
Gastrointestinal Manifestations of SARS-CoV-2: Transmission, Pathogenesis, Immunomodulation, Microflora Dysbiosis, and Clinical Implications
 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Routes of Transmission of SARS-CoV-2
3. Digestive Manifestations and Pathophysiology of SARS-CoV-2 Infection
4. Pathogenesis of SARS-CoV-2 in the GI Tract
5. Role of Receptors in SARS-CoV-2 Pathogenesis
6. Immune Response of the Host to SARS-CoV-2 Infection
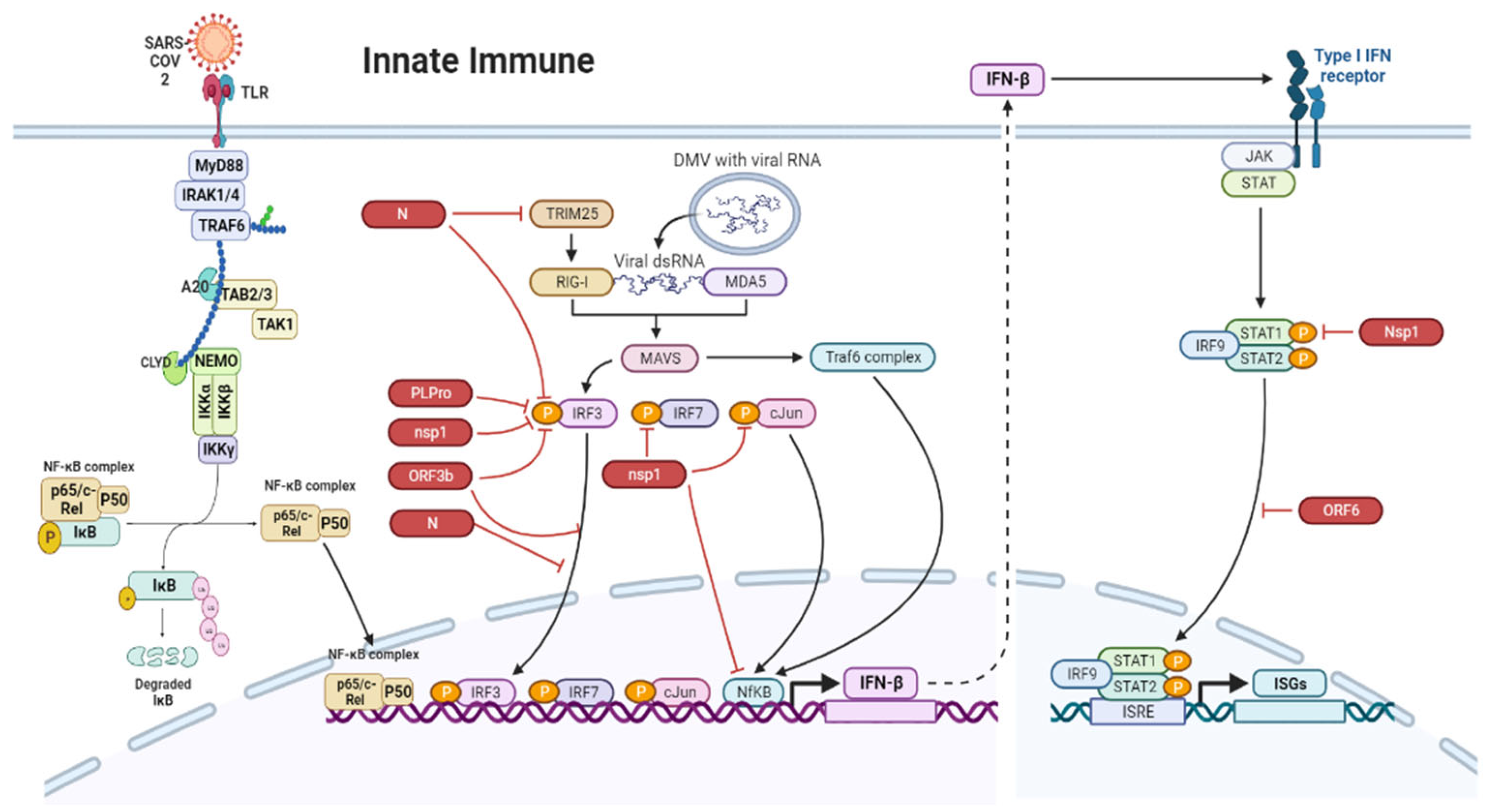
7. Innate Immune Responses in the Gut during SARS-CoV-2 Infection
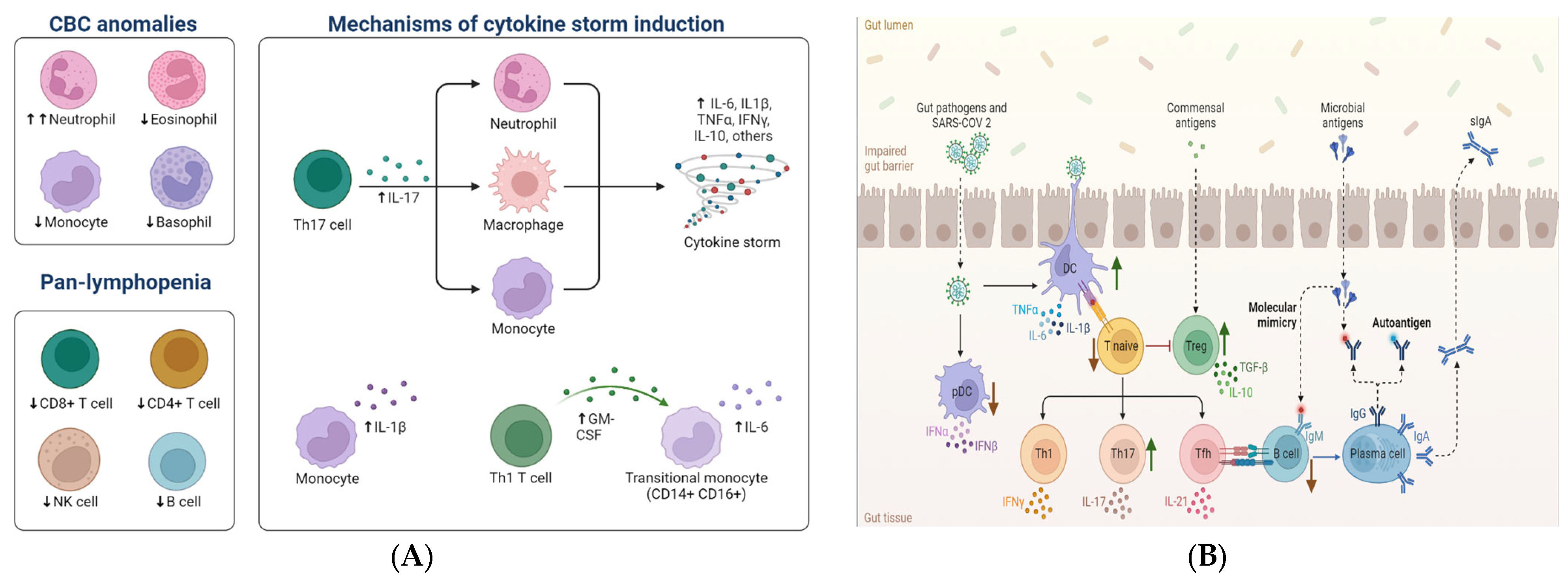
8. Adaptive Immune Response
9. COVID-19 and Gut Microbiota
10. Gut-Lung Axis in COVID-19
11. Therapeutics
12. Limitations of the Study
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Corman, V.M.; Muth, D.; Niemeyer, D.; Drosten, C. Hosts and Sources of Endemic Human Coronaviruses. Adv. Virus Res. 2018, 100, 163–188. [Google Scholar] [PubMed]
- Stanifer, M.L.; Kee, C.; Cortese, M.; Triana, S.; Mukenhirn, M.; Kraeusslich, H.-G.; Alexandrov, T.; Bartenschlager, R.; Boulant, S. Critical Role of Type III Interferon in Controlling SARS-CoV-2 Infection, Replication and Spread in Primary Human Intestinal Epithelial Cells. BioRxiv 2020. [Google Scholar] [CrossRef]
- Neurath, M.F. COVID-19 and Immunomodulation in IBD. Gut 2020, 69, 1335–1342. [Google Scholar] [CrossRef] [PubMed]
- Villapol, S. Gastrointestinal Symptoms Associated with COVID-19: Impact on the Gut Microbiome. Transl. Res. 2020, 226, 57–69. [Google Scholar] [CrossRef]
- Luo, X.; Zhou, G.-Z.; Zhang, Y.; Peng, L.-H.; Zou, L.-P.; Yang, Y.-S. Coronaviruses and Gastrointestinal Diseases. Mil. Med. Res. 2020, 7, 49. [Google Scholar] [CrossRef]
- Hung, I.F.N.; Cheng, V.C.C.; Wu, A.K.L.; Tang, B.S.F.; Chan, K.H.; Chu, C.M.; Wong, M.M.L.; Hui, W.T.; Poon, L.L.M.; Tse, D.M.W. Viral Loads in Clinical Specimens and SARS Manifestations. Emerg. Infect. Dis. 2004, 10, 1550. [Google Scholar] [CrossRef]
- Leung, W.K.; To, K.; Chan, P.K.; Chan, H.L.; Wu, A.K.; Lee, N.; Yuen, K.Y.; Sung, J.J. Enteric Involvement of Severe Acute Respiratory Syndrome-Associated Coronavirus Infection. Gastroenterology 2003, 125, 1011–1017. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and Evolution of Pathogenic Coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef]
- Livanos, A.E.; Jha, D.; Cossarini, F.; Gonzalez-Reiche, A.S.; Tokuyama, M.; Aydillo, T.; Parigi, T.L.; Ramos, I.; Dunleavy, K.; Lee, B. Gastrointestinal Involvement Attenuates COVID-19 Severity and Mortality. MedRxiv 2020. [Google Scholar] [CrossRef]
- Ng, S.C.; Tilg, H. COVID-19 and the Gastrointestinal Tract: More than Meets the Eye. Gut 2020, 69, 973–974. [Google Scholar] [CrossRef]
- Kanwar, A.; Selvaraju, S.; Esper, F. Human Coronavirus-HKU1 Infection among Adults in Cleveland, Ohio. In Open Forum Infectious Diseases; Oxford University Press: Oxford, UK, 2017; Volume 4. [Google Scholar]
- Dominguez, S.R.; Robinson, C.C.; Holmes, K.V. Detection of Four Human Coronaviruses in Respiratory Infections in Children: A One-Year Study in Colorado. J. Med. Virol. 2009, 81, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Assiri, A.; McGeer, A.; Perl, T.M.; Price, C.S.; Al Rabeeah, A.A.; Cummings, D.A.; Alabdullatif, Z.N.; Assad, M.; Almulhim, A.; Makhdoom, H. Hospital Outbreak of Middle East Respiratory Syndrome Coronavirus. N. Engl. J. Med. 2013, 369, 407–416. [Google Scholar] [CrossRef] [PubMed]
