
Rapid Detection of SARS-CoV-2 RNA Using Reverse Transcription Recombinase Polymerase Amplification (RT-RPA) with Lateral Flow for N-Protein Gene and Variant-Specific Deletion–Insertion Mutation in S-Protein Gene
 , , , , , , , ,
, , , , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Primer and Probe Design
2.1.1. Nucleocapsid Phosphoprotein Gene
2.1.2. Omicron BA.1 (del211/ins214), Spike Glycoprotein Gene
2.2. Isothermal Amplification Using RT-RPA
2.2.1. SARS-CoV-2 Partial Nucleocapsid Gene
2.2.2. Omicron BA.1 (del211/ins214)
2.3. Detection of Amplicons Using Lateral Flow Strips
2.4. Limit of Detection and Cross-Reactivity
2.5. Clinical Samples
2.6. RNA Extraction
2.7. Diagnostic Evaluation of RT-RPA-LF in Clinical Samples
2.7.1. SARS-CoV-2 (N) RT-RPA-LF
2.7.2. Omicron BA.1 (S) RT-RPA-LF
2.7.3. Test Accuracy Calculations
2.8. Mutation Frequency of del211/ins214 in the Tested Clinical Samples
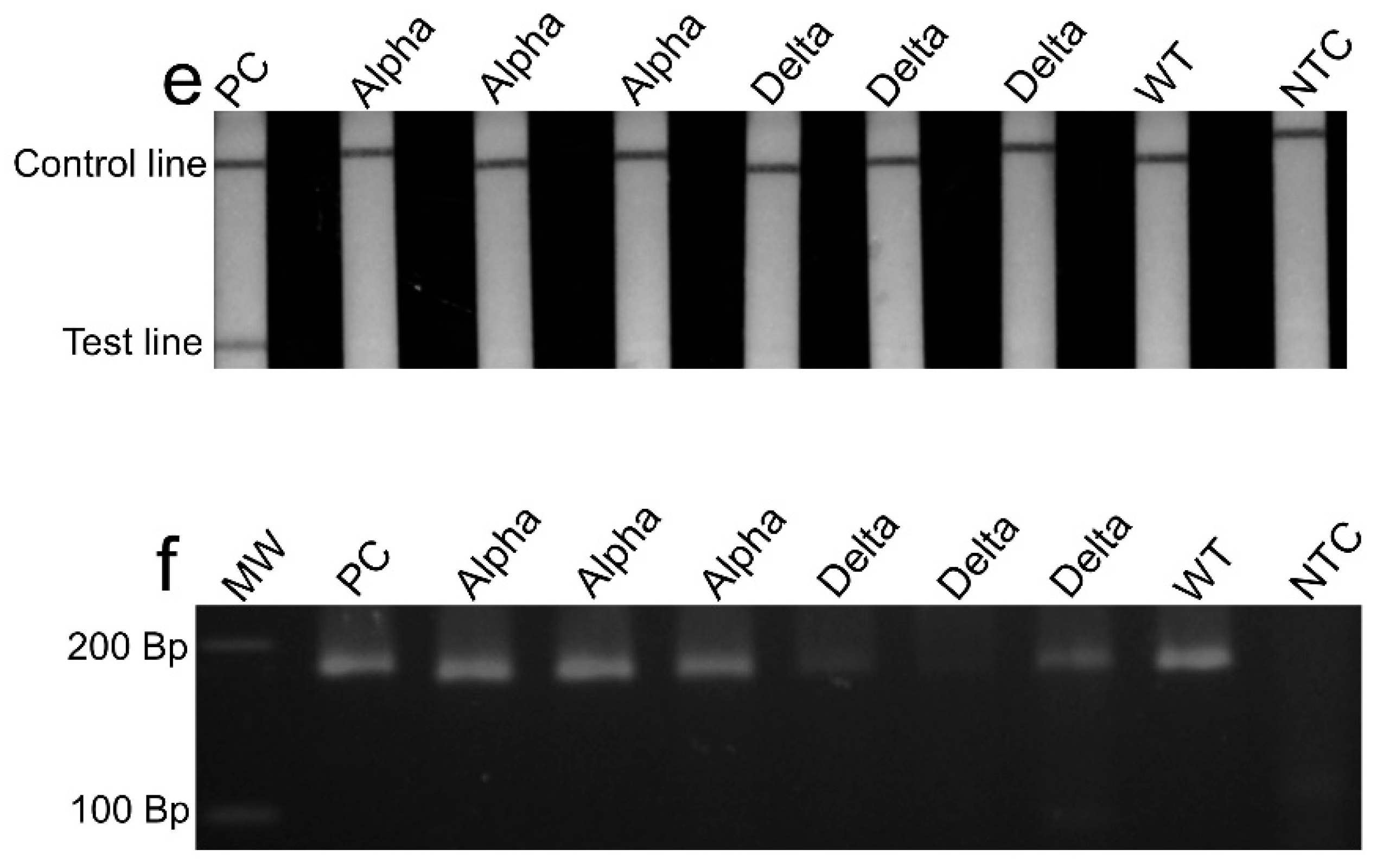
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Primer and Probe Screening of Omicron BA.1
Appendix B. Synthesis of RNA Standards
Appendix B.1. SARS-CoV-2 Partial Nucleocapsid Gene
Appendix B.2. SARS-CoV-2 Partial Spike Gene
Appendix C. Generation of Controls for Omicron BA.1
Appendix C.1. Generation of Positive Control for Omicron BA.1
Appendix C.2. Generation of Negative Control for Omicron BA.1
References
- Callaway, E. Heavily Mutated Omicron Variant Puts Scientists on Alert. Nature 2021, 600, 21. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Bhattacharya, M.; Nag, S.; Dhama, K.; Chakraborty, C. A Detailed Overview of SARS-CoV-2 Omicron: Its Sub-Variants, Mutations and Pathophysiology, Clinical Characteristics, Immunological Landscape, Immune Escape, and Therapies. Viruses 2023, 15, 167. [Google Scholar] [CrossRef] [PubMed]
- Weekly Epidemiological Update on COVID-19—11 May 2023. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19---11-may-2023 (accessed on 12 May 2023).
- Dhawan, M.; Saied, A.R.A.; Mitra, S.; Alhumaydhi, F.A.; Emran, T.B.; Wilairatana, P. Omicron Variant (B.1.1.529) and Its Sublineages: What Do We Know so Far amid the Emergence of Recombinant Variants of SARS-CoV-2? In Biomedicine and Pharmacotherapy; Elsevier Masson s.r.l.: Issy-les-Moulineaux, France, 2022. [Google Scholar] [CrossRef]
