
The Tumor-Specific Immune Landscape in HPV+ Head and Neck Cancer

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. HPV+ HNSCC: A Distinct Form of HNSCC and HPV-Associated Cancer
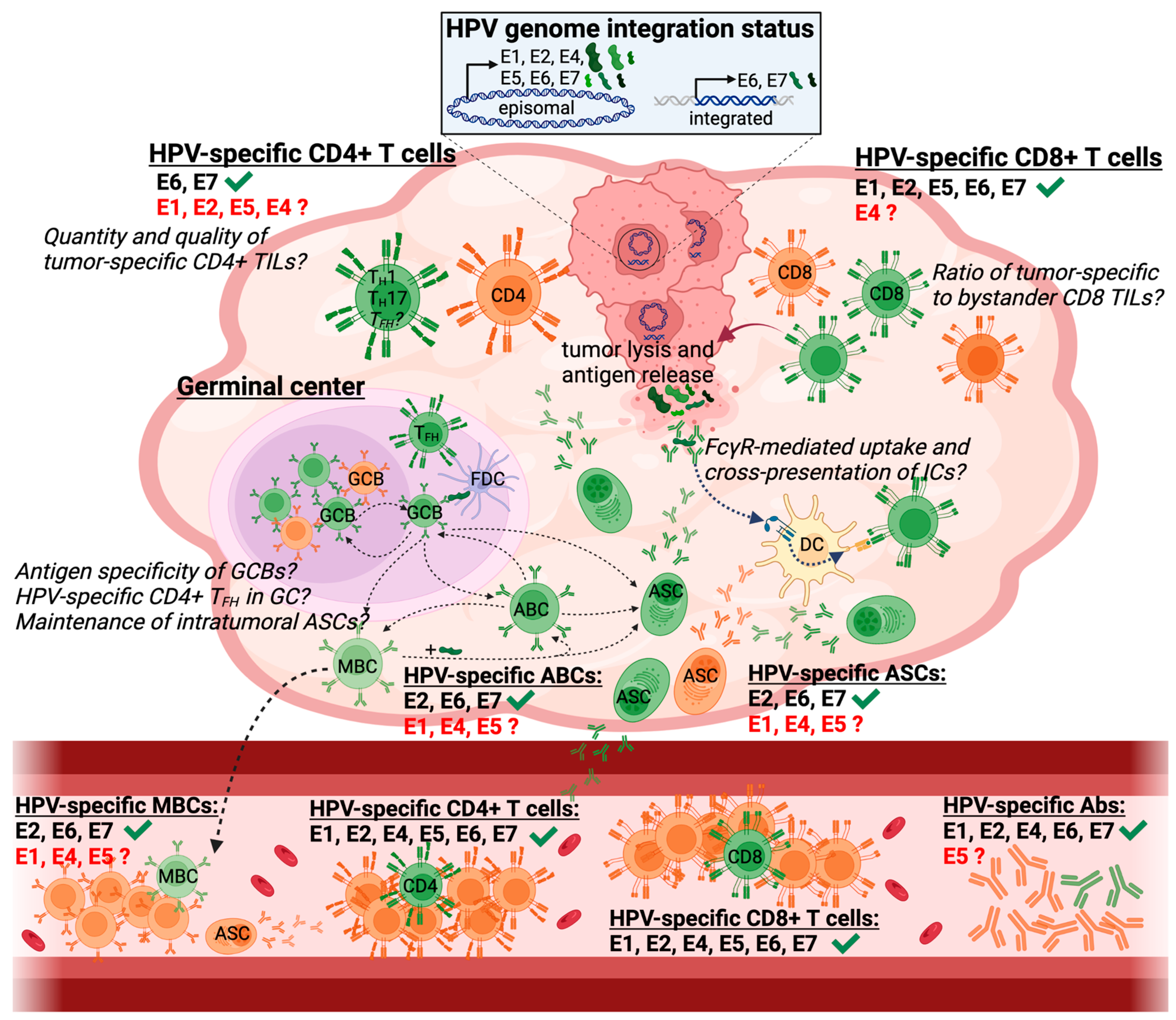
3. HPV-Specific Immune Responses in HPV+ HNSCC
3.1. HPV-Specific Immune Responses in the Peripheral Blood
3.1.1. HPV-Specific T Cell Responses
3.1.2. HPV-Specific Humoral Responses
3.2. Intratumoral Immune Responses in HPV+ HNSCC
3.2.1. Intratumoral Lymphoid Structures
3.2.2. HPV-Specific TIL Responses
3.2.3. HPV-Specific Intratumoral B Cell Responses
4. Implications for Immunotherapy
4.1. Immune Checkpoint Blockade
4.2. Therapeutic Vaccines
4.3. Adoptive Cell Therapies
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chaturvedi, A.K.; Engels, E.A.; Pfeiffer, R.M.; Hernandez, B.Y.; Xiao, W.; Kim, E.; Jiang, B.; Goodman, M.T.; Sibug-Saber, M.; Cozen, W.; et al. Human Papillomavirus and Rising Oropharyngeal Cancer Incidence in the United States. J. Clin. Oncol. 2011, 29, 4294–4301. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.P.; Saha, S.; Yu, M.; Kimple, R.J.; Lambert, P.F. Prevalence of Human Papillomavirus in Oropharyngeal Squamous Cell Carcinoma in the United States Across Time. Chem. Res. Toxicol. 2014, 27, 462–469. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer Statistics, 2023. CA A Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef]
- Scott-Wittenborn, N.; D’Souza, G.; Tewari, S.; Rooper, L.; Troy, T.; Drake, V.; Bigelow, E.O.; Windon, M.J.; Ryan, W.R.; Ha, P.K.; et al. Prevalence of Human Papillomavirus in Head and Neck Cancers at Tertiary Care Centers in the United States over Time. Cancer 2022, 128, 1767–1774. [Google Scholar] [CrossRef] [PubMed]
- Tota, J.E.; Best, A.F.; Zumsteg, Z.S.; Gillison, M.L.; Rosenberg, P.S.; Chaturvedi, A.K. Evolution of the Oropharynx Cancer Epidemic in the United States: Moderation of Increasing Incidence in Younger Individuals and Shift in the Burden to Older Individuals. J. Clin. Oncol. 2019, 37, 1538–1546. [Google Scholar] [CrossRef] [PubMed]
- How Many Cancers Are Linked with HPV Each Year?|CDC. Available online: https://www.cdc.gov/cancer/hpv/statistics/cases.htm (accessed on 3 February 2023).
- Liao, C.-I.; Francoeur, A.A.; Kapp, D.S.; Caesar, M.A.P.; Huh, W.K.; Chan, J.K. Trends in Human Papillomavirus–Associated Cancers, Demographic Characteristics, and Vaccinations in the US, 2001–2017. JAMA Netw. Open 2022, 5, e222530. [Google Scholar] [CrossRef]
- Hildesheim, A.; Schiffman, M.; Bromley, C.; Wacholder, S.; Herrero, R.; Rodriguez, A.C.; Bratti, M.C.; Sherman, M.E.; Scarpidis, U.; Lin, Q.-Q.; et al. Human Papillomavirus Type 16 Variants and Risk of Cervical Cancer. JNCI J. Natl. Cancer Inst. 2001, 93, 315–318. [Google Scholar] [CrossRef]
- Ahmed, H.G.; Bensumaidea, S.H.; Alshammari, F.D.; Alenazi, F.S.H.; ALmutlaq, B.A.; Alturkstani, M.Z.; Aladani, I.A. Prevalence of Human Papillomavirus Subtypes 16 and 18 among Yemeni Patients with Cervical Cancer. Asian Pac. J. Cancer Prev. 2017, 18, 1543–1548. [Google Scholar] [CrossRef]
- Begum, S.; Cao, D.; Gillison, M.; Zahurak, M.; Westra, W.H. Tissue Distribution of Human Papillomavirus 16 DNA Integration in Patients with Tonsillar Carcinoma. Clin. Cancer Res. 2005, 11, 5694–5699. [Google Scholar] [CrossRef]
- Wieland, A.; Patel, M.R.; Cardenas, M.A.; Eberhardt, C.S.; Hudson, W.H.; Obeng, R.C.; Griffith, C.C.; Wang, X.; Chen, Z.G.; Kissick, H.T.; et al. Defining HPV-Specific B Cell Responses in Patients with Head and Neck Cancer. Nature 2021, 597, 274–278. [Google Scholar] [CrossRef]
- Mehanna, H.; Beech, T.; Nicholson, T.; El-Hariry, I.; McConkey, C.; Paleri, V.; Roberts, S. Prevalence of Human Papillomavirus in Oropharyngeal and Nonoropharyngeal Head and Neck Cancer—Systematic Review and Meta-Analysis of Trends by Time and Region. Head Neck 2013, 35, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Bratman, S.V.; Bruce, J.P.; O’Sullivan, B.; Pugh, T.J.; Xu, W.; Yip, K.W.; Liu, F.-F. Human Papillomavirus Genotype Association with Survival in Head and Neck Squamous Cell Carcinoma. JAMA Oncol. 2016, 2, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Mazul, A.L.; Rodriguez-Ormaza, N.; Taylor, J.M.; Desai, D.D.; Brennan, P.; Anantharaman, D.; Gheit, T.; Tommasino, M.; Abedi-Ardekani, B.; Olshan, A.F.; et al. Prognostic Significance of Non-HPV16 Genotypes in Oropharyngeal Squamous Cell Carcinoma. Oral Oncol. 2016, 61, 98–103. [Google Scholar] [CrossRef]
- Kreimer, A.R.; Clifford, G.M.; Boyle, P.; Franceschi, S. Human Papillomavirus Types in Head and Neck Squamous Cell Carcinomas Worldwide: A Systematic Review. Cancer Epidemiol. Biomark. Prev. 2005, 14, 467–475. [Google Scholar] [CrossRef]
- Nulton, T.J.; Olex, A.L.; Dozmorov, M.; Morgan, I.M.; Windle, B. Analysis of The Cancer Genome Atlas Sequencing Data Reveals Novel Properties of the Human Papillomavirus 16 Genome in Head and Neck Squamous Cell Carcinoma. Oncotarget 2017, 8, 17684–17699. [Google Scholar] [CrossRef]
- Li, N.; Franceschi, S.; Howell-Jones, R.; Snijders, P.J.F.; Clifford, G.M. Human Papillomavirus Type Distribution in 30,848 Invasive Cervical Cancers Worldwide: Variation by Geographical Region, Histological Type and Year of Publication. Int. J. Cancer 2011, 128, 927–935. [Google Scholar] [CrossRef]
- Cancer Registration Statistics, England—Office for National Statistics. Available online: https://www.ons.gov.uk/peoplepopulationandcommunity/healthandsocialcare/conditionsanddiseases/bulletins/cancerregistrationstatisticsengland/2016 (accessed on 1 March 2023).