- D’amico, F.; Baumgart, D.C.; Danese, S.; Peyrin-Biroulet, L. Diarrhea during COVID-19 Infection: Pathogenesis, Epidemiology, Prevention, and Management. Clin. Gastroenterol. Hepatol. 2020, 18, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Harmer, D.; Gilbert, M.; Borman, R.; Clark, K.L. Quantitative MRNA Expression Profiling of ACE 2, a Novel Homologue of Angiotensin Converting Enzyme. FEBS Lett. 2002, 532, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.; Royce, S.G.; Tikellis, C.; Shallue, C.; Batu, D.; Velkoska, E.; Burrell, L.M.; Patel, S.K.; Beswick, L.; Jackson, A. Imbalance of the Renin–Angiotensin System May Contribute to Inflammation and Fibrosis in IBD: A Novel Therapeutic Target? Gut 2020, 69, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, R.; Mainali, R.; Ahmadi, S.; Wang, S.; Singh, R.; Kavanagh, K.; Kitzman, D.W.; Kushugulova, A.; Marotta, F.; Yadav, H. Gut Microbiome and Aging: Physiological and Mechanistic Insights. Nutr. Healthy Aging 2018, 4, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Roussos, A.; Koursarakos, P.; Patsopoulos, D.; Gerogianni, I.; Philippou, N. Increased Prevalence of Irritable Bowel Syndrome in Patients with Bronchial Asthma. Respir. Med. 2003, 97, 75–79. [Google Scholar] [CrossRef]
- Rutten, E.P.; Lenaerts, K.; Buurman, W.A.; Wouters, E.F. Disturbed Intestinal Integrity in Patients with COPD: Effects of Activities of Daily Living. Chest 2014, 145, 245–252. [Google Scholar] [CrossRef]
- Dijkman, R.; Jebbink, M.F.; El Idrissi, N.B.; Pyrc, K.; Müller, M.A.; Kuijpers, T.W.; Zaaijer, H.L.; Van Der Hoek, L. Human Coronavirus NL63 and 229E Seroconversion in Children. J. Clin. Microbiol. 2008, 46, 2368–2373. [Google Scholar] [CrossRef]
- Esper, F.; Ou, Z.; Huang, Y.T. Human Coronaviruses Are Uncommon in Patients with Gastrointestinal Illness. J. Clin. Virol. 2010, 48, 131–133. [Google Scholar] [CrossRef]
- Reusken, C.B.; Farag, E.A.; Jonges, M.; Godeke, G.-J.; El-Sayed, A.M.; Pas, S.D.; Raj, V.S.; Mohran, K.A.; Moussa, H.A.; Ghobashy, H. Middle East Respiratory Syndrome Coronavirus (MERS-CoV) RNA and Neutralising Antibodies in Milk Collected According to Local Customs from Dromedary Camels, Qatar, April 2014. Eurosurveillance 2014, 19, 20829. [Google Scholar] [CrossRef] [PubMed]
- Cimolai, N. Features of Enteric Disease from Human Coronaviruses: Implications for COVID-19. J. Med. Virol. 2020, 92, 1834–1844. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Liang, T.J. Is SARS-CoV-2 Also an Enteric Pathogen with Potential Fecal–Oral Transmission? A COVID-19 Virological and Clinical Review. Gastroenterology 2020, 159, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Risku, M.; Lappalainen, S.; Räsänen, S.; Vesikari, T. Detection of Human Coronaviruses in Children with Acute Gastroenteritis. J. Clin. Virol. 2010, 48, 27–30. [Google Scholar] [CrossRef]
- Ning, T.; Liu, S.; Xu, J.; Yang, Y.; Zhang, N.; Xie, S.; Min, L.; Zhang, S.; Zhu, S.; Wang, Y. Potential Intestinal Infection and Faecal-Oral Transmission of Human Coronaviruses. Rev. Med. Virol. 2022, 32, e2363. [Google Scholar] [CrossRef]
