J. Med. 2015, 372, 933–943. [Google Scholar] [CrossRef] [Green Version]
- Oliver, S.E.; Cloud, G.A.; Sánchez, P.J.; Demmler, G.J.; Dankner, W.; Shelton, M.; Jacobs, R.F.; Vaudry, W.; Pass, R.F.; Soong, S.-J. Neurodevelopmental outcomes following ganciclovir therapy in symptomatic congenital cytomegalovirus infections involving the central nervous system. J. Clin. Virol. 2009, 46, S22–S26. [Google Scholar] [CrossRef] [Green Version]
- McGregor, A.; Choi, K.Y. Cytomegalovirus antivirals and development of improved animal models. Expert Opin. Drug Metab. Toxicol. 2011, 7, 1245–1265. [Google Scholar] [CrossRef] [Green Version]
- Razonable, R.R.; Humar, A. Cytomegalovirus in solid organ transplant recipients—Guidelines of the American Society of Transplantation Infectious Diseases Community of Practice. Clin. Transplant. 2019, 33. [Google Scholar] [CrossRef]
- Martin, D.F.; Sierra-Madero, J.; Walmsley, S.; Wolitz, R.A.; Macey, K.; Georgiou, P.; Robinson, C.A.; Stempien, M.J. A controlled trial of valganciclovir as induction therapy for cytomegalovirus retinitis. N. Engl. J. Med. 2002, 346, 1119–1126. [Google Scholar] [CrossRef]
- Lea, A.P.; Bryson, H.M. Cidofovir. Drugs 1996, 52, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Safrin, S.; Cherrington, J.; Jaffe, H.S. Cidofovir. Antivir. Chemother. 1999, 5, 111–120. [Google Scholar]
- Upadhyayula, S.; Michaels, M.G. Ganciclovir, foscarnet, and cidofovir: Antiviral drugs not just for cytomegalovirus. J. Pediatr. Infect. Dis. Soc. 2013, 2, 286–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Z.; Xu, N.; Jiang, L.; Shi, P.; Fan, Z.; Huang, F.; Liu, Q.; Sun, J.; Yu, G. Cidofovir, a Choice for Salvage Treatment of CMV Infection in the Patients with Haploidentical Hematopoitic Stem Cell Transplantation. Blood 2020, 136, 40–41. [Google Scholar] [CrossRef]
- Caruso Brown, A.E.; Cohen, M.N.; Tong, S.; Braverman, R.S.; Rooney, J.F.; Giller, R.; Levin, M.J. Pharmacokinetics and safety of intravenous cidofovir for life-threatening viral infections in pediatric hematopoietic stem cell transplant recipients. Antimicrob. Agents Chemother. 2015, 59, 3718–3725. [Google Scholar] [CrossRef] [Green Version]
- US Food & Drug Administration. Orphan Drug Designations and Approvals. Available online: https://www.accessdata.fda.gov/scripts/opdlisting/oopd/detailedIndex.cfm?cfgridkey=340211 (accessed on 9 April 2023).
- Ligat, G.; Cazal, R.; Hantz, S.; Alain, S. The human cytomegalovirus terminase complex as an antiviral target: A close-up view. FEMS Microbiol. Rev. 2018, 42, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Aryal, S.; Katugaha, S.B.; Cochrane, A.; Brown, A.W.; Nathan, S.D.; Shlobin, O.A.; Ahmad, K.; Marinak, L.; Chun, J.; Fregoso, M. Single-center experience with use of letermovir for CMV prophylaxis or treatment in thoracic organ transplant recipients. Transpl. Infect. Dis. 2019, 21, e13166. [Google Scholar] [CrossRef]
- Vyas, A.; Raval, A.D.; Kamat, S.; LaPlante, K.; Tang, Y.; Chemaly, R.F. In Real-World Outcomes Associated with Letermovir Use for Cytomegalovirus Primary Prophylaxis in Allogeneic-Hematopoietic Cell Transplant Recipients: A Systematic Review and Meta-Analysis of Observational Studies. Open Forum Infect. Dis. 2023, 10, ofac687. [Google Scholar] [CrossRef]
- Wolf, D.G.; Courcelle, C.T.; Prichard, M.N.; Mocarski, E.S. Distinct and separate roles for herpesvirus-conserved UL97 kinase in cytomegalovirus DNA synthesis and encapsidation. Proc. Natl. Acad. Sci. USA 2001, 98, 1895–1900. [Google Scholar] [CrossRef] [Green Version]
- Avery, R.K.; Alain, S.; Alexander, B.D.; Blumberg, E.A.; Chemaly, R.F.; Cordonnier, C.; Duarte, R.F.; Florescu, D.F.; Kamar, N.; Kumar, D. Maribavir for refractory cytomegalovirus infections with or without resistance post-transplant: Results from a phase 3 randomized clinical trial. Clin. Infect. Dis. 2022, 75, 690–701. [Google Scholar] [CrossRef]