- Schache, A.G.; Powell, N.G.; Cuschieri, K.S.; Robinson, M.; Leary, S.; Mehanna, H.; Rapozo, D.; Long, A.; Cubie, H.; Junor, E.; et al. HPV-Related Oropharynx Cancer in the United Kingdom: An Evolution in the Understanding of Disease Etiology. Cancer Res. 2016, 76, 6598–6606. [Google Scholar] [CrossRef]
- Xu, T.; Shen, C.; Wei, Y.; Hu, C.; Wang, Y.; Xiang, J.; Sun, G.; Su, F.; Wang, Q.; Lu, X. Human Papillomavirus (HPV) in Chinese Oropharyngeal Squamous Cell Carcinoma (OPSCC): A Strong Predilection for the Tonsil. Cancer Med. 2020, 9, 6556–6564. [Google Scholar] [CrossRef] [PubMed]
- Oga, E.A.; Schumaker, L.M.; Alabi, B.S.; Obaseki, D.; Umana, A.; Bassey, I.-A.; Ebughe, G.; Oluwole, O.; Akeredolu, T.; Adebamowo, S.N.; et al. Paucity of HPV-Related Head and Neck Cancers (HNC) in Nigeria. PLoS ONE 2016, 11, e0152828. [Google Scholar] [CrossRef]
- de Martel, C.; Plummer, M.; Vignat, J.; Franceschi, S. Worldwide Burden of Cancer Attributable to HPV by Site, Country and HPV Type. Int. J. Cancer 2017, 141, 664–670. [Google Scholar] [CrossRef]
- Kirnbauer, R.; Booy, F.; Cheng, N.; Lowy, D.R.; Schiller, J.T. Papillomavirus L1 Major Capsid Protein Self-Assembles into Virus-like Particles That Are Highly Immunogenic. Proc. Natl. Acad. Sci. USA 1992, 89, 12180–12184. [Google Scholar] [CrossRef]
- Siddiqui, M.A.A.; Perry, C.M. Human Papillomavirus Quadrivalent (Types 6, 11, 16, 18) Recombinant Vaccine (Gardasil). Drugs 2006, 66, 1263–1271; discussion 1272–1273. [Google Scholar] [CrossRef]
- Harper, D.M.; Franco, E.L.; Wheeler, C.; Ferris, D.G.; Jenkins, D.; Schuind, A.; Zahaf, T.; Innis, B.; Naud, P.; De Carvalho, N.S.; et al. Efficacy of a Bivalent L1 Virus-like Particle Vaccine in Prevention of Infection with Human Papillomavirus Types 16 and 18 in Young Women: A Randomised Controlled Trial. Lancet 2004, 364, 1757–1765. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, T.; Spaczynski, M.; Kaufmann, A.; Wysocki, J.; Gałaj, A.; Schulze, K.; Suryakiran, P.; Thomas, F.; Descamps, D. Persistence of Immune Responses to the HPV-16/18 AS04-Adjuvanted Vaccine in Women Aged 15-55 Years and First-Time Modelling of Antibody Responses in Mature Women: Results from an Open-Label 6-Year Follow-up Study. BJOG Int. J. Obstet. Gynaecol. 2015, 122, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Anantharaman, D.; Muwonge, R.; Bhatla, N.; Panicker, G.; Butt, J.; Rani Reddy Poli, U.; Malvi, S.G.; Esmy, P.O.; Lucas, E.; et al. Evaluation of Immune Response to Single Dose of Quadrivalent HPV Vaccine at 10-Year Post-Vaccination. Vaccine 2023, 41, 236–245. [Google Scholar] [CrossRef]
- Pasmans, H.; Schurink-Van’t Klooster, T.M.; Bogaard, M.J.M.; van Rooijen, D.M.; de Melker, H.E.; Welters, M.J.P.; van der Burg, S.H.; van der Klis, F.R.M.; Buisman, A.-M. Long-Term HPV-Specific Immune Response after One versus Two and Three Doses of Bivalent HPV Vaccination in Dutch Girls. Vaccine 2019, 37, 7280–7288. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.A.; Dillner, J.; Beddows, S.; Unger, E.R. Immunogenicity of HPV Prophylactic Vaccines: Serology Assays and Their Use in HPV Vaccine Evaluation and Development. Vaccine 2018, 36, 4792–4799. [Google Scholar] [CrossRef]
- Scherpenisse, M.; Mollers, M.; Schepp, R.M.; Meijer, C.J.L.M.; de Melker, H.E.; Berbers, G.A.M.; van der Klis, F.R.M. Detection of Systemic and Mucosal HPV-Specific IgG and IgA Antibodies in Adolescent Girls One and Two Years after HPV Vaccination. Hum. Vaccines Immunother. 2013, 9, 314–321. [Google Scholar] [CrossRef]
- Costa, A.P.F.; Gonçalves, A.K.; Machado, P.R.L.; de Souza, L.B.F.C.; Sarmento, A.; Cobucci, R.N.O.; Giraldo, P.C.; Witkin, S.S. Immune Response to Human Papillomavirus One Year after Prophylactic Vaccination with AS04-Adjuvanted HPV-16/18 Vaccine: HPV-Specific IgG and IgA Antibodies in the Circulation and the Cervix. Asian Pac. J. Cancer Prev. 2018, 19, 2313–2317. [Google Scholar] [CrossRef]
- Huo, Z.; Bissett, S.L.; Giemza, R.; Beddows, S.; Oeser, C.; Lewis, D.J.M. Systemic and Mucosal Immune Responses to Sublingual or Intramuscular Human Papilloma Virus Antigens in Healthy Female Volunteers. PLoS ONE 2012, 7, e33736. [Google Scholar] [CrossRef]
- Mariz, F.C.; Bender, N.; Anantharaman, D.; Basu, P.; Bhatla, N.; Pillai, M.R.; Prabhu, P.R.; Sankaranarayanan, R.; Eriksson, T.; Pawlita, M.; et al. Peak Neutralizing and Cross-Neutralizing Antibody Levels to Human Papillomavirus Types 6/16/18/31/33/45/52/58 Induced by Bivalent and Quadrivalent HPV Vaccines. Npj Vaccines 2020, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Mariz, F.C.; Gray, P.; Bender, N.; Eriksson, T.; Kann, H.; Apter, D.; Paavonen, J.; Pajunen, E.; Prager, K.M.; Sehr, P.; et al. Sustainability of Neutralising Antibodies Induced by Bivalent or Quadrivalent HPV Vaccines and Correlation with Efficacy: A Combined Follow-up Analysis of Data from Two Randomised, Double-Blind, Multicentre, Phase 3 Trials. Lancet Infect. Dis. 2021, 21, 1458–1468. [Google Scholar] [CrossRef]
- Costa, A.P.; Giraldo, P.C.; Cobucci, R.N.; Consolaro, M.L.; Souza, R.P.; Canário, L.B.; Machado, P.R.; Randall Martins, R.; Vieira Baptista, P.; Eleutério, J., Jr.; et al. Cross-Protective IgG and IgA Antibodies against Oncogenic and Non-Oncogenic HPV Genotypes. Asian Pac. J. Cancer Prev. 2020, 21, 2799–2804. [Google Scholar] [CrossRef]
- Scherpenisse, M.; Schepp, R.M.; Mollers, M.; Meijer, C.J.L.M.; Berbers, G.A.M.; van der Klis, F.R.M. Characteristics of HPV-Specific Antibody Responses Induced by Infection and Vaccination: Cross-Reactivity, Neutralizing Activity, Avidity and IgG Subclasses. PLoS ONE 2013, 8, e74797. [Google Scholar] [CrossRef] [PubMed]