- Cao, Y.; Jian, F.; Wang, J.; Yu, Y.; Song, W.; Yisimayi, A.; Wang, J.; An, R.; Chen, X.; Zhang, N.; et al. Imprinted SARS-CoV-2 Humoral Immunity Induces Convergent Omicron RBD Evolution. Nature 2023, 614, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Khatri, R.; Siddqui, G.; Sadhu, S.; Maithil, V.; Vishwakarma, P.; Lohiya, B.; Goswami, A.; Ahmed, S.; Awasthi, A.; Samal, S. Intrinsic D614G and P681R/H Mutations in SARS-CoV-2 VoCs Alpha, Delta, Omicron and Viruses with D614G plus Key Signature Mutations in Spike Protein Alters Fusogenicity and Infectivity. Med. Microbiol. Immunol. 2023, 212, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Uriu, K.; Ito, J.; Zahradnik, J.; Fujita, S.; Kosugi, Y.; Schreiber, G.; Sato, K. Enhanced Transmissibility, Infectivity, and Immune Resistance of the SARS-CoV-2 Omicron XBB.1.5 Variant. In The Lancet Infectious Diseases; Elsevier Ltd.: Amsterdam, The Netherlands, 2023; pp. 280–281. [Google Scholar] [CrossRef]
- Zappa, M.; Verdecchia, P.; Angeli, F. Is the Competition between Variants the End of Severe Acute Respiratory Syndrome Coronavirus 2 Pandemic? A Journey from Wuhan to XBB.1.16. Eur. J. Intern. Med. 2023. [Google Scholar] [CrossRef]
- Kevadiya, B.D.; Machhi, J.; Herskovitz, J.; Oleynikov, M.D.; Blomberg, W.R.; Bajwa, N.; Soni, D.; Das, S.; Hasan, M.; Patel, M.; et al. Diagnostics for SARS-CoV-2 Infections. Nat. Mater. 2021, 20, 593–605. [Google Scholar] [CrossRef]
- Kwok, H.F. The Significance of Advanced COVID-19 Diagnostic Testing in Pandemic Control Measures. Int. J. Biol. Sci. 2022, 18, 4610–4617. [Google Scholar] [CrossRef]
- Nasereddin, A.; Golan Berman, H.; Wolf, D.G.; Oiknine-Djian, E.; Adar, S. Identification of SARS-CoV-2 Variants of Concern Using Amplicon Next-Generation Sequencing. Microbiol. Spectr. 2022, 10, e00736-22. [Google Scholar] [CrossRef]
- Focosi, D.; Quiroga, R.; McConnell, S.; Johnson, M.C.; Casadevall, A. Convergent Evolution in SARS-CoV-2 Spike Creates a Variant Soup from Which New COVID-19 Waves Emerge. Int. J. Mol. Sci. 2023, 24, 2264. [Google Scholar] [CrossRef]
- Brito, A.F.; Semenova, E.; Dudas, G.; Hassler, G.W.; Kalinich, C.C.; Kraemer, M.U.G.; Ho, J.; Tegally, H.; Githinji, G.; Agoti, C.N.; et al. Global Disparities in SARS-CoV-2 Genomic Surveillance. Nat. Commun. 2022, 13, 7003. [Google Scholar] [CrossRef]
- Ong, D.S.Y.; Koeleman, J.G.M.; Vaessen, N.; Breijer, S.; Paltansing, S.; de Man, P. Rapid Screening Method for the Detection of SARS-CoV-2 Variants of Concern. J. Clin. Virol. 2021, 141, 104903. [Google Scholar] [CrossRef] [PubMed]
- Huanyu, W.; Sophonie, J.; Richard, E.; John, M.; Pamela, S.; Huolin, T.; Jones, M.D.; Leber, A.L. Mutation-Specific SARS-CoV-2 PCR Screen: Rapid and Accurate Detection of Variants of Concern and the Identification of a Newly Emerging Variant with Spike L452R Mutation. J. Clin. Microbiol. 2021, 59, e00926-21. [Google Scholar] [CrossRef]
