
Evidence of Lumpy Skin Disease Virus Transmission from Subclinically Infected Cattle by Stomoxys calcitrans
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Strain
2.2. Vectors
2.3. General Experimental Set-Up
2.4. Experimental Inoculation of Donor Animals
2.5. Disease Status of LSDV-Inoculated Animals
2.6. Placement and Feeding of S. calcitrans on Donor and Acceptor Cattle
2.7. Clinical Scoring
2.8. Sample Collection
2.9. DNA Extraction and Real-Time PCR
2.10. Serological Analysis
2.11. IFNγ Release Assay
3. Result
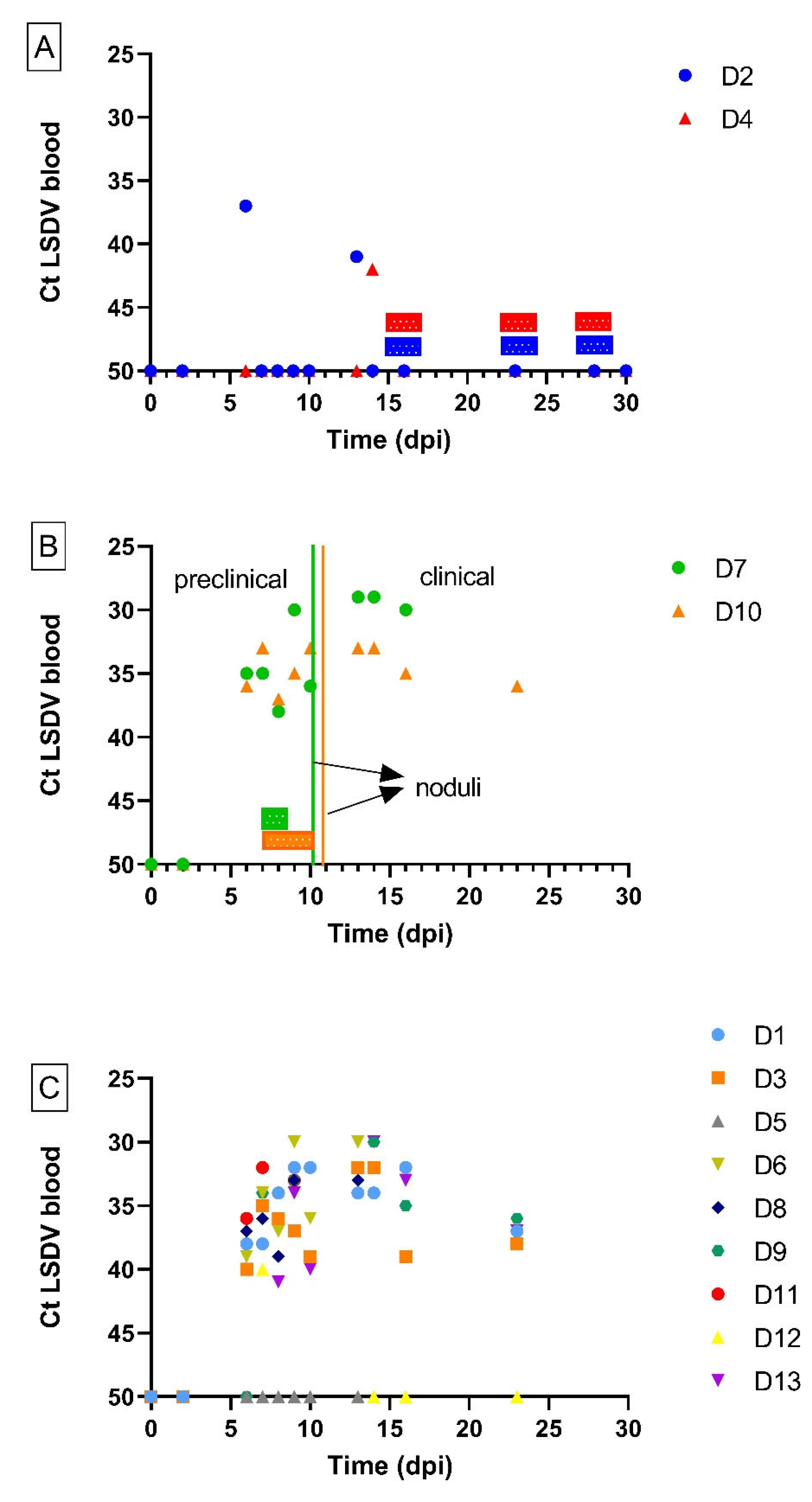
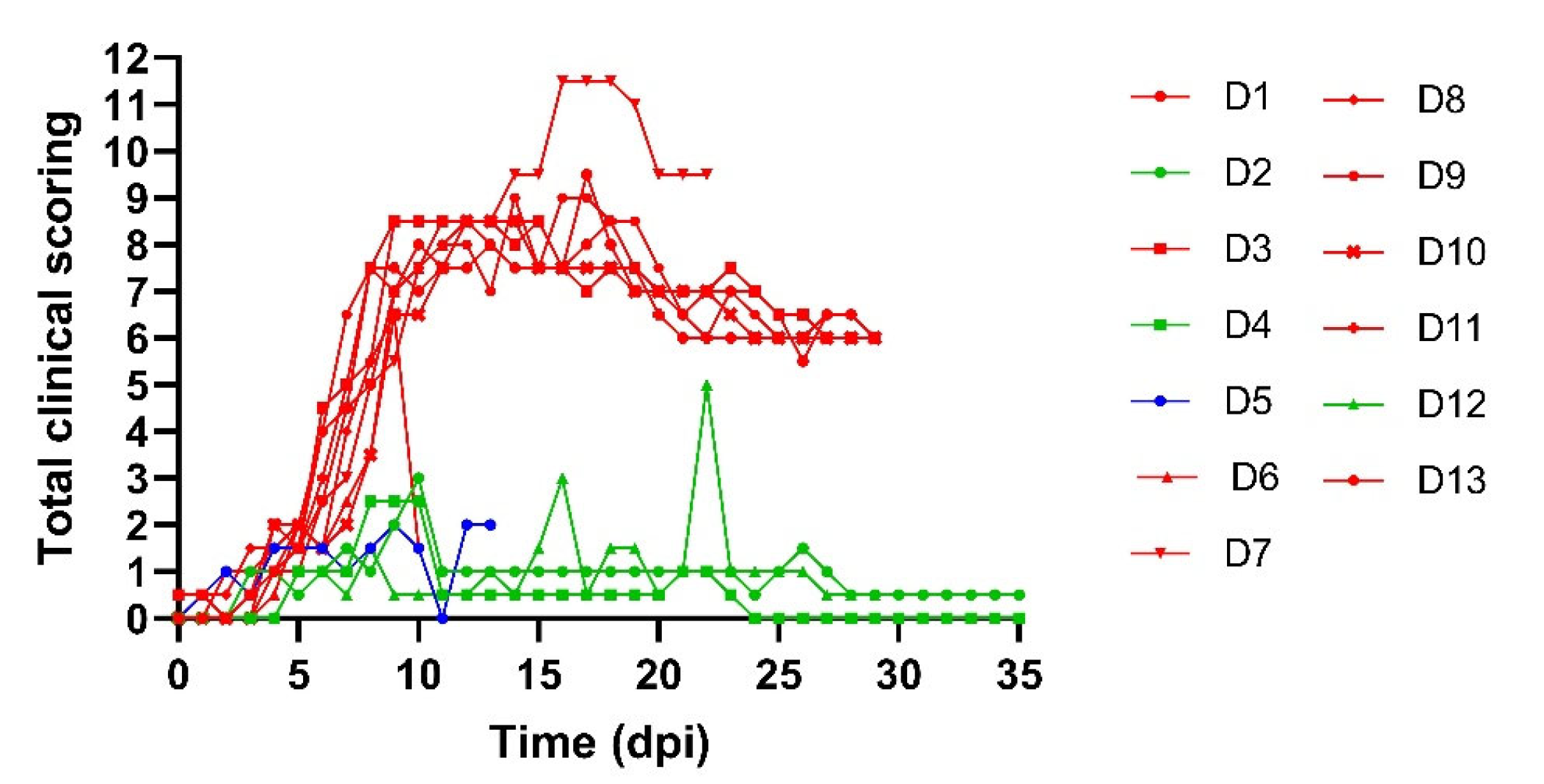
3.1. Donor Animals
3.2. Transmission from Preclinical Cattle
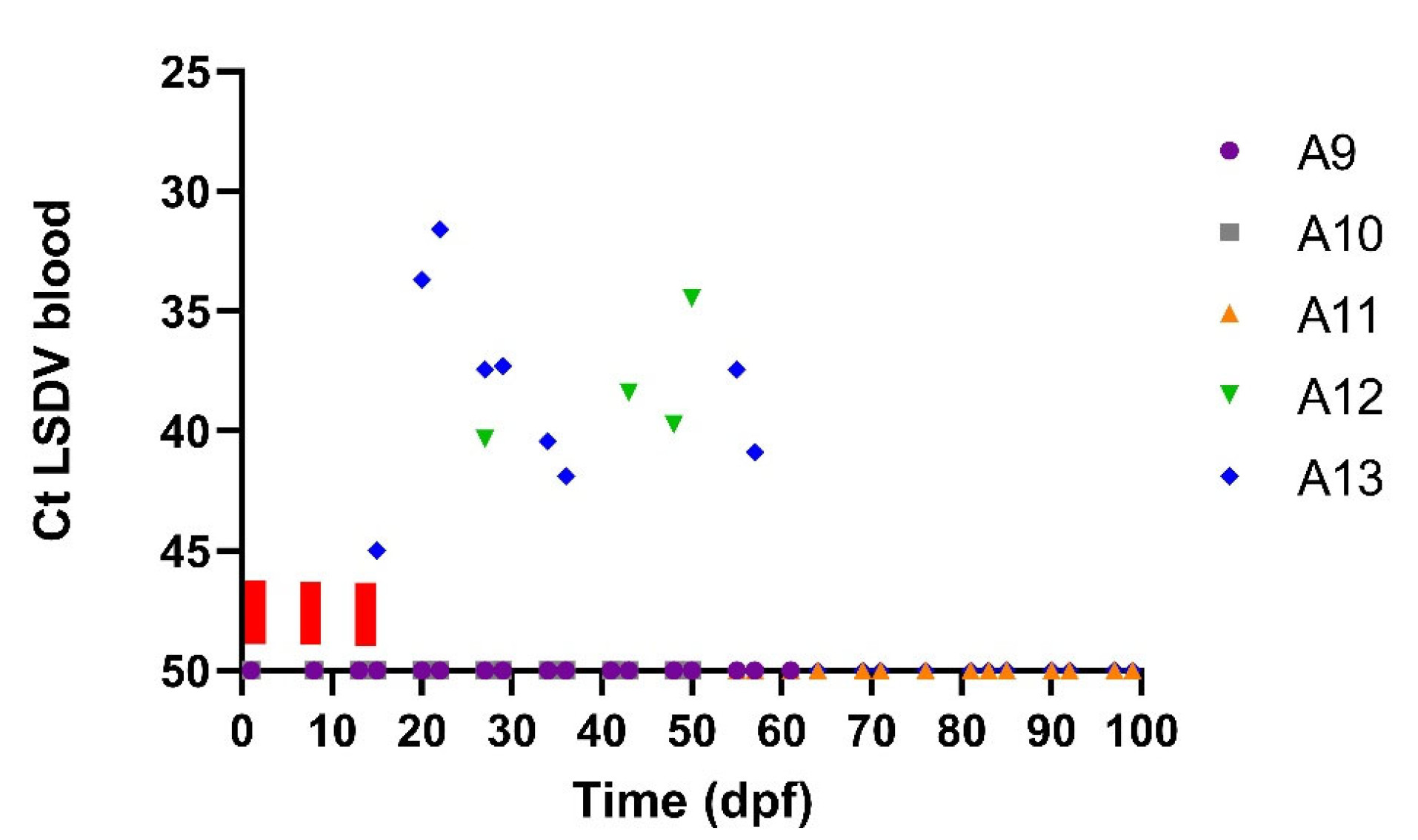
3.3. Transmission from Subclinical Animals
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tuppurainen, E.S.M.; Oura, C.A.L. Review: Lumpy Skin Disease: An Emerging Threat to Europe, the Middle East and Asia. Transbound. Emerg. Dis. 2012, 59, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Mulatu, E.; Feyisa, A. Review: Lumpy Skin Disease. J. Veter Sci. Technol. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Sanz-Bernardo, B.; Haga, I.R.; Wijesiriwardana, N.; Basu, S.; Larner, W.; Diaz, A.V.; Langlands, Z.; Denison, E.; Stoner, J.; White, M.; et al. Quantifying and Modeling the Acquisition and Retention of Lumpy Skin Disease Virus by Hematophagus Insects Reveals Clinically but Not Subclinically Affected Cattle Are Promoters of Viral Transmission and Key Targets for Control of Disease Outbreaks. J. Virol. 2021, 95, e02239-20. [Google Scholar] [CrossRef] [PubMed]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Mechanical transmission of lumpy skin disease virus by Aedes aegypti (Diptera: Culicidae). Epidemiol. Infect. 