- Isho, B.; Florescu, A.; Wang, A.A.; Gommerman, J.L. Fantastic IgA Plasma Cells and Where to Find Them. Immunol. Rev. 2021, 303, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.A.; Kemp, T.J.; Torres, B.N.; Isaacs-Soriano, K.; Ingles, D.; Abrahamsen, M.; Pan, Y.; Lazcano-Ponce, E.; Salmeron, J.; Giuliano, A.R. Quadrivalent Human Papillomavirus (HPV) Vaccine Induces HPV-Specific Antibodies in the Oral Cavity: Results from the Mid-Adult Male Vaccine Trial. J. Infect. Dis. 2016, 214, 1276–1283. [Google Scholar] [CrossRef]
- Mehanna, H.; Bryant, T.S.; Babrah, J.; Louie, K.; Bryant, J.L.; Spruce, R.J.; Batis, N.; Olaleye, O.; Jones, J.; Struijk, L.; et al. Human Papillomavirus (HPV) Vaccine Effectiveness and Potential Herd Immunity for Reducing Oncogenic Oropharyngeal HPV-16 Prevalence in the United Kingdom: A Cross-Sectional Study. Clin. Infect. Dis. 2019, 69, 1296–1302. [Google Scholar] [CrossRef]
- Chaturvedi, A.K.; Graubard, B.I.; Broutian, T.; Pickard, R.K.L.; Tong, Z.-Y.; Xiao, W.; Kahle, L.; Gillison, M.L. Effect of Prophylactic Human Papillomavirus (HPV) Vaccination on Oral HPV Infections among Young Adults in the United States. J. Clin. Oncol. 2018, 36, 262–267. [Google Scholar] [CrossRef]
- Hirth, J.M.; Chang, M.; Resto, V.A.; Guo, F.; Berenson, A.B. Prevalence of Oral Human Papillomavirus by Vaccination Status among Young Adults (18–30years Old). Vaccine 2017, 35, 3446–3451. [Google Scholar] [CrossRef]
- Yang, D.X.; Soulos, P.R.; Davis, B.; Gross, C.P.; Yu, J.B. Impact of Widespread Cervical Cancer Screening: Number of Cancers Prevented and Changes in Race-Specific Incidence. Am. J. Clin. Oncol. 2018, 41, 289–294. [Google Scholar] [CrossRef]
- Lei, J.; Ploner, A.; Elfström, K.M.; Wang, J.; Roth, A.; Fang, F.; Sundström, K.; Dillner, J.; Sparén, P. HPV Vaccination and the Risk of Invasive Cervical Cancer. N. Engl. J. Med. 2020, 383, 1340–1348. [Google Scholar] [CrossRef]
- Viens, L.J. Human Papillomavirus–Associated Cancers—United States, 2008–2012. Morb. Mortal. Wkly. Rep. 2016, 65, 661–666. [Google Scholar] [CrossRef]
- Human Papillomavirus–Attributable Cancers—United States, 2012–2016|MMWR. Available online: https://www.cdc.gov/mmwr/volumes/68/wr/mm6833a3.htm?s_cid=mm6833a3_w (accessed on 3 April 2023).
- FDA Licensure of Quadrivalent Human Papillomavirus Vaccine (HPV4, Gardasil) for Use in Males and Guidance from the Advisory Committee on Immunization Practices (ACIP). Available online: https://www.cdc.gov/mmwr/preview/mmwrhtml/mm5920a5.htm (accessed on 30 March 2023).
- HPV Vaccine Age Limit: You Might Not Be Too Old—What You Should Know|Memorial Sloan Kettering Cancer Center. Available online: https://www.mskcc.org/news/think-you-re-too-old-get-hpv-vaccine-prevent-cancer-maybe-not (accessed on 9 March 2023).
- Pingali, C. National, Regional, State, and Selected Local Area Vaccination Coverage among Adolescents Aged 13–17 Years—United States, 2020. Morb. Mortal. Wkly. Rep. 2021, 70, 1183. [Google Scholar] [CrossRef]
- Kreimer, A.R.; Johansson, M.; Yanik, E.L.; Katki, H.A.; Check, D.P.; Lang Kuhs, K.A.; Willhauck-Fleckenstein, M.; Holzinger, D.; Hildesheim, A.; Pfeiffer, R.; et al. Kinetics of the Human Papillomavirus Type 16 E6 Antibody Response Prior to Oropharyngeal Cancer. J. Natl. Cancer Inst. 2017, 109, djx005. [Google Scholar] [CrossRef]
- Kreimer, A.R.; Ferreiro-Iglesias, A.; Nygard, M.; Bender, N.; Schroeder, L.; Hildesheim, A.; Robbins, H.A.; Pawlita, M.; Langseth, H.; Schlecht, N.F.; et al. Timing of HPV16-E6 Antibody Seroconversion before OPSCC: Findings from the HPVC3 Consortium. Ann. Oncol. 2019, 30, 1335–1343. [Google Scholar] [CrossRef]
- Leshchiner, I.; Mroz, E.A.; Cha, J.; Rosebrock, D.; Spiro, O.; Bonilla-Velez, J.; Faquin, W.C.; Lefranc-Torres, A.; Lin, D.T.; Michaud, W.A.; et al. Inferring Early Genetic Progression in Cancers with Unobtainable Premalignant Disease. Nat. Cancer 2023, 4, 550–563. [Google Scholar] [CrossRef]
- Gillison, M.L.; Chaturvedi, A.K.; Anderson, W.F.; Fakhry, C. Epidemiology of Human Papillomavirus–Positive Head and Neck Squamous Cell Carcinoma. J. Clin. Oncol. 2015, 33, 3235–3242. [Google Scholar] [CrossRef]
- Ang, K.K.; Harris, J.; Wheeler, R.; Weber, R.; Rosenthal, D.I.; Nguyen-Tân, P.F.; Westra, W.H.; Chung, C.H.; Jordan, R.C.; Lu, C.; et al. Human Papillomavirus and Survival of Patients with Oropharyngeal Cancer. N. Engl. J. Med. 2010, 363, 24–35. [Google Scholar] [CrossRef]
- Amin, M.B.; Greene, F.L.; Edge, S.B.; Compton, C.C.; Gershenwald, J.E.; Brookland, R.K.; Meyer, L.; Gress, D.M.; Byrd, D.R.; Winchester, D.P. The Eighth Edition AJCC Cancer Staging Manual: Continuing to Build a Bridge from a Population-Based to a More “Personalized” Approach to Cancer Staging. CA Cancer J. Clin. 2017, 67, 93–99. [Google Scholar] [CrossRef]
- Windon, M.J.; D’Souza, G.; Rettig, E.M.; Westra, W.H.; van Zante, A.; Wang, S.J.; Ryan, W.R.; Mydlarz, W.K.; Ha, P.K.; Miles, B.A.; et al. Increasing Prevalence of HPV-Positive Oropharyngeal Cancers among Older Adults. Cancer 2018, 124, 2993–2999. [Google Scholar] [CrossRef]
- Cline, B.J.; Simpson, M.C.; Gropler, M.; Bukatko, A.R.; Adjei Boakye, E.; Mohammed, K.A.; Osazuwa-Peters, N. Change in Age at Diagnosis of Oropharyngeal Cancer in the United States, 1975–2016. Cancers 2020, 12, 3191. [Google Scholar] [CrossRef]
- Stump-Sutliff, K.A. Head and Neck Cancer Treatment Complications. Available online: https://www.webmd.com/cancer/head-neck-cancer-treatment-complications (accessed on 3 February 2023).
- Oral Complications of Chemotherapy and Head/Neck Radiation (PDQ®)–Patient Version—NCI. Available online: https://www.cancer.gov/about-cancer/treatment/side-effects/mouth-throat/oral-complications-pdq (accessed on 30 March 2023).