- Isakbaeva, E.T.; Khetsuriani, N.; Beard, R.S.; Peck, A.; Erdman, D.; Monroe, S.S.; Tong, S.; Ksiazek, T.G.; Lowther, S.; Smith, I.P. SARS-Associated Coronavirus Transmission, United States. Emerg. Infect. Dis. 2004, 10, 225. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, C.; Zhao, G.; Chu, H.; Wang, D.; Yan, H.H.-N.; Poon, V.K.-M.; Wen, L.; Wong, B.H.-Y.; Zhao, X. Human Intestinal Tract Serves as an Alternative Infection Route for Middle East Respiratory Syndrome Coronavirus. Sci. Adv. 2017, 3, eaao4966. [Google Scholar] [CrossRef]
- Chan, P.K.; To, W.-K.; Ng, K.-C.; Lam, R.K.; Ng, T.-K.; Chan, R.C.; Wu, A.; Yu, W.-C.; Lee, N.; Hui, D.S. Laboratory Diagnosis of SARS. Emerg. Infect. Dis. 2004, 10, 825. [Google Scholar] [CrossRef]
- Xu, Y.; Li, X.; Zhu, B.; Liang, H.; Fang, C.; Gong, Y.; Guo, Q.; Sun, X.; Zhao, D.; Shen, J. Characteristics of Pediatric SARS-CoV-2 Infection and Potential Evidence for Persistent Fecal Viral Shedding. Nat. Med. 2020, 26, 502–505. [Google Scholar] [CrossRef]
- Zhang, J.; Garrett, S.; Sun, J. Gastrointestinal Symptoms, Pathophysiology, and Treatment in COVID-19. Genes Dis. 2021, 8, 385–400. [Google Scholar] [CrossRef]
- Talbot, H.K.B.; Crowe, J.E., Jr.; Edwards, K.M.; Griffin, M.R.; Zhu, Y.; Weinberg, G.A.; Szilagyi, P.G.; Hall, C.B.; Podsiad, A.B.; Iwane, M. Coronavirus Infection and Hospitalizations for Acute Respiratory Illness in Young Children. J. Med. Virol. 2009, 81, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Sonkar, C.; Kashyap, D.; Varshney, N.; Baral, B.; Jha, H.C. Impact of Gastrointestinal Symptoms in COVID-19: A Molecular Approach. SN Compr. Clin. Med. 2020, 2, 2658–2669. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Tang, M.; Zheng, X.; Liu, Y.; Li, X.; Shan, H. Evidence for Gastrointestinal Infection of SARS-CoV-2. Gastroenterology 2020, 158, 1831–1833. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tang, C.; Tian, D.; Hou, X.; Yang, Y. Management of Digestive Disorders and Procedures Associated with COVID-19. Am. J. Gastroenterol. 2020, 115, 1153–1155. [Google Scholar] [CrossRef]
- Wu, Y.; Guo, C.; Tang, L.; Hong, Z.; Zhou, J.; Dong, X.; Yin, H.; Xiao, Q.; Tang, Y.; Qu, X. Prolonged Presence of SARS-CoV-2 Viral RNA in Faecal Samples. Lancet Gastroenterol. Hepatol. 2020, 5, 434–435. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Wang, B.; Zhang, T.; Xu, J.; Shang, S. The Mechanism and Treatment of Gastrointestinal Symptoms in Patients with COVID-19. Am. J. Physiol.-Gastrointest. Liver Physiol. 2020, 319, G245–G252. [Google Scholar] [CrossRef]
- Zhang, D.; Li, S.; Wang, N.; Tan, H.-Y.; Zhang, Z.; Feng, Y. The Cross-Talk between Gut Microbiota and Lungs in Common Lung Diseases. Front. Microbiol. 2020, 11, 301. [Google Scholar] [CrossRef]
- Delgado-Gonzalez, P.; Gonzalez-Villarreal, C.A.; Roacho-Perez, J.A.; Quiroz-Reyes, A.G.; Islas, J.F.; Delgado-Gallegos, J.L.; Arellanos-Soto, D.; Galan-Huerta, K.A.; Garza-Treviño, E.N. Inflammatory Effect on the Gastrointestinal System Associated with COVID-19. World J. Gastroenterol. 2021, 27, 4160. [Google Scholar] [CrossRef]
- Devaux, C.A.; Lagier, J.-C.; Raoult, D. New Insights into the Physiopathology of COVID-19: SARS-CoV-2-Associated Gastrointestinal Illness. Front. Med. 2021, 8, 640073. [Google Scholar] [CrossRef]
- Tian, Y.; Rong, L.; Nian, W.; He, Y. Gastrointestinal Features in COVID-19 and the Possibility of Faecal Transmission. Aliment. Pharmacol. Ther. 2020, 51, 843–851. [Google Scholar] [CrossRef]