- Chung, H.-Y.; Jian, M.; Chang, C.-K.; Chen, C.-S.; Li, S.-Y.; Lin, J.-C.; Yeh, K.-M.; Yang, Y.-S.; Chen, C.-W.; Hsieh, S.-S.; et al. The Application of a Novel 5-in-1 Multiplex Reverse Transcriptase–Polymerase Chain Reaction Assay for Rapid Detection of SARS-CoV-2 and Differentiation between Variants of Concern. Int. J. Infect. Dis. 2023, 127, 56–62. [Google Scholar] [CrossRef]
- Frediani, J.K.; Levy, J.M.; Rao, A.; Bassit, L.; Figueroa, J.; Vos, M.B.; Wood, A.; Jerris, R.; Leung-Pineda, V.; Gonzalez, M.D.; et al. Multidisciplinary Assessment of the Abbott BinaxNOW SARS-CoV-2 Point-of-Care Antigen Test in the Context of Emerging Viral Variants and Self-Administration. Sci. Rep. 2021, 11, 14604. [Google Scholar] [CrossRef] [PubMed]
- Yamayoshi, S.; Sakai-Tagawa, Y.; Koga, M.; Akasaka, O.; Nakachi, I.; Koh, H.; Maeda, K.; Adachi, E.; Saito, M.; Nagai, H.; et al. Comparison of Rapid Antigen Tests for COVID-19. Viruses 2020, 12, 1420. [Google Scholar] [CrossRef] [PubMed]
- García-Bernalt Diego, J.; Fernández-Soto, P.; Muro, A. The Future of Point-of-Care Nucleic Acid Amplification Diagnostics after COVID-19: Time to Walk the Walk. Int. J. Mol. Sci. 2022, 23, 14110. [Google Scholar] [CrossRef]
- Alhamid, G.; Tombuloglu, H.; Rabaan, A.A.; Al-Suhaimi, E. SARS-CoV-2 Detection Methods: A Comprehensive Review. Saudi J. Biol. Sci. 2022, 29, 103465. [Google Scholar] [CrossRef]
- Piepenburg, O.; Williams, C.H.; Stemple, D.L.; Armes, N.A. DNA Detection Using Recombination Proteins. PLoS Biol. 2006, 4, 1115–1121. [Google Scholar] [CrossRef]
- Zaghloul, H.; El-Shahat, M. Recombinase Polymerase Amplification as a Promising Tool in Hepatitis C Virus Diagnosis. World J. Hepatol. 2014, 6, 916–922. [Google Scholar] [CrossRef]
- Yang, J.; Hu, X.; Wang, W.; Yang, Y.; Zhang, X.; Fang, W.; Zhang, L.; Li, S.; Gu, B. RT-LAMP Assay for Rapid Detection of the R203M Mutation in SARS-CoV-2 Delta Variant. Emerg. Microbes Infect. 2022, 11, 978–987. [Google Scholar] [CrossRef]
- Tang, G.; Zhang, Z.; Tan, W.; Long, F.; Sun, J.; Li, Y.; Zou, S.; Yang, Y.; Cai, K.; Li, S.; et al. RT-RPA-Cas12a-Based Assay Facilitates the Discrimination of SARS-CoV-2 Variants of Concern. Sens. Actuators B Chem. 2023, 381, 133433. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Yang, L.; Zhang, X.; Tang, Y.; Wang, Y.; Shao, X.; Gao, S.; Liu, X.; Wang, P. Rapid and Sensitive Genotyping of SARS-CoV-2 Key Mutation L452R with an RPA-PfAgo Method. Anal. Chem. 2022, 94, 17151–17159. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A Tool to Design Target-Specific Primers for Polymerase Chain Reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- TwistAmp® DNA Amplification Kits Assay Design Manual. Available online: https://www.twistdx.co.uk/wp-content/uploads/2021/04/twistamp-assay-design-manual-v2-5.pdf (accessed on 6 April 2023).