- Saba, N.F.; Pamulapati, S.; Patel, B.; Mody, M.; Strojan, P.; Takes, R.; Mäkitie, A.A.; Cohen, O.; Pace-Asciak, P.; Vermorken, J.B.; et al. Novel Immunotherapeutic Approaches to Treating HPV-Related Head and Neck Cancer. Cancers 2023, 15, 1959. [Google Scholar] [CrossRef]
- Silver, J.A.; Turkdogan, S.; Roy, C.F.; Subramaniam, T.; Henry, M.; Sadeghi, N. De-Escalation Strategies for Human Papillomavirus-Associated Oropharyngeal Squamous Cell Carcinoma—Where Are We Now? Curr. Oncol. 2022, 29, 3668–3697. [Google Scholar] [CrossRef]
- Hamauchi, S.; Yokota, T.; Mizumachi, T.; Onozawa, Y.; Ogawa, H.; Onoe, T.; Kamijo, T.; Iida, Y.; Nishimura, T.; Onitsuka, T.; et al. Safety and Efficacy of Concurrent Carboplatin or Cetuximab plus Radiotherapy for Locally Advanced Head and Neck Cancer Patients Ineligible for Treatment with Cisplatin. Int. J. Clin. Oncol. 2019, 24, 468–475. [Google Scholar] [CrossRef]
- Mehanna, H.; Rischin, D.; Wong, S.J.; Gregoire, V.; Ferris, R.; Waldron, J.; Le, Q.-T.; Forster, M.; Gillison, M.; Laskar, S.; et al. De-Escalation After DE-ESCALATE and RTOG 1016: A Head and Neck Cancer InterGroup Framework for Future De-Escalation Studies. J. Clin. Oncol. 2020, 38, 2552–2557. [Google Scholar] [CrossRef]
- Gillison, M.L.; D’Souza, G.; Westra, W.; Sugar, E.; Xiao, W.; Begum, S.; Viscidi, R. Distinct Risk Factor Profiles for Human Papillomavirus Type 16-Positive and Human Papillomavirus Type 16-Negative Head and Neck Cancers. J. Natl. Cancer Inst. 2008, 100, 407–420. [Google Scholar] [CrossRef]
- Jethwa, A.R.; Khariwala, S.S. Tobacco-Related Carcinogenesis in Head and Neck Cancer. Cancer Metastasis Rev. 2017, 36, 411–423. [Google Scholar] [CrossRef]
- Secretan, B.; Straif, K.; Baan, R.; Grosse, Y.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; Galichet, L.; et al. A Review of Human Carcinogens—Part E: Tobacco, Areca Nut, Alcohol, Coal Smoke, and Salted Fish. Lancet Oncol. 2009, 10, 1033–1034. [Google Scholar] [CrossRef]
- Stransky, N.; Egloff, A.M.; Tward, A.D.; Kostic, A.D.; Cibulskis, K.; Sivachenko, A.; Kryukov, G.V.; Lawrence, M.S.; Sougnez, C.; McKenna, A.; et al. The Mutational Landscape of Head and Neck Squamous Cell Carcinoma. Science 2011, 333, 1157–1160. [Google Scholar] [CrossRef]
- Agrawal, N.; Frederick, M.J.; Pickering, C.R.; Bettegowda, C.; Chang, K.; Li, R.J.; Fakhry, C.; Xie, T.-X.; Zhang, J.; Wang, J.; et al. Exome Sequencing of Head and Neck Squamous Cell Carcinoma Reveals Inactivating Mutations in NOTCH1. Science 2011, 333, 1154–1157. [Google Scholar] [CrossRef]
- Seiwert, T.Y.; Zuo, Z.; Keck, M.K.; Khattri, A.; Pedamallu, C.S.; Stricker, T.; Brown, C.; Pugh, T.J.; Stojanov, P.; Cho, J.; et al. Integrative and Comparative Genomic Analysis of HPV-Positive and HPV-Negative Head and Neck Squamous Cell Carcinomas. Clin. Cancer Res. 2015, 21, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Yoshinouchi, M.; Yamada, T.; Kizaki, M.; Fen, J.; Koseki, T.; Ikeda, Y.; Nishihara, T.; Yamato, K. In Vitro and In Vivo Growth Suppression of Human Papillomavirus 16-Positive Cervical Cancer Cells by E6 SiRNA. Mol. Ther. 2003, 8, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Jabbar, S.F.; Abrams, L.; Glick, A.; Lambert, P.F. Persistence of High-Grade Cervical Dysplasia and Cervical Cancer Requires the Continuous Expression of the Human Papillomavirus Type 16 E7 Oncogene. Cancer Res. 2009, 69, 4407–4414. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.; Allen-Hoffmann, B.L.; Lambert, P.F. Integration of Human Papillomavirus Type 16 into the Human Genome Correlates with a Selective Growth Advantage of Cells. J. Virol. 1995, 69, 2989–2997. [Google Scholar] [CrossRef]
- Romanczuk, H.; Howley, P.M. Disruption of Either the E1 or the E2 Regulatory Gene of Human Papillomavirus Type 16 Increases Viral Immortalization Capacity. Proc. Natl. Acad. Sci. USA 1992, 89, 3159–3163. [Google Scholar] [CrossRef]
- McBride, A.A.; Warburton, A. The Role of Integration in Oncogenic Progression of HPV-Associated Cancers. PLOS Pathog. 2017, 13, e1006211. [Google Scholar] [CrossRef]
- Matsukura, T.; Koi, S.; Sugase, M. Both Episomal and Integrated Forms of Human Papillomavirus Type 16 Are Involved in Invasive Cervical Cancers. Virology 1989, 172, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.D.; Chen, Z.; Saller, C.; Tarvin, K.; Carvalho, A.L.; Scapulatempo-Neto, C.; Silveira, H.C.; Fregnani, J.H.; Creighton, C.J.; Anderson, M.L.; et al. Integrated Genomic and Molecular Characterization of Cervical Cancer. Nature 2017, 543, 378–384. [Google Scholar] [CrossRef]
- Ramqvist, T.; Mints, M.; Tertipis, N.; Näsman, A.; Romanitan, M.; Dalianis, T. Studies on Human Papillomavirus (HPV) 16 E2, E5 and E7 MRNA in HPV-Positive Tonsillar and Base of Tongue Cancer in Relation to Clinical Outcome and Immunological Parameters. Oral Oncol. 2015, 51, 1126–1131. [Google Scholar] [CrossRef]
- Gleber-Netto, F.O.; Rao, X.; Guo, T.; Xi, Y.; Gao, M.; Shen, L.; Erikson, K.; Kalu, N.N.; Ren, S.; Xu, G.; et al. Variations in HPV Function Are Associated with Survival in Squamous Cell Carcinoma. JCI Insight 2019, 4, e124762. [Google Scholar] [CrossRef]
- Eberhardt, C.S.; Kissick, H.T.; Patel, M.R.; Cardenas, M.A.; Prokhnevska, N.; Obeng, R.C.; Nasti, T.H.; Griffith, C.C.; Im, S.J.; Wang, X.; et al. Functional HPV-Specific PD-1+ Stem-like CD8 T Cells in Head and Neck Cancer. Nature 2021, 597, 279–284. [Google Scholar] [CrossRef]
- Krishna, S.; Ulrich, P.; Wilson, E.; Parikh, F.; Narang, P.; Yang, S.; Read, A.K.; Kim-Schulze, S.; Park, J.G.; Posner, M.; et al. Human Papilloma Virus Specific Immunogenicity and Dysfunction of CD8+ T Cells in Head and Neck Cancer. Cancer Res. 2018, 78, 6159–6170. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.K.; Arsov, C.; Schirlau, K.; Bas, M.; Friebe-Hoffmann, U.; Klussmann, J.P.; Scheckenbach, K.; Balz, V.; Bier, H.; Whiteside, T.L. T Cells Specific for HPV16 E7 Epitopes in Patients with Squamous Cell Carcinoma of the Oropharynx. Int. J. Cancer 2006, 118, 1984–1991. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, K.H.; Neller, M.A.; Srihari, S.; Crooks, P.; Lekieffre, L.; Aftab, B.T.; Liu, H.; Smith, C.; Kenny, L.; Porceddu, S.; et al. Profiling HPV-16–Specific T Cell Responses Reveals Broad Antigen Reactivities in Oropharyngeal Cancer Patients. J. Exp. Med. 2020, 217, e20200389. [Google Scholar] [CrossRef]
- Masterson, L.; Lechner, M.; Loewenbein, S.; Mohammed, H.; Davies-Husband, C.; Fenton, T.; Sudhoff, H.; Jani, P.; Goon, P.; Sterling, J. CD8+ T Cell Response to Human Papillomavirus 16 E7 Is Able to Predict Survival Outcome in Oropharyngeal Cancer. Eur. J. Cancer 2016, 67, 141–151. [Google Scholar] [CrossRef]