- Cooney, J.; Appiahene, P.; Findlay, R.; Al-Hillawi, L.; Rafique, K.; Laband, W.; Shandro, B.; Poullis, A. COVID-19 Infection Causing Residual Gastrointestinal Symptoms–a Single UK Centre Case Series. Clin. Med. 2022, 22, 181. [Google Scholar] [CrossRef] [PubMed]
- Bogariu, A.M.; Dumitrascu, D.L. Digestive Involvement in the Long-COVID Syndrome. Med. Pharm. Rep. 2022, 95, 5. [Google Scholar] [CrossRef] [PubMed]
- Baig, A.M. Chronic COVID Syndrome: Need for an Appropriate Medical Terminology for Long-COVID and COVID Long-Haulers. J. Med. Virol. 2021, 93, 2555–2556. [Google Scholar] [CrossRef] [PubMed]
- Halpin, S.; O’Connor, R.; Sivan, M. Long COVID and Chronic COVID Syndromes. J. Med. Virol. 2021, 93, 1242. [Google Scholar] [CrossRef]
- Weng, J.; Li, Y.; Li, J.; Shen, L.; Zhu, L.; Liang, Y.; Lin, X.; Jiao, N.; Cheng, S.; Huang, Y. Gastrointestinal Sequelae 90 Days after Discharge for COVID-19. Lancet Gastroenterol. Hepatol. 2021, 6, 344–346. [Google Scholar] [CrossRef]
- Effenberger, M.; Grabherr, F.; Mayr, L.; Schwaerzler, J.; Nairz, M.; Seifert, M.; Hilbe, R.; Seiwald, S.; Scholl-Buergi, S.; Fritsche, G. Faecal Calprotectin Indicates Intestinal Inflammation in COVID-19. Gut 2020, 69, 1543–1544. [Google Scholar] [CrossRef]
- Cortes, G.M.; Marcialis, M.A.; Bardanzellu, F.; Corrias, A.; Fanos, V.; Mussap, M. Inflammatory Bowel Disease and COVID-19: How Microbiomics and Metabolomics Depict Two Sides of the Same Coin. Front. Microbiol. 2022, 13, 856165. [Google Scholar] [CrossRef]
- Satsangi, J.; Silverberg, M.S.; Vermeire, S.; Colombel, J.F. The Montreal Classification of Inflammatory Bowel Disease: Controversies, Consensus, and Implications. Gut 2006, 55, 749–753. [Google Scholar] [CrossRef]
- Sultan, K.; Mone, A.; Durbin, L.; Khuwaja, S.; Swaminath, A. Review of Inflammatory Bowel Disease and COVID-19. World J. Gastroenterol. 2020, 26, 5534. [Google Scholar] [CrossRef]
- Monteleone, G.; Ardizzone, S. Are Patients with Inflammatory Bowel Disease at Increased Risk for Covid-19 Infection? J. Crohn’s Colitis 2020, 14, 1334–1336. [Google Scholar] [CrossRef]
- Dipasquale, V.; Cucchiara, S.; Martinelli, M.; Miele, E.; Aloi, M.; Romano, C. Challenges in Paediatric Inflammatory Bowel Diseases in the COVID-19 Time. Dig. Liver Dis. 2020, 52, 593–594. [Google Scholar] [CrossRef] [PubMed]
- Bourgonje, A.R.; van Linschoten, R.C.; West, R.L.; van Dijk, M.A.; van Leer-Buter, C.C.; Kats-Ugurlu, G.; Pierik, M.J.; Festen, E.A.; Weersma, R.K.; Dijkstra, G. Treatment of Severe Acute Ulcerative Colitis in SARS-CoV-2 Infected Patients: Report of Three Cases and Discussion of Treatment Options. Ther. Adv. Gastroenterol. 2021, 14, 17562848211012596. [Google Scholar] [CrossRef]
- Imperatore, N.; Bennato, R.; D’Avino, A.; Lombardi, G.; Manguso, F. SARS-CoV-2 as a Trigger for de Novo Ulcerative Colitis. Inflamm. Bowel Dis. 2021, 27, e87–e88. [Google Scholar] [CrossRef] [PubMed]
- Rubin, D.T.; Abreu, M.T.; Rai, V.; Siegel, C.A.; Ahuja, V.; Allez, M.; Ananthakrishnan, A.N.; Bernstein, C.N.; Braun, J.G.; Chowers, Y. Management of Patients with Crohn’s Disease and Ulcerative Colitis during the Coronavirus Disease-2019 Pandemic: Results of an International Meeting. Gastroenterology 2020, 159, 6–13. [Google Scholar] [CrossRef]
- Abdulla, M.; Mohammed, N.; AlQamish, J.; Mosli, M. Inflammatory Bowel Disease and COVID-19 Outcomes: A Meta-Analysis. Sci. Rep. 2022, 12, 21333. [Google Scholar] [CrossRef] [PubMed]