2001, 126, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.S.M.; Lubinga, J.C.; Stoltsz, W.H.; Troskie, M.; Carpenter, S.T.; Coetzer, J.A.W.; Venter, E.H.; Oura, C.A.L. Mechanical transmission of lumpy skin disease virus by Rhipicephalus appendiculatus male ticks. Epidemiol. Infect. 2013, 141, 425–430. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.; Lubinga, J.C.; Stoltsz, W.H.; Troskie, M.; Carpenter, S.T.; Coetzer, J.A.; Venter, E.H.; Oura, C.A. Evidence of vertical transmission of lumpy skin disease virus in Rhipicephalus decoloratus ticks. Ticks Tick Borne Dis. 2013, 4, 329–333. [Google Scholar] [CrossRef]
- Lubinga, J.C.; Tuppurainen, E.S.M.; Stoltsz, W.H.; Ebersohn, K.; Coetzer, J.A.W.; Venter, E. Detection of lumpy skin disease virus in saliva of ticks fed on lumpy skin disease virus-infected cattle. Exp. Appl. Acarol. 2013, 61, 129–138. [Google Scholar] [CrossRef]
- Lubinga, J.C.; Clift, S.J.; Tuppurainen, E.S.; Stoltsz, W.H.; Babiuk, S.; Coetzer, J.A.; Venter, E.H. Demonstration of lumpy skin disease virus infection in Amblyomma hebraeum and Rhipicephalus appendiculatus ticks using immunohistochemistry. Ticks Tick-Borne Dis. 2014, 5, 113–120. [Google Scholar]
- Tuppurainen, E.S.M.; Stoltsz, W.H.; Troskie, M.; Wallace, D.B.; Oura, C.A.L.; Mellor, P.S.; Coetzer, J.A.W.; Venter, E. A potential role for ixodid (hard) tick vectors in the transmission of lumpy skin disease virus in cattle. Transbound. Emerg. Dis. 2011, 58, 93–104. [Google Scholar]
- Sohier, C.; Haegeman, A.; Mostin, L.; De Leeuw, I.; Van Campe, W.; De Vleeschauwer, A.; Tuppurainen, E.S.M.; van den Berg, T.; De Regge, N.; De Clercq, K. Experimental evidence of mechanical lumpy skin disease virus transmission by Stomoxys calcitrans biting flies and Haematopota spp. Sci. Rep. 2019, 9, 20076. [Google Scholar] [CrossRef]