- Heusinkveld, M.; Goedemans, R.; Briet, R.; Gelderblom, H.; Nortier, J.; Gorter, A.; Smit, V.; Langeveld, A.; Jansen, J.; van der Burg, S. Systemic and Local Human Papillomavirus 16-Specific T-Cell Immunity in Patients with Head and Neck Cancer. Int. J. Cancer 2012, 131, E74–E85. [Google Scholar] [CrossRef] [PubMed]
- Montes, M.; Rufer, N.; Appay, V.; Reynard, S.; Pittet, M.J.; Speiser, D.E.; Guillaume, P.; Cerottini, J.-C.; Romero, P.; Leyvraz, S. Optimum In Vitro Expansion of Human Antigen-Specific CD8 T Cells for Adoptive Transfer Therapy. Clin. Exp. Immunol. 2005, 142, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Lang Kuhs, K.A.; Kreimer, A.R.; Trivedi, S.; Holzinger, D.; Pawlita, M.; Pfeiffer, R.M.; Gibson, S.P.; Schmitt, N.C.; Hildesheim, A.; Waterboer, T.; et al. HPV16E6 Antibodies Are Sensitive for HPV-Driven Oropharyngeal Cancer and Associated with Recurrence. Cancer 2017, 123, 4382–4390. [Google Scholar] [CrossRef]
- Dahlstrom, K.R.; Anderson, K.S.; Cheng, J.N.; Chowell, D.; Li, G.; Posner, M.; Sturgis, E.M. HPV Serum Antibodies as Predictors of Survival and Disease Progression in Patients with HPV-Positive Squamous Cell Carcinoma of the Oropharynx. Clin. Cancer Res. 2015, 21, 2861–2869. [Google Scholar] [CrossRef]
- Anderson, K.S.; Wong, J.; D’Souza, G.; Riemer, A.B.; Lorch, J.; Haddad, R.; Pai, S.I.; Longtine, J.; McClean, M.; LaBaer, J.; et al. Serum Antibodies to the HPV16 Proteome as Biomarkers for Head and Neck Cancer. Br. J. Cancer 2011, 104, 1896–1905. [Google Scholar] [CrossRef]
- Anderson, K.S.; Dahlstrom, K.R.; Cheng, J.N.; Alam, R.; Li, G.; Wei, Q.; Gross, N.D.; Chowell, D.; Posner, M.; Sturgis, E.M. HPV16 Antibodies as Risk Factors for Oropharyngeal Cancer and Their Association with Tumor HPV and Smoking Status. Oral Oncol. 2015, 51, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Lang Kuhs, K.A.; Pawlita, M.; Gibson, S.P.; Schmitt, N.C.; Trivedi, S.; Argiris, A.; Kreimer, A.R.; Ferris, R.L.; Waterboer, T. Characterization of Human Papillomavirus Antibodies in Individuals with Head and Neck Cancer. Cancer Epidemiol. 2016, 42, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Dahlstrom, K.R.; Anderson, K.S.; Field, M.S.; Chowell, D.; Ning, J.; Li, N.; Wei, Q.; Li, G.; Sturgis, E.M. Diagnostic Accuracy of Serum Antibodies to Human Papillomavirus Type 16 Early Antigens in the Detection of Human Papillomavirus-Related Oropharyngeal Cancer. Cancer 2017, 123, 4886–4894. [Google Scholar] [CrossRef] [PubMed]
- Robbins, H.A.; Ferreiro-Iglesias, A.; Waterboer, T.; Brenner, N.; Nygard, M.; Bender, N.; Schroeder, L.; Hildesheim, A.; Pawlita, M.; D’Souza, G.; et al. Absolute Risk of Oropharyngeal Cancer after an HPV16-E6 Serology Test and Potential Implications for Screening: Results from the Human Papillomavirus Cancer Cohort Consortium. J. Clin. Oncol. 2022, 40, 3613–3622. [Google Scholar] [CrossRef]
- Busch, C.-J.; Hoffmann, A.S.; Viarisio, D.; Becker, B.T.; Rieckmann, T.; Betz, C.; Bender, N.; Schroeder, L.; Hussein, Y.; Petersen, E.; et al. Detection of Stage I HPV-Driven Oropharyngeal Cancer in Asymptomatic Individuals in the Hamburg City Health Study Using HPV16 E6 Serology—A Proof-of-Concept Study. eClinicalMedicine 2022, 53, 101659. [Google Scholar] [CrossRef] [PubMed]
- Spector, M.E.; Sacco, A.G.; Bellile, E.; Taylor, J.M.G.; Jones, T.; Sun, K.; Brown, W.C.; Birkeland, A.C.; Bradford, C.R.; Wolf, G.T.; et al. E6 and E7 Antibody Levels Are Potential Biomarkers of Recurrence in Patients with Advanced Stage Human Papillomavirus Positive Oropharyngeal Squamous Cell Carcinoma. Clin. Cancer Res. 2017, 23, 2723–2729. [Google Scholar] [CrossRef]
- Fakhry, C.; Qualliotine, J.R.; Zhang, Z.; Agrawal, N.; Gaykalova, D.A.; Bishop, J.A.; Subramaniam, R.M.; Koch, W.M.; Chung, C.H.; Eisele, D.W.; et al. Serum Antibodies to HPV16 Early Proteins Warrant Investigation as Potential Biomarkers for Risk Stratification and Recurrence of HPV-Associated Oropharyngeal Cancer. Cancer Prev. Res. 2016, 9, 135–141. [Google Scholar] [CrossRef]
- Zhang, Y.; Waterboer, T.; Haddad, R.I.; Miles, B.A.; Wentz, A.; Gross, N.D.; Fakhry, C.; Quon, H.; Lorch, J.H.; Gourin, C.G.; et al. Human Papillomavirus (HPV) 16 Antibodies at Diagnosis of HPV-Related Oropharyngeal Cancer and Antibody Trajectories after Treatment. Oral Oncol. 2017, 67, 77–82. [Google Scholar] [CrossRef]
- Wieland, A.; Ahmed, R. Fc Receptors in Antimicrobial Protection. Curr. Top Microbiol. Immunol. 2019, 423, 119–150. [Google Scholar] [CrossRef]
- Bruhns, P.; Iannascoli, B.; England, P.; Mancardi, D.A.; Fernandez, N.; Jorieux, S.; Daëron, M. Specificity and Affinity of Human Fcγ Receptors and Their Polymorphic Variants for Human IgG Subclasses. Blood 2009, 113, 3716–3725. [Google Scholar] [CrossRef] [PubMed]
- Bournazos, S.; Ravetch, J.V. Diversification of IgG Effector Functions. Int. Immunol. 2017, 29, 303–310. [Google Scholar] [CrossRef]
- Hudson, W.H.; Wieland, A. Technology Meets TILs: Deciphering T Cell Function in the-Omics Era. Cancer Cell 2023, 41, 41–57. [Google Scholar] [CrossRef]
- Akkaya, M.; Kwak, K.; Pierce, S.K. B Cell Memory: Building Two Walls of Protection against Pathogens. Nat. Rev. Immunol. 2020, 20, 229–238. [Google Scholar] [CrossRef]
- Weisel, F.; Shlomchik, M. Memory B Cells of Mice and Humans. Annu. Rev. Immunol. 2017, 35, 255–284. [Google Scholar] [CrossRef]
- Davis, C.W.; Jackson, K.J.L.; McCausland, M.M.; Darce, J.; Chang, C.; Linderman, S.L.; Chennareddy, C.; Gerkin, R.; Brown, S.J.; Wrammert, J.; et al. Influenza Vaccine–Induced Human Bone Marrow Plasma Cells Decline within a Year after Vaccination. Science 2020, 370, 237–241. [Google Scholar] [CrossRef]
- Portugal, S.; Obeng-Adjei, N.; Moir, S.; Crompton, P.D.; Pierce, S.K. Atypical Memory B Cells in Human Chronic Infectious Diseases: An Interim Report. Cell. Immunol. 2017, 321, 18–25. [Google Scholar] [CrossRef]
- Slifka, M.K.; Ahmed, R. Limiting Dilution Analysis of Virus-Specific Memory B Cells by an ELISPOT Assay. J. Immunol. Methods 1996, 199, 37–46. [Google Scholar] [CrossRef]
- Crotty, S.; Aubert, R.D.; Glidewell, J.; Ahmed, R. Tracking Human Antigen-Specific Memory B Cells: A Sensitive and Generalized ELISPOT System. J. Immunol. Methods 2004, 286, 111–122. [Google Scholar] [CrossRef]
- Jahnmatz, M.; Kesa, G.; Netterlid, E.; Buisman, A.-M.; Thorstensson, R.; Ahlborg, N. Optimization of a Human IgG B-Cell ELISpot Assay for the Analysis of Vaccine-Induced B-Cell Responses. J. Immunol. Methods 2013, 391, 50–59. [Google Scholar] [CrossRef]