- Abrams, J.Y.; Godfred-Cato, S.E.; Oster, M.E.; Chow, E.J.; Koumans, E.H.; Bryant, B.; Leung, J.W.; Belay, E.D. Multisystem Inflammatory Syndrome in Children Associated with Severe Acute Respiratory Syndrome Coronavirus 2: A Systematic Review. J. Pediatr. 2020, 226, 45–54. [Google Scholar] [CrossRef]
- Yonker, L.M.; Gilboa, T.; Ogata, A.F.; Senussi, Y.; Lazarovits, R.; Boribong, B.P.; Bartsch, Y.C.; Loiselle, M.; Rivas, M.N.; Porritt, R.A. Multisystem Inflammatory Syndrome in Children Is Driven by Zonulin-Dependent Loss of Gut Mucosal Barrier. J. Clin. Investig. 2021, 131, e149633. [Google Scholar] [CrossRef]
- Camargo, S.M.; Singer, D.; Makrides, V.; Huggel, K.; Pos, K.M.; Wagner, C.A.; Kuba, K.; Danilczyk, U.; Skovby, F.; Kleta, R. Tissue-Specific Amino Acid Transporter Partners ACE2 and Collectrin Differentially Interact with Hartnup Mutations. Gastroenterology 2009, 136, 872–882. [Google Scholar] [CrossRef]
- Hashimoto, T.; Perlot, T.; Rehman, A.; Trichereau, J.; Ishiguro, H.; Paolino, M.; Sigl, V.; Hanada, T.; Hanada, R.; Lipinski, S. ACE2 Links Amino Acid Malnutrition to Microbial Ecology and Intestinal Inflammation. Nature 2012, 487, 477–481. [Google Scholar] [CrossRef]
- Zhang, H.; Kang, Z.; Gong, H.; Xu, D.; Wang, J.; Li, Z.; Li, Z.; Cui, X.; Xiao, J.; Zhan, J. Digestive System Is a Potential Route of COVID-19: An Analysis of Single-Cell Coexpression Pattern of Key Proteins in Viral Entry Process. Gut 2020, 69, 1010–1018. [Google Scholar] [CrossRef]
- Jakhmola, S.; Indari, O.; Chatterjee, S.; Jha, H.C. SARS-CoV-2, an Underestimated Pathogen of the Nervous System. SN Compr. Clin. Med. 2020, 2, 2137–2146. [Google Scholar] [CrossRef] [PubMed]
- Kalantar-Zadeh, K.; Ward, S.A.; Kalantar-Zadeh, K.; El-Omar, E.M. Considering the Effects of Microbiome and Diet on SARS-CoV-2 Infection: Nanotechnology Roles. ACS Nano 2020, 14, 5179–5182. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, S.; Li, D.; Wei, D.-Q.; Zhao, J.; Wang, J. Human Intestinal Defensin 5 Inhibits SARS-CoV-2 Invasion by Cloaking ACE2. Gastroenterology 2020, 159, 1145–1147. [Google Scholar] [CrossRef] [PubMed]
- Niv, Y. Defensin 5 for Prevention of SARS-CoV-2 Invasion and Covid-19 Disease. Med. Hypotheses 2020, 143, 110244. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Han, B.; Wang, J. COVID-19: Gastrointestinal Manifestations and Potential Fecal–Oral Transmission. Gastroenterology 2020, 158, 1518–1519. [Google Scholar] [CrossRef]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. HLH across Speciality Collaboration, UK: COVID-19: Consider Cytokine Storm Syndromes and Immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Yantiss, R.K.; Qin, L.; He, B.; Crawford, C.V.; Seshan, S.; Patel, S.; Wahid, N.; Jessurun, J. Intestinal Abnormalities in Patients with SARS-CoV-2 Infection: Histopathologic Changes Reflect Mechanisms of Disease. Am. J. Surg. Pathol. 2022, 46, 89–96. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.L.C.; Lely, A.T.; van Navis, G.; van Goor, H. Tissue Distribution of ACE2 Protein, the Functional Receptor for SARS Coronavirus. A First Step in Understanding SARS Pathogenesis. J. Pathol. A J. Pathol. Soc. Great Br. Irel. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Li, H.; Liu, S.-M.; Yu, X.-H.; Tang, S.-L.; Tang, C.-K. Coronavirus Disease 2019 (COVID-19): Current Status and Future Perspectives. Int. J. Antimicrob. Agents 2020, 55, 105951. [Google Scholar] [CrossRef]