- Issimov, A.; Kutumbetov, L.; Orynbayev, M.B.; Khairullin, B.; Myrzakhmetova, B.; Sultankulova, K.; White, P.J. Mechanical Transmission of Lumpy Skin Disease Virus by Stomoxys spp. (Stomoxys calsitrans, Stomoxys sitiens, Stomoxys indica), Diptera: Muscidae. Anim. Open. Access. J. 2020, 10, 477. [Google Scholar] [CrossRef] [PubMed]
- Baldacchino, F.; Muenworn, V.; Desquesnes, M.; Desoli, F.; Charoenviriyaphap, T.; Duvallet, G. Transmission of pathogens by Stomoxys flies (Diptera, Muscidae): A review. Parasite 2013, 20, 26. [Google Scholar] [CrossRef]
- Haegeman, A.; De Leeuw, I.; Mostin, L.; Van Campe, W.; Aerts, L.; Venter, E.; Tuppurainen, E.; Saegerman, C.; De Clercq, K. Comparative Evaluation of Lumpy Skin Disease Virus-Based Live Attenuated Vaccines. Vaccines 2021, 9, 473. [Google Scholar] [CrossRef] [PubMed]
- Carn, V.M.; Kitching, R.P. An investigation of possible routes of transmission of lumpy skin disease virus (Neethling). Epidemiol. Infect. 1995, 114, 219–226. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.; Venter, E.H.; Coetzer, J.A. The detection of lumpy skin disease virus in samples of experimentally infected cattle using different diagnostic techniques. Onderstepoort J. Veter. Res. 2005, 72, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.; Dietze, K.; Wolff, J.; Bergmann, H.; Beltran-Alcrudo, D.; Fahrion, A.; Lamien, C.E.; Busch, F.; Sauter-Louis, C.; Conraths, F.J.; et al. Review: Vaccines and Vaccination against Lumpy Skin Disease. Vaccines 2021, 9, 1136. [Google Scholar] [CrossRef] [PubMed]
- Babiuk, S.; Bowden, T.R.; Parkyn, G.; Dalman, B.; Manning, L.; Neufeld, J.; Embury-Hyatt, C.; Copps, J.; Boyle, D.B. Quantification of lumpy skin disease virus following experimental infection in cattle. Transbound. Emerg. Dis. 2008, 55, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Schumann, H.; Zumpt, F. The Stomoxyine Biting Flies of the World (Diptera: Muscidae).—175 S., 97 Abb., 19 S. Literatur, Stuttgart 1973: Gustav Fischer Verlag. DM 98,-. Dtsch. Entomol. Z. 1973, 20, 433. [Google Scholar] [CrossRef]