- Cillo, A.R.; Kürten, C.H.L.; Tabib, T.; Qi, Z.; Onkar, S.; Wang, T.; Liu, A.; Duvvuri, U.; Kim, S.; Soose, R.J.; et al. Immune Landscape of Viral- and Carcinogen-Driven Head and Neck Cancer. Immunity 2020, 52, 183–199.e9. [Google Scholar] [CrossRef]
- Russell, S.; Angell, T.; Lechner, M.; Liebertz, D.; Correa, A.; Sinha, U.; Kokot, N.; Epstein, A. Immune Cell Infiltration Patterns and Survival in Head and Neck Squamous Cell Carcinoma. Head Neck Oncol. 2013, 5, 24. [Google Scholar]
- Wood, O.; Woo, J.; Seumois, G.; Savelyeva, N.; McCann, K.J.; Singh, D.; Jones, T.; Peel, L.; Breen, M.S.; Ward, M.; et al. Gene Expression Analysis of TIL Rich HPV-Driven Head and Neck Tumors Reveals a Distinct B-Cell Signature When Compared to HPV Independent Tumors. Oncotarget 2016, 7, 56781–56797. [Google Scholar] [CrossRef]
- Ruffin, A.T.; Cillo, A.R.; Tabib, T.; Liu, A.; Onkar, S.; Kunning, S.R.; Lampenfeld, C.; Atiya, H.I.; Abecassis, I.; Kürten, C.H.L.; et al. B Cell Signatures and Tertiary Lymphoid Structures Contribute to Outcome in Head and Neck Squamous Cell Carcinoma. Nat. Commun. 2021, 12, 3349. [Google Scholar] [CrossRef]
- Kansy, B.A.; Concha-Benavente, F.; Srivastava, R.M.; Jie, H.-B.; Shayan, G.; Lei, Y.; Moskovitz, J.; Moy, J.; Li, J.; Brandau, S.; et al. PD-1 Status in CD8+ T Cells Associates with Survival and Anti-PD-1 Therapeutic Outcomes in Head and Neck Cancer. Cancer Res. 2017, 77, 6353–6364. [Google Scholar] [CrossRef]
- Gameiro, S.F.; Ghasemi, F.; Barrett, J.W.; Koropatnick, J.; Nichols, A.C.; Mymryk, J.S.; Maleki Vareki, S. Treatment-Naïve HPV+ Head and Neck Cancers Display a T-Cell-Inflamed Phenotype Distinct from Their HPV- Counterparts That Has Implications for Immunotherapy. Oncoimmunology 2018, 7, e1498439. [Google Scholar] [CrossRef]
- Lechner, A.; Schlößer, H.; Rothschild, S.I.; Thelen, M.; Reuter, S.; Zentis, P.; Shimabukuro-Vornhagen, A.; Theurich, S.; Wennhold, K.; Garcia-Marquez, M.; et al. Characterization of Tumor-Associated T-Lymphocyte Subsets and Immune Checkpoint Molecules in Head and Neck Squamous Cell Carcinoma. Oncotarget 2017, 8, 44418–44433. [Google Scholar] [CrossRef]
- Lechner, A.; Schlößer, H.A.; Thelen, M.; Wennhold, K.; Rothschild, S.I.; Gilles, R.; Quaas, A.; Siefer, O.G.; Huebbers, C.U.; Cukuroglu, E.; et al. Tumor-Associated B Cells and Humoral Immune Response in Head and Neck Squamous Cell Carcinoma. Oncoimmunology 2019, 8, 1535293. [Google Scholar] [CrossRef]
- Crotty, S. T Follicular Helper Cell Differentiation, Function, and Roles in Disease. Immunity 2014, 41, 529–542. [Google Scholar] [CrossRef]
- Helmink, B.A.; Reddy, S.M.; Gao, J.; Zhang, S.; Basar, R.; Thakur, R.; Yizhak, K.; Sade-Feldman, M.; Blando, J.; Han, G.; et al. B Cells and Tertiary Lymphoid Structures Promote Immunotherapy Response. Nature 2020, 577, 549–555. [Google Scholar] [CrossRef]
- Cabrita, R.; Lauss, M.; Sanna, A.; Donia, M.; Skaarup Larsen, M.; Mitra, S.; Johansson, I.; Phung, B.; Harbst, K.; Vallon-Christersson, J.; et al. Tertiary Lymphoid Structures Improve Immunotherapy and Survival in Melanoma. Nature 2020, 577, 561–565. [Google Scholar] [CrossRef]
- Petitprez, F.; de Reyniès, A.; Keung, E.Z.; Chen, T.W.-W.; Sun, C.-M.; Calderaro, J.; Jeng, Y.-M.; Hsiao, L.-P.; Lacroix, L.; Bougoüin, A.; et al. B Cells Are Associated with Survival and Immunotherapy Response in Sarcoma. Nature 2020, 577, 556–560. [Google Scholar] [CrossRef]
- Schumacher, T.N.; Thommen, D.S. Tertiary Lymphoid Structures in Cancer. Science 2022, 375, eabf9419. [Google Scholar] [CrossRef] [PubMed]
- Oguejiofor, K.; Hall, J.; Slater, C.; Betts, G.; Hall, G.; Slevin, N.; Dovedi, S.; Stern, P.L.; West, C.M.L. Stromal Infiltration of CD8 T Cells Is Associated with Improved Clinical Outcome in HPV-Positive Oropharyngeal Squamous Carcinoma. Br. J. Cancer 2015, 113, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Näsman, A.; Romanitan, M.; Nordfors, C.; Grün, N.; Johansson, H.; Hammarstedt, L.; Marklund, L.; Munck-Wikland, E.; Dalianis, T.; Ramqvist, T. Tumor Infiltrating CD8+ and Foxp3+ Lymphocytes Correlate to Clinical Outcome and Human Papillomavirus (HPV) Status in Tonsillar Cancer. PLoS ONE 2012, 7, e38711. [Google Scholar] [CrossRef]
- Cheng, Y.; Gunasegaran, B.; Singh, H.D.; Dutertre, C.-A.; Loh, C.Y.; Lim, J.Q.; Crawford, J.C.; Lee, H.K.; Zhang, X.; Lee, B.; et al. Non-Terminally Exhausted Tumor-Resident Memory HBV-Specific T Cell Responses Correlate with Relapse-Free Survival in Hepatocellular Carcinoma. Immunity 2021, 54, 1825–1840.e7. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.; Stromhaug, K.; Klaeger, S.; Kula, T.; Frederick, D.T.; Le, P.M.; Forman, J.; Huang, T.; Li, S.; Zhang, W.; et al. Phenotype, Specificity and Avidity of Antitumour CD8+ T Cells in Melanoma. Nature 2021, 596, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Rosato, P.C.; Wijeyesinghe, S.; Stolley, J.M.; Nelson, C.E.; Davis, R.L.; Manlove, L.S.; Pennell, C.A.; Blazar, B.R.; Chen, C.C.; Geller, M.A.; et al. Virus-Specific Memory T Cells Populate Tumors and Can Be Repurposed for Tumor Immunotherapy. Nat. Commun. 2019, 10, 567. [Google Scholar] [CrossRef]
- Simoni, Y.; Becht, E.; Fehlings, M.; Loh, C.Y.; Koo, S.-L.; Teng, K.W.W.; Yeong, J.P.S.; Nahar, R.; Zhang, T.; Kared, H.; et al. Bystander CD8+ T Cells Are Abundant and Phenotypically Distinct in Human Tumour Infiltrates. Nature 2018, 557, 575–579. [Google Scholar] [CrossRef]
- Scheper, W.; Kelderman, S.; Fanchi, L.F.; Linnemann, C.; Bendle, G.; de Rooij, M.A.J.; Hirt, C.; Mezzadra, R.; Slagter, M.; Dijkstra, K.; et al. Low and Variable Tumor Reactivity of the Intratumoral TCR Repertoire in Human Cancers. Nat. Med. 2019, 25, 89–94. [Google Scholar] [CrossRef]
- Sudmeier, L.J.; Hoang, K.B.; Nduom, E.K.; Wieland, A.; Neill, S.G.; Schniederjan, M.J.; Ramalingam, S.S.; Olson, J.J.; Ahmed, R.; Hudson, W.H. Distinct Phenotypic States and Spatial Distribution of CD8+ T Cell Clonotypes in Human Brain Metastases. Cell Rep. Med. 2022, 3, 100620. [Google Scholar] [CrossRef]
- Sekine, T.; Perez-Potti, A.; Nguyen, S.; Gorin, J.-B.; Wu, V.H.; Gostick, E.; Llewellyn-Lacey, S.; Hammer, Q.; Falck-Jones, S.; Vangeti, S.; et al. TOX Is Expressed by Exhausted and Polyfunctional Human Effector Memory CD8+ T Cells. Sci. Immunol. 2020, 5, eaba7918. [Google Scholar] [CrossRef]