- Morniroli, D.; Giannì, M.L.; Consales, A.; Pietrasanta, C.; Mosca, F. Human Sialome and Coronavirus Disease-2019 (COVID-19) Pandemic: An Understated Correlation? Front. Immunol. 2020, 11, 1480. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Bansal, V.; Feschotte, C. A Single-Cell RNA Expression Map of Human Coronavirus Entry Factors. Cell Rep. 2020, 32, 108175. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Kopetz, S.; Vilar, E.; Shen, J.P.; Chen, K.; Maitra, A. Relative Abundance of SARS-CoV-2 Entry Genes in the Enterocytes of the Lower Gastrointestinal Tract. Genes 2020, 11, 645. [Google Scholar] [CrossRef] [PubMed]
- Gheblawi, M.; Wang, K.; Viveiros, A.; Nguyen, Q.; Zhong, J.-C.; Turner, A.J.; Raizada, M.K.; Grant, M.B.; Oudit, G.Y. Angiotensin-Converting Enzyme 2: SARS-CoV-2 Receptor and Regulator of the Renin-Angiotensin System: Celebrating the 20th Anniversary of the Discovery of ACE2. Circ. Res. 2020, 126, 1456–1474. [Google Scholar] [CrossRef] [PubMed]
- Harrison, A.G.; Lin, T.; Wang, P. Mechanisms of SARS-CoV-2 Transmission and Pathogenesis. Trends Immunol. 2020, 41, 1100–1115. [Google Scholar] [CrossRef] [PubMed]
- Kuba, K.; Imai, Y.; Ohto-Nakanishi, T.; Penninger, J.M. Trilogy of ACE2: A Peptidase in the Renin–Angiotensin System, a SARS Receptor, and a Partner for Amino Acid Transporters. Pharmacol. Ther. 2010, 128, 119–128. [Google Scholar] [CrossRef]
- Bayati, A.; Kumar, R.; Francis, V.; McPherson, P.S. SARS-CoV-2 Infects Cells after Viral Entry via Clathrin-Mediated Endocytosis. J. Biol. Chem. 2021, 296, 100306. [Google Scholar] [CrossRef]
- Garg, M.; Burrell, L.M.; Velkoska, E.; Griggs, K.; Angus, P.W.; Gibson, P.R.; Lubel, J.S. Upregulation of Circulating Components of the Alternative Renin-Angiotensin System in Inflammatory Bowel Disease: A Pilot Study. J. Renin-Angiotensin-Aldosterone Syst. 2015, 16, 559–569. [Google Scholar] [CrossRef]
- Tao, S.-S.; Wang, X.-Y.; Yang, X.-K.; Liu, Y.-C.; Fu, Z.-Y.; Zhang, L.-Z.; Wang, Z.-X.; Ni, J.; Shuai, Z.-W.; Pan, H.-F. COVID-19 and Inflammatory Bowel Disease Crosstalk: From Emerging Association to Clinical Proposal. J. Med. Virol. 2022, 94, 5640–5652. [Google Scholar] [CrossRef]
- Nayak, B.; Lal, G.; Kumar, S.; Das, C.J.; Saraya, A. Host Response to SARS-CoV2 and Emerging Variants in Pre-Existing Liver and Gastrointestinal Diseases. Front. Cell. Infect. Microbiol. 2021, 11, 753249. [Google Scholar] [CrossRef] [PubMed]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of Gut Microbiota in Type 2 Diabetes Pathophysiology. EBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.W.; Kitai, T.; Hazen, S.L. Gut Microbiota in Cardiovascular Health and Disease. Circ. Res. 2017, 120, 1183–1196. [Google Scholar] [CrossRef] [PubMed]
- Crowe, C.R.; Chen, K.; Pociask, D.A.; Alcorn, J.F.; Krivich, C.; Enelow, R.I.; Ross, T.M.; Witztum, J.L.; Kolls, J.K. Critical Role of IL-17RA in Immunopathology of Influenza Infection. J. Immunol. 2009, 183, 5301–5310. [Google Scholar] [CrossRef]
- Hörnich, B.F.; Großkopf, A.K.; Schlagowski, S.; Tenbusch, M.; Kleine-Weber, H.; Neipel, F.; Stahl-Hennig, C.; Hahn, A.S. SARS-CoV-2 and SARS-CoV Spike-Mediated Cell-Cell Fusion Differ in Their Requirements for Receptor Expression and Proteolytic Activation. J. Virol. 2021, 95, e00002-21. [Google Scholar] [CrossRef]