- Aerts, L.; Haegeman, A.; De Leeuw, I.; Philips, W.; Van Campe, W.; Behaeghel, I.; Mostin, L.; De Clercq, K. Detection of Clinical and Subclinical Lumpy Skin Disease Using Ear Notch Testing and Skin Biopsies. Microorganisms 2021, 9, 2171. [Google Scholar] [CrossRef]
- Haegeman, A.; Zro, K.; Vandenbussche, F.; Demeestere, L.; Van Campe, W.; Ennaji, M.; De Clercq, K. Development and validation of three Capripoxvirus real-time PCRs for parallel testing. J. Virol. Methods 2013, 193, 446–451. [Google Scholar] [CrossRef]
- Haegeman, A.; De Leeuw, I.; Mostin, L.; Van Campe, W.; Aerts, L.; Vastag, M.; De Clercq, K. An Immunoperoxidase Monolayer Assay (IPMA) for the detection of lumpy skin disease antibodies. J. Virol. Methods 2020, 277, 113800. [Google Scholar] [CrossRef] [PubMed]
- WOAH—World Organisation for Animal Health. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. 2018. Available online: https://www.woah.org/en/produit/manual-of-diagnostic-tests-and-vaccines-for-terrestrial-animals-2018/ (accessed on 12 January 2023).
- Haegeman, A.; Zro, K.; Sammin, D.; Vandenbussche, F.; Ennaji, M.M.; De Clercq, K. Investigation of a Possible Link Between Vaccination and the 2010 Sheep Pox Epizootic in Morocco. Transbound. Emerg. Dis. 2016, 63, e278–e287. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.M.; Venter, E.H.; Coetzer, J.A.W.; Bell-Sakyi, L. Lumpy skin disease: Attempted propagation in tick cell lines and presence of viral DNA in field ticks collected from naturally-infected cattle. Ticks Tick Borne Dis. 2015, 6, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Hameed, M.; Wahaab, A.; Nawaz, M.; Khan, S.; Nazir, J.; Liu, K.; Wei, J.; Ma, Z. Potential Role of Birds in Japanese Encephalitis Virus Zoonotic Transmission and Genotype Shift. Viruses 2021, 13, 357. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.L.; Wang, Q.; Yang, X.Y.; Li, D.L.; Zhao, B.; Ge, G.Y.; Zong, Y.; Li, J.M.; Leng, X.; Shi, K.; et al. Seroprevalence and Risk Factors of the Bluetongue Virus in Cattle in China From 1988 to 2019: A Comprehensive Literature Review and Meta-Analysis. Front. Vet. Sci. 2021, 7. Available online: https://www.frontiersin.org/articles/10.3389/fvets.2020.550381 (accessed on 12 January 2023). [CrossRef]
- Food and Agriculture Organisation of the United Nations. Sustainable prevention, control and elimination of Lumpy Skin Disease—Eastern Europe and the Balkans. FAO Anim. Prod. Health Position Pap. 2017, 2, 25. [Google Scholar]
- Kumar, P.; Kumari, R.R.; Devi, S.; Tripathi, M.K.; Singh, J.; Kumar, R.; Kumar, M. Emergence and transboundary spread of lumpy skin disease in South Asia. Indian J. Anim. Sci. 2021, 91, 507–517. [Google Scholar] [CrossRef]
- EFSA Panel on Animal Health and Welfare (AHAW); Nielsen, S.S.; Alvarez, J.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Rojas, J.L.G.; Schmidt, C.G.; et al. Assessment of the control measures for category A diseases of Animal Health Law: Lumpy Skin Disease. EFSA J. 2022, 20, e07121. [Google Scholar]
- Gubbins, S.; Stegeman, A.; Klement, E.; Pite, L.; Broglia, A.; Cortiñas Abrahantes, J. Inferences about the transmission of lumpy skin disease virus between herds from outbreaks in Albania in 2016. Prev. Veter. Med. 2020, 181, 104602. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | dpi | Donor Animals | Acceptor Animals | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D1 | D2 | D3 | D4 | D5 | D6 | D7 | D8 | D9 | D10 | D11 | D12 | D13 | A1 | A2 | A3 | A4 | A5 | A6 | A7 | A8 | A9 | A10 | A11 | A12 | A13 | ||
| 600 | 7–8–9–10 | 600 | 200 | 200 | 200 | ||||||||||||||||||||||
| 300 | 8–9–10 | 300 | 100 | 100 | 100 | ||||||||||||||||||||||
| 300 | 7–8 | 300 | 100 | 100 | 100 | ||||||||||||||||||||||