- Ning, J.; Gavil, N.V.; Wu, S.; Wijeyesinghe, S.; Weyu, E.; Ma, J.; Li, M.; Grigore, F.-N.; Dhawan, S.; Skorput, A.G.J.; et al. Functional Virus-Specific Memory T Cells Survey Glioblastoma. Cancer Immunol. Immunother. 2022, 71, 1863–1875. [Google Scholar] [CrossRef] [PubMed]
- Çuburu, N.; Bialkowski, L.; Pontejo, S.M.; Sethi, S.K.; Bell, A.T.F.; Kim, R.; Thompson, C.D.; Lowy, D.R.; Schiller, J.T. Harnessing Anti-Cytomegalovirus Immunity for Local Immunotherapy against Solid Tumors. Proc. Natl. Acad. Sci. USA 2022, 119, e2116738119. [Google Scholar] [CrossRef] [PubMed]
- Welters, M.J.P.; Ma, W.; Santegoets, S.J.A.M.; Goedemans, R.; Ehsan, I.; Jordanova, E.S.; van Ham, V.J.; van Unen, V.; Koning, F.; van Egmond, S.I.; et al. Intratumoral HPV16-Specific T Cells Constitute a Type I–Oriented Tumor Microenvironment to Improve Survival in HPV16-Driven Oropharyngeal Cancer. Clin. Cancer Res. 2018, 24, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Santegoets, S.J.; Welters, M.J.P.; Schrikkema, D.S.; Freriks, M.R.; Kok, H.; Weissbrich, B.; van den Branden, A.; Linnemann, C.; Schumacher, T.N.; Adhikary, S.; et al. The Common HLA Class I-Restricted Tumor-Infiltrating T Cell Response in HPV16-Induced Cancer. Cancer Immunol. Immunother. 2022, 72, 1553–1565. [Google Scholar] [CrossRef]
- McInnis, C.; Bhatia, S.; Vijaykumar, B.; Tian, Q.; Sun, Y.; Leistritz-Edwards, D.; Quinn, C.T.; Uppaluri, R.; Egloff, A.M.; Srinivasan, L.; et al. Identification of HPV16 E1 and E2-Specific T Cells in the Oropharyngeal Cancer Tumor Microenvironment. J. Immunother. Cancer 2023, 11, e006721. [Google Scholar] [CrossRef]
- Im, S.J.; Hashimoto, M.; Gerner, M.Y.; Lee, J.; Kissick, H.T.; Burger, M.C.; Shan, Q.; Hale, J.S.; Lee, J.; Nasti, T.H.; et al. Defining CD8+ T Cells That Provide the Proliferative Burst after PD-1 Therapy. Nature 2016, 537, 417–421. [Google Scholar] [CrossRef]
- He, R.; Hou, S.; Liu, C.; Zhang, A.; Bai, Q.; Han, M.; Yang, Y.; Wei, G.; Shen, T.; Yang, X.; et al. Follicular CXCR5- Expressing CD8(+) T Cells Curtail Chronic Viral Infection. Nature 2016, 537, 412–428. [Google Scholar] [CrossRef]
- Utzschneider, D.T.; Charmoy, M.; Chennupati, V.; Pousse, L.; Ferreira, D.P.; Calderon-Copete, S.; Danilo, M.; Alfei, F.; Hofmann, M.; Wieland, D.; et al. T Cell Factor 1-Expressing Memory-like CD8(+) T Cells Sustain the Immune Response to Chronic Viral Infections. Immunity 2016, 45, 415–427. [Google Scholar] [CrossRef]
- Wu, T.; Ji, Y.; Moseman, E.A.; Xu, H.C.; Manglani, M.; Kirby, M.; Anderson, S.M.; Handon, R.; Kenyon, E.; Elkahloun, A.; et al. The TCF1-Bcl6 Axis Counteracts Type I Interferon to Repress Exhaustion and Maintain T Cell Stemness. Sci. Immunol. 2016, 1, eaai8593. [Google Scholar] [CrossRef]
- Miller, B.C.; Sen, D.R.; Al Abosy, R.; Bi, K.; Virkud, Y.V.; LaFleur, M.W.; Yates, K.B.; Lako, A.; Felt, K.; Naik, G.S.; et al. Subsets of Exhausted CD8+ T Cells Differentially Mediate Tumor Control and Respond to Checkpoint Blockade. Nat. Immunol. 2019, 20, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, I.; Schaeuble, K.; Chennupati, V.; Fuertes Marraco, S.A.; Calderon-Copete, S.; Pais Ferreira, D.; Carmona, S.J.; Scarpellino, L.; Gfeller, D.; Pradervand, S.; et al. Intratumoral Tcf1+PD-1+CD8+ T Cells with Stem-like Properties Promote Tumor Control in Response to Vaccination and Checkpoint Blockade Immunotherapy. Immunity 2019, 50, 195–211.e10. [Google Scholar] [CrossRef] [PubMed]
- Sade-Feldman, M.; Yizhak, K.; Bjorgaard, S.L.; Ray, J.P.; de Boer, C.G.; Jenkins, R.W.; Lieb, D.J.; Chen, J.H.; Frederick, D.T.; Barzily-Rokni, M.; et al. Defining T Cell States Associated with Response to Checkpoint Immunotherapy in Melanoma. Cell 2018, 175, 998–1013.e20. [Google Scholar] [CrossRef] [PubMed]
- Jansen, C.S.; Prokhnevska, N.; Master, V.A.; Sanda, M.G.; Carlisle, J.W.; Bilen, M.A.; Cardenas, M.; Wilkinson, S.; Lake, R.; Sowalsky, A.G.; et al. An Intra-Tumoral Niche Maintains and Differentiates Stem-like CD8 T Cells. Nature 2019, 576, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Hudson, W.H.; Gensheimer, J.; Hashimoto, M.; Wieland, A.; Valanparambil, R.M.; Li, P.; Lin, J.-X.; Konieczny, B.T.; Im, S.J.; Freeman, G.J.; et al. Proliferating Transitory T Cells with an Effector-like Transcriptional Signature Emerge from PD-1+ Stem-like CD8+ T Cells during Chronic Infection. Immunity 2019, 51, 1043–1058.e4. [Google Scholar] [CrossRef]
- Zander, R.; Schauder, D.; Xin, G.; Nguyen, C.; Wu, X.; Zajac, A.; Cui, W. CD4+ T Cell Help Is Required for the Formation of a Cytolytic CD8+ T Cell Subset That Protects against Chronic Infection and Cancer. Immunity 2019, 51, 1028–1042.e4. [Google Scholar] [CrossRef]
- Prabhakar, A.T.; James, C.D.; Das, D.; Otoa, R.; Day, M.; Burgner, J.; Fontan, C.T.; Wang, X.; Glass, S.H.; Wieland, A.; et al. CK2 Phosphorylation of Human Papillomavirus 16 E2 on Serine 23 Promotes Interaction with TopBP1 and Is Critical for E2 Interaction with Mitotic Chromatin and the Viral Life Cycle. mBio 2021, 12, e0116321. [Google Scholar] [CrossRef]
- Ellebedy, A.H.; Jackson, K.J.L.; Kissick, H.T.; Nakaya, H.I.; Davis, C.W.; Roskin, K.M.; McElroy, A.K.; Oshansky, C.M.; Elbein, R.; Thomas, S.; et al. Defining Antigen-Specific Plasmablast and Memory B Cell Subsets in Human Blood after Viral Infection or Vaccination. Nat. Immunol. 2016, 17, 1226–1234. [Google Scholar] [CrossRef]
- Li, G.-M.; Chiu, C.; Wrammert, J.; McCausland, M.; Andrews, S.F.; Zheng, N.-Y.; Lee, J.-H.; Huang, M.; Qu, X.; Edupuganti, S.; et al. Pandemic H1N1 Influenza Vaccine Induces a Recall Response in Humans That Favors Broadly Cross-Reactive Memory B Cells. Proc. Natl. Acad. Sci. USA 2012, 109, 9047–9052. [Google Scholar] [CrossRef]
- Wrammert, J.; Koutsonanos, D.; Li, G.-M.; Edupuganti, S.; Sui, J.; Morrissey, M.; McCausland, M.; Skountzou, I.; Hornig, M.; Lipkin, W.I.; et al. Broadly Cross-Reactive Antibodies Dominate the Human B Cell Response against 2009 Pandemic H1N1 Influenza Virus Infection. J. Exp. Med. 2011, 208, 181–193. [Google Scholar] [CrossRef]
- Wrammert, J.; Smith, K.; Miller, J.; Langley, W.A.; Kokko, K.; Larsen, C.; Zheng, N.-Y.; Mays, I.; Garman, L.; Helms, C.; et al. Rapid Cloning of High-Affinity Human Monoclonal Antibodies against Influenza Virus. Nature 2008, 453, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.W.; Jackson, K.J.L.; McElroy, A.K.; Halfmann, P.; Huang, J.; Chennareddy, C.; Piper, A.E.; Leung, Y.; Albariño, C.G.; Crozier, I.; et al. Longitudinal Analysis of the Human B Cell Response to Ebola Virus Infection. Cell 2019, 177, 1566–1582.e17. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, N.L.; Reuter-Monslow, M.A.; Sei, J.; Durr, E.; Davis, C.W.; Chang, C.; McCausland, M.; Wieland, A.; Krah, D.; Rouphael, N.; et al. Breadth and Functionality of Varicella-Zoster Virus Glycoprotein-Specific Antibodies Identified after Zostavax Vaccination in Humans. J. Virol. 2018, 92, e00269-18. [Google Scholar] [CrossRef] [PubMed]