- Kang, C. Maribavir: First approval. Drugs 2022, 82, 335–340. [Google Scholar] [CrossRef]
- Ornaghi, S.; Davis, J.N.; Gorres, K.L.; Miller, G.; Paidas, M.J.; van den Pol, A.N. Mood stabilizers inhibit cytomegalovirus infection. Virology 2016, 499, 121–135. [Google Scholar] [CrossRef]
- John, G.T.; Manivannan, J.; Chandy, S.; Peter, S.; Jacob, C.K. Leflunomide therapy for cytomegalovirus disease in renal allograft recepients. Transplantation 2004, 77, 1460–1461. [Google Scholar] [CrossRef]
- Gokarn, A.; Toshniwal, A.; Pathak, A.; Arora, S.; Bonda, A.; Punatar, S.; Nayak, L.; Dwivedi, P.; Bhat, V.; Biswas, S. Use of leflunomide for treatment of cytomegalovirus infection in recipients of allogeneic stem cell transplant. Biol. Blood Marrow Transplant. 2019, 25, 1832–1836. [Google Scholar] [CrossRef]
- Garofoli, F.; Lombardi, G.; Angelini, M.; Campanini, G.; Zavattoni, M.; Baldanti, F. Onset of valganciclovir resistance in two infants with congenital cytomegalovirus infection. Int. J. Infect. Dis. 2020, 98, 150–152. [Google Scholar] [CrossRef]
- Chou, S. Advances in the genotypic diagnosis of cytomegalovirus antiviral drug resistance. Antivir. Res. 2020, 176, 104711. [Google Scholar] [CrossRef]
- Torii, Y.; Horiba, K.; Kawada, J.-i.; Haruta, K.; Yamaguchi, M.; Suzuki, T.; Uryu, H.; Kashiwa, N.; Goishi, K.; Ogi, T. Detection of antiviral drug resistance in patients with congenital cytomegalovirus infection using long-read sequencing: A retrospective observational study. BMC Infect. Dis. 2022, 22, 568. [Google Scholar] [CrossRef]
- Plotkin, S.; Furukawa, T.; Zygraich, N.; Huygelen, C. Candidate cytomegalovirus strain for human vaccination. Infect. Immun. 1975, 12, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Elek, S.; Stern, H. Development of a vaccine against mental retardation caused by cytomegalovirus infection in utero. Lancet 1974, 303, 1–5. [Google Scholar] [CrossRef]
- Plotkin, S.A.; Boppana, S.B. Vaccination against the human cytomegalovirus. Vaccine 2019, 37, 7437–7442. [Google Scholar] [CrossRef]
- Scarpini, S.; Morigi, F.; Betti, L.; Dondi, A.; Biagi, C.; Lanari, M. Development of a vaccine against human cytomegalovirus: Advances, barriers, and implications for the clinical practice. Vaccines 2021, 9, 551. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.S.; Herold, B.C.; Permar, S.R. A new era in cytomegalovirus vaccinology: Considerations for rational design of next-generation vaccines to prevent congenital cytomegalovirus infection. NPJ Vaccines 2018, 3, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipowicz, W.; Jaskiewicz, L.; Kolb, F.A.; Pillai, R.S. Post-transcriptional gene silencing by siRNAs and miRNAs. Curr. Opin. Struct. Biol. 2005, 15, 331–341. [Google Scholar] [CrossRef]
- Lam, J.K.; Chow, M.Y.; Zhang, Y.; Leung, S.W. siRNA versus miRNA as therapeutics for gene silencing. Mol. Ther. Nucleic Acids 2015, 4, e252. [Google Scholar] [CrossRef] [Green Version]
- Xiaofei, E.; Stadler, B.M.; Debatis, M.; Wang, S.; Lu, S.; Kowalik, T.F. RNA interference-mediated targeting of human cytomegalovirus immediate-early or early gene products inhibits viral replication with differential effects on cellular functions. J. Virol. 2012, 86, 5660. [Google Scholar]
- Møller, R.; Schwarz, T.M.; Noriega, V.M.; Panis, M.; Sachs, D.; Tortorella, D.; tenOever, B.R. miRNA-mediated targeting of human cytomegalovirus reveals biological host and viral targets of IE2. Proc. Natl. Acad. Sci. USA 2018, 115, 1069–1074. [Google Scholar] [CrossRef] [Green Version]