| 600 | 8 | 600 | 200 | 200 | 200 | ||||||||||||||||||||||
| 380 | 10 | 380 | 160 | 60 | 160 | ||||||||||||||||||||||
| 200 | 15–16–17 | 200 | 100 | 100 | |||||||||||||||||||||||
| 300 | 15–16–17 | 300 | 100 | 100 | 100 | ||||||||||||||||||||||
| 180 | 22–23–24 | 180 | 90 | 90 | |||||||||||||||||||||||
| 270 | 22–23–24 | 270 | 90 | 90 | 90 | ||||||||||||||||||||||
| 300 | 27–28–29 | 300 | 100 | 100 | 100 | ||||||||||||||||||||||
| 200 | 27–28–29 | 200 | 100 | 100 | |||||||||||||||||||||||
| 3630 | |||||||||||||||||||||||||||
| ID Donor Animal | Peak Ct in Blood (dpi) | Viremic Period (dpi) | Noduli (dpi): FIRST Appearance (Generalization) | Ct Nodules on 9/10 dpi | IPMA | Clinical Diagnosis |
|---|---|---|---|---|---|---|
| D1 | 31.80 (9) | 6 until 23 † | 7 (10) | 18.32 | pos | clinical |
| D2 | 37.48 (6) | 6 and 13 | none | / | pos | subclinical |
| D3 | 31.56 (14) | 6 until 23 † | 6 (9) | 31.99 | pos | clinical |
| D4 | 41.99 (14) | 14 | none | / | pos | subclinical |
| D5 | / | / | none | / | neg | non-infected |
| D6 | 29.75 (13) | 6 until 13 † | 9 (10) | 21.8 | pos | clinical |
| D7 | 28.61 (13) | 6 until 16 † | 10 (10) | 13.45 | pos | clinical |
| D8 | 32.02 (10) | 6 until 13 † | 7 (9) | 14.72 | pos | clinical |
| D9 | 30.06 (14) | 7 until 23 † | 6 (7) | 13.98 | pos | clinical |
| D10 | 32.96 (14) | 6 until 23 † | 11 (13) | 19.59 | pos | clinical |
| D11 | 31.80 (7) | 6 until 10 † | 6 (/) | † | neg | clinical |
| D12 | 40.20 (7) | 7 | none | / | pos | subclinical |
| D13 | 30.23 (14) | 6 until 23 † | 6 (9) | 14.82 | pos | clinical |
| D1 | D2 | D3 | D4 | D5 | D6 | D7 | D8 | D9 | D10 | D11 | D12 | D13 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1/50 | 1/300 | 1/50 | 1/300 | 1/50 | 1/300 | 1/50 | 1/300 | 1/50 | 1/300 | 1/50 | 1/300 | 1/50 | 1/300 | 1/50 | 1/300 | 1/50 | 1/300 | 1/50 | 1/300 | 1/50 | 1/300 | 1/50 | 1/300 | 1/50 | 1/300 | |