- Colbert, J.D.; Cruz, F.M.; Rock, K.L. Cross-Presentation of Exogenous Antigens on MHC I Molecules. Curr. Opin. Immunol. 2020, 64, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Twomey, J.D.; Zhang, B. Cancer Immunotherapy Update: FDA-Approved Checkpoint Inhibitors and Companion Diagnostics. AAPS J. 2021, 23, 39. [Google Scholar] [CrossRef]
- Patel, J.J.; Levy, D.A.; Nguyen, S.A.; Knochelmann, H.M.; Day, T.A. Impact of PD-L1 Expression and Human Papillomavirus Status in Anti-PD1/PDL1 Immunotherapy for Head and Neck Squamous Cell Carcinoma-Systematic Review and Meta-Analysis. Head Neck 2020, 42, 774–786. [Google Scholar] [CrossRef]
- Smalley Rumfield, C.; Roller, N.; Pellom, S.T.; Schlom, J.; Jochems, C. Therapeutic Vaccines for HPV-Associated Malignancies. Immunotargets Ther. 2020, 9, 167–200. [Google Scholar] [CrossRef]
- Ha, S.-J.; Mueller, S.N.; Wherry, E.J.; Barber, D.L.; Aubert, R.D.; Sharpe, A.H.; Freeman, G.J.; Ahmed, R. Enhancing Therapeutic Vaccination by Blocking PD-1-Mediated Inhibitory Signals during Chronic Infection. J. Exp. Med. 2008, 205, 543–555. [Google Scholar] [CrossRef]
- Kamphorst, A.O.; Wieland, A.; Nasti, T.; Yang, S.; Zhang, R.; Barber, D.L.; Konieczny, B.T.; Daugherty, C.Z.; Koenig, L.; Yu, K.; et al. Rescue of Exhausted CD8 T Cells by PD-1 Targeted Therapies Is CD28-Dependent. Science 2017, 355, 1423–1427. [Google Scholar] [CrossRef]
- Saxena, M.; van der Burg, S.H.; Melief, C.J.M.; Bhardwaj, N. Therapeutic Cancer Vaccines. Nat. Rev. Cancer 2021, 21, 360–378. [Google Scholar] [CrossRef]
- Kenter, G.G.; Welters, M.J.P.; Valentijn, A.R.P.M.; Lowik, M.J.G.; Berends-van der Meer, D.M.A.; Vloon, A.P.G.; Essahsah, F.; Fathers, L.M.; Offringa, R.; Drijfhout, J.W.; et al. Vaccination against HPV-16 Oncoproteins for Vulvar Intraepithelial Neoplasia. N. Engl. J. Med. 2009, 361, 1838–1847. [Google Scholar] [CrossRef]
- van Poelgeest, M.I.E.; Welters, M.J.P.; Vermeij, R.; Stynenbosch, L.F.M.; Loof, N.M.; Berends-van der Meer, D.M.A.; Löwik, M.J.G.; Hamming, I.L.E.; van Esch, E.M.G.; Hellebrekers, B.W.J.; et al. Vaccination against Oncoproteins of HPV16 for Noninvasive Vulvar/Vaginal Lesions: Lesion Clearance Is Related to the Strength of the T-Cell Response. Clin. Cancer Res. 2016, 22, 2342–2350. [Google Scholar] [CrossRef]
- Kenter, G.G.; Welters, M.J.P.; Valentijn, A.R.P.M.; Lowik, M.J.G.; Berends-van der Meer, D.M.A.; Vloon, A.P.G.; Drijfhout, J.W.; Wafelman, A.R.; Oostendorp, J.; Fleuren, G.J.; et al. Phase I Immunotherapeutic Trial with Long Peptides Spanning the E6 and E7 Sequences of High-Risk Human Papillomavirus 16 in End-Stage Cervical Cancer Patients Shows Low Toxicity and Robust Immunogenicity. Clin. Cancer Res. 2008, 14, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Massarelli, E.; William, W.; Johnson, F.; Kies, M.; Ferrarotto, R.; Guo, M.; Feng, L.; Lee, J.J.; Tran, H.; Kim, Y.U.; et al. Combining Immune Checkpoint Blockade and Tumor-Specific Vaccine for Patients with Incurable Human Papillomavirus 16–Related Cancer: A Phase 2 Clinical Trial. JAMA Oncol. 2019, 5, 67–73. [Google Scholar] [CrossRef]
- Melief, C.J.M.; Welters, M.J.P.; Vergote, I.; Kroep, J.R.; Kenter, G.G.; Ottevanger, P.B.; Tjalma, W.A.A.; Denys, H.; van Poelgeest, M.I.E.; Nijman, H.W.; et al. Strong Vaccine Responses during Chemotherapy Are Associated with Prolonged Cancer Survival. Sci. Transl. Med. 2020, 12, eaaz8235. [Google Scholar] [CrossRef] [PubMed]
- Welters, M.J.; van der Sluis, T.C.; van Meir, H.; Loof, N.M.; van Ham, V.J.; van Duikeren, S.; Santegoets, S.J.; Arens, R.; de Kam, M.L.; Cohen, A.F.; et al. Vaccination during Myeloid Cell Depletion by Cancer Chemotherapy Fosters Robust T Cell Responses. Sci. Transl. Med. 2016, 8, 334ra52. [Google Scholar] [CrossRef]
- Stevanović, S.; Draper, L.M.; Langhan, M.M.; Campbell, T.E.; Kwong, M.L.; Wunderlich, J.R.; Dudley, M.E.; Yang, J.C.; Sherry, R.M.; Kammula, U.S.; et al. Complete Regression of Metastatic Cervical Cancer after Treatment with Human Papillomavirus-Targeted Tumor-Infiltrating T Cells. J. Clin. Oncol. 2015, 33, 1543–1550. [Google Scholar] [CrossRef]
- Stevanović, S.; Pasetto, A.; Helman, S.R.; Gartner, J.J.; Prickett, T.D.; Howie, B.; Robins, H.S.; Robbins, P.F.; Klebanoff, C.A.; Rosenberg, S.A.; et al. Landscape of Immunogenic Tumor Antigens in Successful Immunotherapy of Virally Induced Epithelial Cancer. Science 2017, 356, 200–205. [Google Scholar] [CrossRef]
- Stevanović, S.; Helman, S.R.; Wunderlich, J.R.; Langhan, M.M.; Doran, S.L.; Kwong, M.L.M.; Somerville, R.P.T.; Klebanoff, C.A.; Kammula, U.S.; Sherry, R.M.; et al. A Phase II Study of Tumor-Infiltrating Lymphocyte Therapy for Human Papillomavirus-Associated Epithelial Cancers. Clin. Cancer Res. 2019, 25, 1486–1493. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.Y.; Campbell, T.E.; Draper, L.M.; Stevanović, S.; Weissbrich, B.; Yu, Z.; Restifo, N.P.; Rosenberg, S.A.; Trimble, C.L.; Hinrichs, C.S. Engineered T Cells Targeting E7 Mediate Regression of Human Papillomavirus Cancers in a Murine Model. JCI Insight 2018, 3, e99488. [Google Scholar] [CrossRef]
- Doran, S.L.; Stevanović, S.; Adhikary, S.; Gartner, J.J.; Jia, L.; Kwong, M.L.M.; Faquin, W.C.; Hewitt, S.M.; Sherry, R.M.; Yang, J.C.; et al. T-Cell Receptor Gene Therapy for Human Papillomavirus-Associated Epithelial Cancers: A First-in-Human, Phase I/II Study. J. Clin. Oncol. 2019, 37, 2759–2768. [Google Scholar] [CrossRef] [PubMed]
- Nagarsheth, N.B.; Norberg, S.M.; Sinkoe, A.L.; Adhikary, S.; Meyer, T.J.; Lack, J.B.; Warner, A.C.; Schweitzer, C.; Doran, S.L.; Korrapati, S.; et al. TCR-Engineered T Cells Targeting E7 for Patients with Metastatic HPV-Associated Epithelial Cancers. Nat. Med. 2021, 27, 419–425. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conarty, J.P.; Wieland, A. The Tumor-Specific Immune Landscape in HPV+ Head and Neck Cancer. Viruses 2023, 15, 1296. https://doi.org/10.3390/v15061296
Conarty JP, Wieland A. The Tumor-Specific Immune Landscape in HPV+ Head and Neck Cancer. Viruses. 2023; 15(6):1296. https://doi.org/10.3390/v15061296
Chicago/Turabian StyleConarty, Jacob P., and Andreas Wieland. 2023. "The Tumor-Specific Immune Landscape in HPV+ Head and Neck Cancer" Viruses 15, no. 6: 1296. https://doi.org/10.3390/v15061296