- Elbe, S.; Buckland-Merrett, G. Data, Disease and Diplomacy: GISAID’s Innovative Contribution to Global Health. Glob. Chall. 2017, 1, 33–46. [Google Scholar] [CrossRef]
- O’Toole, Á.; Scher, E.; Underwood, A.; Jackson, B.; Hill, V.; McCrone, J.T.; Colquhoun, R.; Ruis, C.; Abu-Dahab, K.; Taylor, B.; et al. Assignment of Epidemiological Lineages in an Emerging Pandemic Using the Pangolin Tool. Virus Evol. 2021, 7, veab064. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- MGHD. Universal Lateral Flow Dipstick for the Detection of Biotin-and FITC-Labeled Analytes (Proteins, Genomic Amplificates); REF. Available online: https://www.milenia-biotec.com/uploads/2019/07/MGHD_C.pdf (accessed on 14 April 2023).
- CDC 2019-Novel Coronavirus (2019-NCoV) Real-Time RT-PCR Diagnostic Panel For Emergency Use Only Instructions for Use. Available online: https://www.fda.gov/media/134922/download (accessed on 10 April 2023).
- Shirato, K.; Nao, N.; Katano, H.; Takayama, I.; Saito, S.; Kato, F.; Katoh, H.; Sakata, M.; Nakatsu, Y.; Mori, Y.; et al. Development of Genetic Diagnostic Methods for Detection for Novel Coronavirus 2019(NCoV-2019) in Japan. Jpn. J. Infect. Dis. 2020, 73, 304–307. [Google Scholar] [CrossRef]
- Cherkaoui, D.; Heaney, J.; Huang, D.; Byott, M.; Miller, B.S.; Nastouli, E.; McKendry, R.A. Clinical Validation of a Rapid Variant-Proof RT-RPA Assay for the Detection of SARS-CoV-2. Diagnostics 2022, 12, 1263. [Google Scholar] [CrossRef]
- El Wahed, A.A.; Patel, P.; Maier, M.; Pietsch, C.; Rüster, D.; Böhlken-Fascher, S.; Kissenkötter, J.; Behrmann, O.; Frimpong, M.; Diagne, M.M.; et al. Suitcase Lab for Rapid Detection of SARS-CoV-2 Based on Recombinase Polymerase Amplification Assay. Anal. Chem. 2021, 93, 2627–2634. [Google Scholar] [CrossRef]
- Liu, D.; Shen, H.; Zhang, Y.; Shen, D.; Zhu, M.; Song, Y.; Zhu, Z.; Yang, C. A Microfluidic-Integrated Lateral Flow Recombinase Polymerase Amplification (MI-IF-RPA) Assay for Rapid COVID-19 Detection. Lab. Chip 2021, 21, 2019–2026. [Google Scholar] [CrossRef]
- Loan Dao Thi, V.; Herbst, K.; Boerner, K.; Meurer, M.; Kremer, L.P.; Kirrmaier, D.; Freistaedter, A.; Papagiannidis, D.; Galmozzi, C.; Stanifer, M.L.; et al. A Colorimetric RT-LAMP Assay and LAMP-Sequencing for Detecting SARS-CoV-2 RNA in Clinical Samples. Sci. Transl. Med. 2020, 12, eabc7075. [Google Scholar] [CrossRef] [PubMed]
- Sherrill-Mix, S.; Hwang, Y.; Roche, A.M.; Glascock, A.; Weiss, S.R.; Li, Y.; Haddad, L.; Deraska, P.; Monahan, C.; Kromer, A.; et al. Detection of SARS-CoV-2 RNA Using RT-LAMP and Molecular Beacons. Genome Biol. 2021, 22, 169. [Google Scholar] [CrossRef] [PubMed]
- Muenchhoff, M.; Mairhofer, H.; Nitschko, H.; Grzimek-Koschewa, N.; Hoffmann, D.; Berger, A.; Rabenau, H.; Widera, M.; Ackermann, N.; Konrad, R.; et al. Multicentre Comparison of Quantitative PCR-Based Assays to Detect SARS-CoV-2, Germany, March 2020. Eurosurveillance 2020, 25, 2001057. [Google Scholar] [CrossRef]