| 0 dpi | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N |
| 2 dpi | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N |
| 6 dpi | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N |
| 7 dpi | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N |
| 8 dpi | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N |
| 9 dpi | P | N | N | N | N | N | N | N | N | N | P | N | N | N | N | N | N | N | N | N | N | N | P | N | P | neg |
| 10 dpi | P | P | P | N | N | N | N | N | N | N | P | N | N | N | N | N | N | N | N | N | N | N | P | P | P | P |
| 13 dpi | P | P | P | N | P | N | P | N | N | N | P | P | P | P | P | N | P | N | P | N | † | † | P | P | P | P |
| 14 dpi | P | P | P | N | P | P | P | N | † | † | † | † | P | P | † | † | P | N | P | P | † | † | P | P | P | P |
| 16 dpi | P | P | P | P | P | P | P | N | † | † | † | † | P | P | † | † | P | N | P | P | † | † | P | P | P | P |
| 23 dpi | P | P | P | P | P | P | P | N | † | † | † | † | † | † | † | † | P | P | P | P | † | † | P | P | P | P |
| 28 dpi | † | † | P | P | P | P | P | N | † | † | † | † | † | † | † | † | † | † | † | † | † | † | † | † | † | † |
| 30 dpi | † | † | P | P | † | † | P | N | † | † | † | † | † | † | † | † | † | † | † | † | † | † | † | † | † | † |
| 36 dpi | † | † | P | P | † | † | P | N | † | † | † | † | † | † | † | † | † | † | † | † | † | † | † | † | † | † |
| P | Strong Positive | P | Positive | P | Weak Positive | N | Negative | † | Euthanasia | |||||||||||||||||
| ID Acceptor Animal | Fed on Donor Animal | First Day Flies on (dpi) | Incubation Period: Min-Max (Days) * | Peak Ct in Blood (dpf) | Viraemic Period (dpf) | Nodules Present | IPMA | ELISA | Clinical Diagnosis |
|---|---|---|---|---|---|---|---|---|---|
| A1 | preclinical D10 | 10 dpi | / | / | / | no | neg | neg | non-infected |
| A2 | preclinical D7 | 7 dpi | / | / | / | no | neg | neg | non-infected |