- Dönmüş, B.; Ünal, S.; Kirmizitaş, F.C.; Türkoğlu Laçin, N. Virus-associated ribozymes and nano carriers against COVID-19. Artif. Cells Nanomed. Biotechnol. 2021, 49, 204–218. [Google Scholar] [CrossRef]
- Scherr, M.; Grez, M.; Ganser, A.; Engels, J.W. Specific hammerhead ribozyme-mediated cleavage of mutant N-ras mRNA in vitro and ex vivo: Oligoribonucleotides as therapeutic agents. J. Biol. Chem. 1997, 272, 14304–14313. [Google Scholar] [CrossRef] [Green Version]
- Panda, K.; Alagarasu, K.; Parashar, D. Oligonucleotide-based approaches to inhibit dengue virus replication. Molecules 2021, 26, 956. [Google Scholar] [CrossRef]
- Walker, S.C.; Engelke, D.R. Ribonuclease P: The evolution of an ancient RNA enzyme. Crit. Rev. Biochem. Mol. Biol. 2006, 41, 77–102. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Shu, R.; Liu, J. The development and improvement of ribonucleic acid therapy strategies. Mol. Ther. Nucleic Acids 2021, 26, 997–1013. [Google Scholar] [CrossRef]
- Trang, P.; Lee, M.; Nepomuceno, E.; Kim, J.; Zhu, H.; Liu, F. Effective inhibition of human cytomegalovirus gene expression and replication by a ribozyme derived from the catalytic RNA subunit of RNase P from Escherichia coli. Proc. Natl. Acad. Sci. USA 2000, 97, 5812–5817. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Trang, P.; Umamoto, S.; Hai, R.; Liu, F. RNase P ribozyme inhibits cytomegalovirus replication by blocking the expression of viral capsid proteins. Nucleic Acids Res. 2004, 32, 3427–3434. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Vu, G.-P.; Qian, H.; Chen, Y.-C.; Wang, Y.; Reeves, M.; Zen, K.; Liu, F. Engineered RNase P ribozymes effectively inhibit human cytomegalovirus gene expression and replication. Viruses 2014, 6, 2376–2391. [Google Scholar] [CrossRef] [Green Version]
- Deng, Q.; Liu, Y.; Li, X.; Yan, B.; Sun, X.; Tang, W.; Trang, P.; Yang, Z.; Gong, H.; Wang, Y. Inhibition of human cytomegalovirus major capsid protein expression and replication by ribonuclease P–associated external guide sequences. RNA 2019, 25, 645–655. [Google Scholar] [CrossRef]
- Tripathi, S.; Khatri, P.; Fatima, Z.; Pandey, R.P.; Hameed, S. A Landscape of CRISPR/Cas Technique for Emerging Viral Disease Diagnostics and Therapeutics: Progress and Prospects. Pathogens 2023, 12, 56. [Google Scholar] [CrossRef]
- Asmamaw, M.; Zawdie, B. Mechanism and applications of CRISPR/Cas-9-mediated genome editing. Biol. Targets Ther. 2021, 15, 353–361. [Google Scholar]
- Gergen, J.; Coulon, F.; Creneguy, A.; Elain-Duret, N.; Gutierrez, A.; Pinkenburg, O.; Verhoeyen, E.; Anegon, I.; Nguyen, T.H.; Halary, F.A. Multiplex CRISPR/Cas9 system impairs HCMV replication by excising an essential viral gene. PLoS ONE 2018, 13, e0192602. [Google Scholar] [CrossRef] [Green Version]
- van Diemen, F.R.; Kruse, E.M.; Hooykaas, M.J.; Bruggeling, C.E.; Schürch, A.C.; van Ham, P.M.; Imhof, S.M.; Nijhuis, M.; Wiertz, E.J.; Lebbink, R.J. CRISPR/Cas9-mediated genome editing of herpesviruses limits productive and latent infections. PLoS Pathog. 2016, 12, e1005701. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Deng, J.; Zhang, Q.; Ma, P.; Lv, L.; Zhang, Y.; Li, C.; Zhang, Y. Targeting human cytomegalovirus IE genes by CRISPR/Cas9 nuclease effectively inhibits viral replication and reactivation. Arch. Virol. 2020, 165, 1827–1835. [Google Scholar] [CrossRef] [PubMed]
- Bloom, K.; Mussolino, C.; Arbuthnot, P. Transcription activator-like effector (TALE) nucleases and repressor TALEs for antiviral gene therapy. Curr. Stem Cell Rep. 2015, 1, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-J.; Chen, Y.-C. Potential application of TALENs against murine cytomegalovirus latent infections. Viruses 2019, 11, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ku, T.-H.; Zhang, T.; Luo, H.; Yen, T.M.; Chen, P.-W.; Han, Y.; Lo, Y.-H. Nucleic acid aptamers: An emerging tool for biotechnology and biomedical sensing. Sensors 2015, 15, 16281–16313. [Google Scholar] [CrossRef] [Green Version]