- Sun, Y.; Yu, L.; Liu, C.; Ye, S.; Chen, W.; Li, D.; Huang, W. One-Tube SARS-CoV-2 Detection Platform Based on RT-RPA and CRISPR/Cas12a. J. Transl. Med. 2021, 19, 74. [Google Scholar] [CrossRef]
- Lau, Y.L.; Ismail, I.B.; Mustapa, N.I.B.; Lai, M.Y.; Soh, T.S.T.; Hassan, A.H.; Peariasamy, K.M.; Lee, Y.L.; Kahar, M.K.B.A.; Chong, J.; et al. Development of a Reverse Transcription Recombinase Polymerase Amplification Assay for Rapid and Direct Visual Detection of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). PLoS ONE 2021, 16, e0245164. [Google Scholar] [CrossRef]
- Ghosh, P.; Chowdhury, R.; Hossain, M.E.; Hossain, F.; Miah, M.; Rashid, M.U.; Baker, J.; Rahman, M.Z.; Rahman, M.; Ma, X.; et al. Evaluation of Recombinase-Based Isothermal Amplification Assays for Point-of-Need Detection of SARS-CoV-2 in Resource-Limited Settings. Int. J. Infect. Dis. 2022, 114, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Shelite, T.R.; Uscanga-Palomeque, A.C.; Castellanos-Gonzalez, A.; Melby, P.C.; Travi, B.L. Isothermal Recombinase Polymerase Amplification-Lateral Flow Detection of SARS-CoV-2, the Etiological Agent of COVID-19. J. Virol. Methods 2021, 296, 114227. [Google Scholar] [CrossRef]
- Aoki, M.N.; de Oliveira Coelho, B.; Góes, L.G.B.; Minoprio, P.; Durigon, E.L.; Morello, L.G.; Marchini, F.K.; Riediger, I.N.; do Carmo Debur, M.; Nakaya, H.I.; et al. Colorimetric RT-LAMP SARS-CoV-2 Diagnostic Sensitivity Relies on Color Interpretation and Viral Load. Sci. Rep. 2021, 11, 9026. [Google Scholar] [CrossRef]
- Mayuramart, O.; Nimsamer, P.; Rattanaburi, S.; Chantaravisoot, N.; Khongnomnan, K.; Chansaenroj, J.; Puenpa, J.; Suntronwong, N.; Vichaiwattana, P.; Poovorawan, Y.; et al. Detection of Severe Acute Respiratory Syndrome Coronavirus 2 and Influenza Viruses Based on CRISPR-Cas12a. Exp. Biol. Med. 2021, 246, 400–405. [Google Scholar] [CrossRef]
- Aman, R.; Marsic, T.; Sivakrishna Rao, G.; Mahas, A.; Ali, Z.; Alsanea, M.; Al-Qahtani, A.; Alhamlan, F.; Mahfouz, M. ISCAN-V2: A One-Pot RT-RPA–CRISPR/Cas12b Assay for Point-of-Care SARS-CoV-2 Detection. Front. Bioeng. Biotechnol. 2022, 9, 800104. [Google Scholar] [CrossRef]
- Bruner, K.M.; Wang, Z.; Simonetti, F.R.; Bender, A.M.; Kwon, K.J.; Sengupta, S.; Fray, E.J.; Beg, S.A.; Antar, A.A.R.; Jenike, K.M.; et al. A Quantitative Approach for Measuring the Reservoir of Latent HIV-1 Proviruses. Nature 2019, 566, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, K.; Kakumu, S.; Wakita, T.; Ishikawa, T.; Itoh, Y.; Takayanagi, M.; Higashi, Y.; Shibata, M.; Morishima, T. Detection of Hepatitis C Virus by Polymerase Chain Reaction and Response to Interferon-α Therapy: Relationship to Genotypes of Hepatitis C Virus. Hepatology 1992, 16, 293–299. [Google Scholar] [CrossRef]