- Puoti, M.G.; Rybak, A.; Kiparissi, F.; Gaynor, E.; Borrelli, O. SARS-CoV-2 and the Gastrointestinal Tract in Children. Front. Pediatr. 2021, 9, 617980. [Google Scholar] [CrossRef]
- Triana, S.; Metz-Zumaran, C.; Ramirez, C.; Kee, C.; Doldan, P.; Shahraz, M.; Schraivogel, D.; Gschwind, A.R.; Sharma, A.K.; Steinmetz, L.M. Single-Cell Analyses Reveal SARS-CoV-2 Interference with Intrinsic Immune Response in the Human Gut. Mol. Syst. Biol. 2021, 17, e10232. [Google Scholar] [CrossRef]
- Domínguez-Díaz, C.; García-Orozco, A.; Riera-Leal, A.; Padilla-Arellano, J.R.; Fafutis-Morris, M. Microbiota and Its Role on Viral Evasion: Is It with Us or against Us? Front. Cell. Infect. Microbiol. 2019, 9, 256. [Google Scholar] [CrossRef]
- Budden, K.F.; Gellatly, S.L.; Wood, D.L.; Cooper, M.A.; Morrison, M.; Hugenholtz, P.; Hansbro, P.M. Emerging Pathogenic Links between Microbiota and the Gut–Lung Axis. Nat. Rev. Microbiol. 2017, 15, 55–63. [Google Scholar] [CrossRef]
- Frieman, M.; Baric, R. Mechanisms of Severe Acute Respiratory Syndrome Pathogenesis and Innate Immunomodulation. Microbiol. Mol. Biol. Rev. 2008, 72, 672–685. [Google Scholar] [CrossRef]
- Kamitani, W.; Narayanan, K.; Huang, C.; Lokugamage, K.; Ikegami, T.; Ito, N.; Kubo, H.; Makino, S. Severe Acute Respiratory Syndrome Coronavirus Nsp1 Protein Suppresses Host Gene Expression by Promoting Host MRNA Degradation. Proc. Natl. Acad. Sci. USA 2006, 103, 12885–12890. [Google Scholar] [CrossRef] [PubMed]
- Perlot, T.; Penninger, J.M. ACE2–From the Renin–Angiotensin System to Gut Microbiota and Malnutrition. Microbes Infect. 2013, 15, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Merad, M.; Martin, J.C. Pathological Inflammation in Patients with COVID-19: A Key Role for Monocytes and Macrophages. Nat. Rev. Immunol. 2020, 20, 355–362. [Google Scholar] [CrossRef]
- Baindara, P.; Sarker, M.B.; Earhart, A.P.; Mandal, S.M.; Schrum, A.G. NOTCH Signaling in COVID-19: A Central Hub Controlling Genes, Proteins, and Cells That Mediate SARS-CoV-2 Entry, the Inflammatory Response, and Lung Regeneration. Front. Cell. Infect. Microbiol. 2022, 1100, 928704. [Google Scholar] [CrossRef]
- Shang, Y.; Smith, S.; Hu, X. Role of Notch Signaling in Regulating Innate Immunity and Inflammation in Health and Disease. Protein Cell 2016, 7, 159–174. [Google Scholar] [CrossRef]
- Vázquez-Ulloa, E.; Lizano, M.; Sjöqvist, M.; Olmedo-Nieva, L.; Contreras-Paredes, A. Deregulation of the Notch Pathway as a Common Road in Viral Carcinogenesis. Rev. Med. Virol. 2018, 28, e1988. [Google Scholar] [CrossRef]
- Rizzo, P.; Vieceli Dalla Sega, F.; Fortini, F.; Marracino, L.; Rapezzi, C.; Ferrari, R. COVID-19 in the Heart and the Lungs: Could We “Notch” the Inflammatory Storm? Basic Res. Cardiol. 2020, 115, 31. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 183, 1735. [Google Scholar] [CrossRef]
- Qiu, H.; Tang, X.; Ma, J.; Shaverdashvili, K.; Zhang, K.; Bedogni, B. Notch1 Autoactivation via Transcriptional Regulation of Furin, Which Sustains Notch1 Signaling by Processing Notch1-Activating Proteases ADAM10 and Membrane Type 1 Matrix Metalloproteinase. Mol. Cell. Biol. 2015, 35, 3622–3632. [Google Scholar] [CrossRef]
- Heurich, A.; Hofmann-Winkler, H.; Gierer, S.; Liepold, T.; Jahn, O.; Pöhlmann, S. TMPRSS2 and ADAM17 Cleave ACE2 Differentially and Only Proteolysis by TMPRSS2 Augments Entry Driven by the Severe Acute Respiratory Syndrome Coronavirus Spike Protein. J. Virol. 2014, 88, 1293–1307. [Google Scholar] [CrossRef] [PubMed]