| A3 | preclinical D7 | 7 dpi | / | / | / | no | neg | neg | non-infected |
| A4 | preclinical D7 | 7 dpi | / | / | / | no | neg | neg | non-infected |
| A5 | preclinical D10 | 7 dpi | / | / | / | no | neg | neg | non-infected |
| A6 | preclinical D10 | 7 dpi | / | / | / | no | neg | neg | non-infected |
| A7 | preclinical D10 | 10 dpi | / | / | / | no | neg | neg | non-infected |
| A8 | preclinical D10 | 7 dpi | / | / | / | no | neg | neg | non-infected |
| A9 | subclinical D2 | 15 dpi | / | / | / | no | pos | neg | undetermined |
| A10 | subclinical D2 | 15 dpi | / | / | / | no | neg | neg | non-infected |
| A11 | subclinical D2–D4 | 15 dpi | / | / | / | no | neg | neg | non-infected |
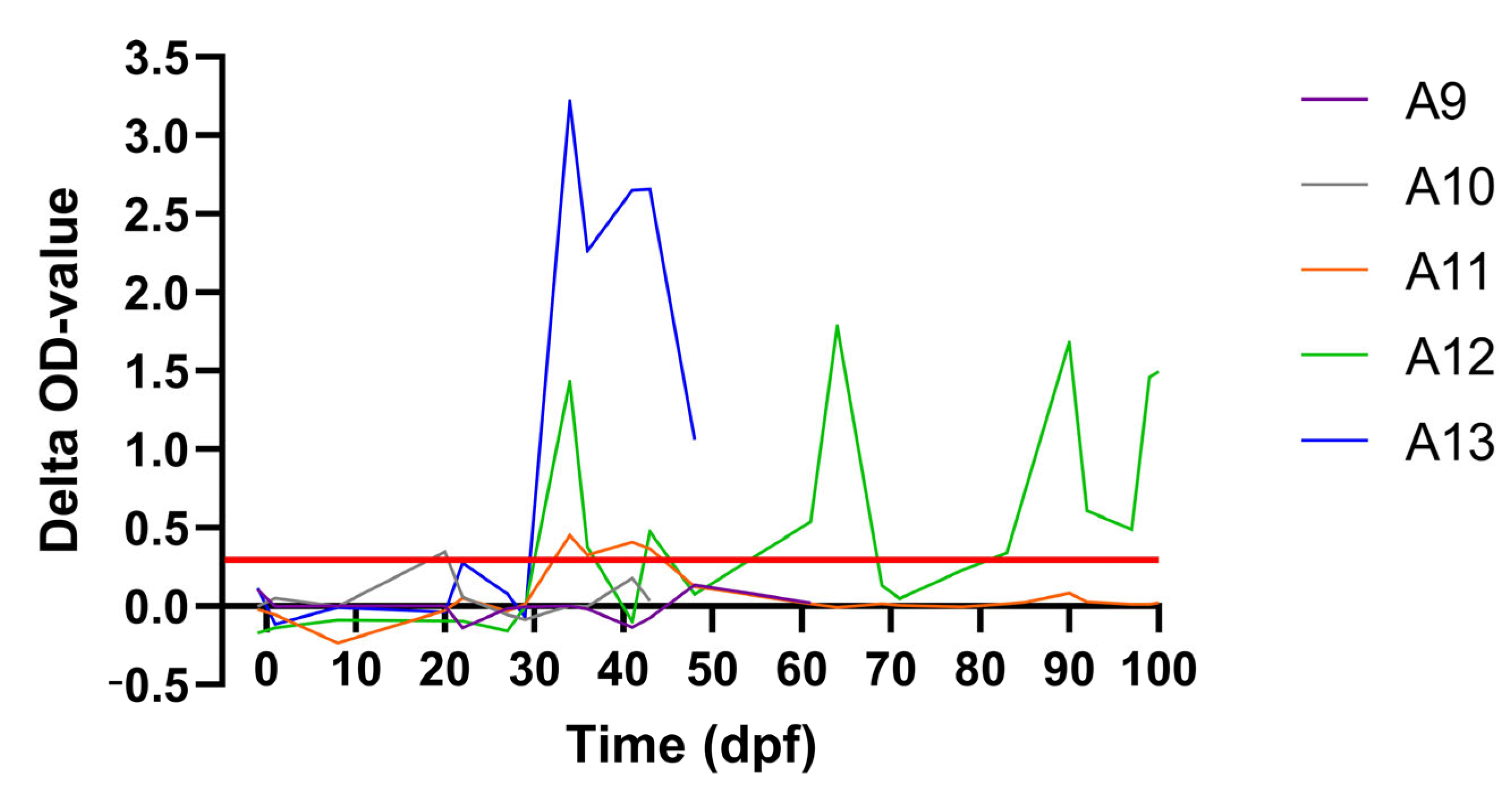
| A12 | subclinical D4 | 15 dpi | 13–27 | 34.44 (50) | 27, 43 until 57 | no | pos | pos | subclinical |
| A13 | subclinical D4 | 15 dpi | 6–20 | 31.59 (22) | 20 until 36 | yes | pos | pos | clinical |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haegeman, A.; Sohier, C.; Mostin, L.; De Leeuw, I.; Van Campe, W.; Philips, W.; De Regge, N.; De Clercq, K. Evidence of Lumpy Skin Disease Virus Transmission from Subclinically Infected Cattle by Stomoxys calcitrans. Viruses 2023, 15, 1285. https://doi.org/10.3390/v15061285
Haegeman A, Sohier C, Mostin L, De Leeuw I, Van Campe W, Philips W, De Regge N, De Clercq K. Evidence of Lumpy Skin Disease Virus Transmission from Subclinically Infected Cattle by Stomoxys calcitrans. Viruses. 2023; 15(6):1285. https://doi.org/10.3390/v15061285
Chicago/Turabian StyleHaegeman, Andy, Charlotte Sohier, Laurent Mostin, Ilse De Leeuw, Willem Van Campe, Wannes Philips, Nick De Regge, and Kris De Clercq. 2023. "Evidence of Lumpy Skin Disease Virus Transmission from Subclinically Infected Cattle by Stomoxys calcitrans" Viruses 15, no. 6: 1285. https://doi.org/10.3390/v15061285