- Savino, S.; Desmet, T.; Franceus, J. Insertions and Deletions in Protein Evolution and Engineering. Biotechnol. Adv. 2022, 60, 108010. [Google Scholar] [CrossRef] [PubMed]
- Cantoni, D.; Murray, M.J.; Kalemera, M.D.; Dicken, S.J.; Stejskal, L.; Brown, G.; Lytras, S.; Coey, J.D.; McKenna, J.; Bridgett, S.; et al. Evolutionary Remodelling of N-terminal Domain Loops Fine-tunes SARS-CoV-2 Spike. EMBO Rep. 2022, 23, e54322. [Google Scholar] [CrossRef]
- Menéndez-Arias, L.; Matamoros, T.; Cases-González, C.E. Insertions and Deletions in HIV-1 Reverse Transcriptase: Consequences for Drug Resistance and Viral Fitness. Curr. Pharm. Des. 2006, 12, 1811–1825. [Google Scholar] [CrossRef] [PubMed]
- Migueres, M.; Lhomme, S.; Trémeaux, P.; Dimeglio, C.; Ranger, N.; Latour, J.; Dubois, M.; Nicot, F.; Miedouge, M.; Mansuy, J.M.; et al. Evaluation of Two RT-PCR Screening Assays for Identifying SARS-CoV-2 Variants. J. Clin. Virol. 2021, 143, 104969. [Google Scholar] [CrossRef] [PubMed]
- Ao, D.; He, X.; Hong, W.; Wei, X. The Rapid Rise of SARS-CoV-2 Omicron Subvariants with Immune Evasion Properties: XBB.1.5 and BQ.1.1 Subvariants. MedComm 2023, 4, e239. [Google Scholar] [CrossRef] [PubMed]
- Babu, B.; Ochoa-Corona, F.M.; Paret, M.L. Recombinase Polymerase Amplification Applied to Plant Virus Detection and Potential Implications. Anal. Biochem. 2018, 546, 72–77. [Google Scholar] [CrossRef]
- Prediger, E. Calculations: Converting from Nanograms to Copy Number. Available online: https://sg.idtdna.com/pages/education/decoded/article/calculations-converting-from-nanograms-to-copy-number (accessed on 5 April 2023).








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malaga, J.L.; Pajuelo, M.J.; Okamoto, M.; Tsinda, E.K.; Otani, K.; Tsukayama, P.; Mascaro, L.; Cuicapuza, D.; Katsumi, M.; Kawamura, K.; et al. Rapid Detection of SARS-CoV-2 RNA Using Reverse Transcription Recombinase Polymerase Amplification (RT-RPA) with Lateral Flow for N-Protein Gene and Variant-Specific Deletion–Insertion Mutation in S-Protein Gene. Viruses 2023, 15, 1254. https://doi.org/10.3390/v15061254
Malaga JL, Pajuelo MJ, Okamoto M, Tsinda EK, Otani K, Tsukayama P, Mascaro L, Cuicapuza D, Katsumi M, Kawamura K, et al. Rapid Detection of SARS-CoV-2 RNA Using Reverse Transcription Recombinase Polymerase Amplification (RT-RPA) with Lateral Flow for N-Protein Gene and Variant-Specific Deletion–Insertion Mutation in S-Protein Gene. Viruses. 2023; 15(6):1254. https://doi.org/10.3390/v15061254
Chicago/Turabian StyleMalaga, Jose L., Monica J. Pajuelo, Michiko Okamoto, Emmanuel Kagning Tsinda, Kanako Otani, Pablo Tsukayama, Lucero Mascaro, Diego Cuicapuza, Masamichi Katsumi, Kazuhisa Kawamura, and et al. 2023. "Rapid Detection of SARS-CoV-2 RNA Using Reverse Transcription Recombinase Polymerase Amplification (RT-RPA) with Lateral Flow for N-Protein Gene and Variant-Specific Deletion–Insertion Mutation in S-Protein Gene" Viruses 15, no. 6: 1254. https://doi.org/10.3390/v15061254