- Gelinas, A.D.; Tan, T.K.; Liu, S.; Jaramillo, J.G.; Chadwick, J.; Harding, A.C.; Zhang, C.; Ream, B.E.; Chase, C.N.; Otis, M.R. Broadly neutralizing aptamers to SARS-CoV-2: A diverse panel of modified DNA antiviral agents. Mol. Ther. Nucleic Acids 2023, 31, 370–382. [Google Scholar] [CrossRef]
- Jeon, S.H.; Kayhan, B.; Ben-Yedidia, T.; Arnon, R. A DNA aptamer prevents influenza infection by blocking the receptor binding region of the viral hemagglutinin. J. Biol. Chem. 2004, 279, 48410–48419. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, N.; Lischka, P.; Wagenknecht, N.; Stamminger, T. Inhibition of human cytomegalovirus replication via peptide aptamers directed against the nonconventional nuclear localization signal of the essential viral replication factor pUL84. J. Virol. 2009, 83, 11902–11913. [Google Scholar] [CrossRef] [Green Version]
- Group, V.S. A randomized controlled clinical trial of intravitreous fomivirsen for treatment of newly diagnosed peripheral cytomegalovirus retinitis in patients with AIDS. Am. J. Ophthalmol. 2002, 133, 467–474. [Google Scholar]
- Bhat, G.R.; Sethi, I.; Rah, B.; Kumar, R.; Afroze, D. Innovative in Silico Approaches for Characterization of Genes and Proteins. Front. Genet. 2022, 13, 865182. [Google Scholar] [CrossRef]
- Hagedorn, P.H.; Hansen, B.R.; Koch, T.; Lindow, M. Managing the sequence-specificity of antisense oligonucleotides in drug discovery. Nucleic Acids Res. 2017, 45, 2262–2282. [Google Scholar] [CrossRef] [Green Version]
- Maus, A.; Strait, L.; Zhu, D. Nanoparticles as delivery vehicles for antiviral therapeutic drugs. Eng. Regen. 2021, 2, 31–46. [Google Scholar] [CrossRef]
- Jarach, N.; Dodiuk, H.; Kenig, S. Polymers in the medical antiviral front-line. Polymers 2020, 12, 1727. [Google Scholar] [CrossRef]
- Hawner, M.; Ducho, C. Cellular targeting of oligonucleotides by conjugation with small molecules. Molecules 2020, 25, 5963. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Generic Name | Mechanism of Action | Route of Administration | Major Toxicity |
|---|---|---|---|
| Ganciclovir and Valganciclovir | Inhibition of viral DNA replication by targeting DNA polymerase | Intravenous, Oral | Bone marrow suppression, encephalopathy, carcinogenicity and possibly hepatotoxicity and reduced fertility |
| Foscarnet | Pyrophosphate analog/Inhibition of viral DNA replication by targeting DNA polymerase | Intravenous | Nephrotoxicity, hypocalcemia, electrolytes imbalance, genital ulceration |
| Cidofivir | Nucleotide analog/Inhibition of viral DNA replication by targeting DNA polymerase | Intravenous | Nephrotoxicity |
| Letermovir | Binds to components of the terminase complex (UL56/UL89) | Intravenous, Oral | Bone marrow suppression and nephrotoxicity |
| Maribavir | Inhibition of CMV UL97 gene product | Oral | Taste disturbance and gastrointestinal |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panda, K.; Parashar, D.; Viswanathan, R. An Update on Current Antiviral Strategies to Combat Human Cytomegalovirus Infection. Viruses 2023, 15, 1358. https://doi.org/10.3390/v15061358
Panda K, Parashar D, Viswanathan R. An Update on Current Antiviral Strategies to Combat Human Cytomegalovirus Infection. Viruses. 2023; 15(6):1358. https://doi.org/10.3390/v15061358
Chicago/Turabian StylePanda, Kingshuk, Deepti Parashar, and Rajlakshmi Viswanathan. 2023. "An Update on Current Antiviral Strategies to Combat Human Cytomegalovirus Infection" Viruses 15, no. 6: 1358. https://doi.org/10.3390/v15061358