- Doberstein, K.; Steinmeyer, N.; Hartmetz, A.-K.; Eberhardt, W.; Mittelbronn, M.; Harter, P.N.; Juengel, E.; Blaheta, R.; Pfeilschifter, J.; Gutwein, P. MicroRNA-145 Targets the Metalloprotease ADAM17 and Is Suppressed in Renal Cell Carcinoma Patients. Neoplasia 2013, 15, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Bartolomé, A.; Liang, J.; Wang, P.; Ho, D.D.; Pajvani, U.B. Angiotensin Converting Enzyme 2 Is a Novel Target of the γ-Secretase Complex. Sci. Rep. 2021, 11, 9803. [Google Scholar] [CrossRef] [PubMed]
- Magro, G. SARS-CoV-2 and COVID-19: Is Interleukin-6 (IL-6) the “culprit Lesion” of ARDS Onset? What Is There besides Tocilizumab? SGP130Fc. Cytokine X 2020, 2, 100029. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.-Y.; Ma, Y.-T.; Zhang, J.-Y.; Xie, X. COVID-19 and the Cardiovascular System. Nat. Rev. Cardiol. 2020, 17, 259–260. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, P.S.; Wimmers, F.; Mok, C.K.P.; Perera, R.A.P.M.; Scott, M.; Hagan, T.; Sigal, N.; Feng, Y.; Bristow, L.; Tak-Yin Tsang, O.; et al. Systems Biological Assessment of Immunity to Mild versus Severe COVID-19 Infection in Humans. Science 2020, 369, 1210–1220. [Google Scholar] [CrossRef]
- Braun, J.; Loyal, L.; Frentsch, M.; Wendisch, D.; Georg, P.; Kurth, F.; Hippenstiel, S.; Dingeldey, M.; Kruse, B.; Fauchere, F.; et al. SARS-CoV-2-Reactive T Cells in Healthy Donors and Patients with COVID-19. Nature 2020, 587, 270–274. [Google Scholar] [CrossRef]
- Lehmann, M.; Allers, K.; Heldt, C.; Meinhardt, J.; Schmidt, F.; Rodriguez-Sillke, Y.; Kunkel, D.; Schumann, M.; Böttcher, C.; Stahl-Hennig, C.; et al. Human Small Intestinal Infection by SARS-CoV-2 Is Characterized by a Mucosal Infiltration with Activated CD8+ T Cells. Mucosal Immunol. 2021, 14, 1381–1392. [Google Scholar] [CrossRef]
- Gaebler, C.; Wang, Z.; Lorenzi, J.C.C.; Muecksch, F.; Finkin, S.; Tokuyama, M.; Cho, A.; Jankovic, M.; Schaefer-Babajew, D.; Oliveira, T.Y.; et al. Evolution of Antibody Immunity to SARS-CoV-2. Nature 2021, 591, 639–644. [Google Scholar] [CrossRef]
- Feng, Z.; Wang, Y.; Qi, W. The Small Intestine, an Underestimated Site of SARS-CoV-2 Infection: From Red Queen Effect to Probiotics. Preprints.org 2020, 2020030161. [Google Scholar] [CrossRef]
- Stenstad, H.; Ericsson, A.; Johansson-Lindbom, B.; Svensson, M.; Marsal, J.; Mack, M.; Picarella, D.; Soler, D.; Marquez, G.; Briskin, M. Gut-Associated Lymphoid Tissue–Primed CD4+ T Cells Display CCR9-Dependent and-Independent Homing to the Small Intestine. Blood 2006, 107, 3447–3454. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Mackay, C.R. Diet, Gut Microbiota and Immune Responses. Nat. Immunol. 2011, 12, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The Microbiome and Innate Immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Donati Zeppa, S.; Agostini, D.; Piccoli, G.; Stocchi, V.; Sestili, P. Gut Microbiota Status in COVID-19: An Unrecognized Player? Front. Cell. Infect. Microbiol. 2020, 10, 576551. [Google Scholar] [CrossRef]
- Hills, R.D., Jr.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C.; Theberge, C.R. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients 2019, 11, 1613. [Google Scholar] [CrossRef] [PubMed]
- van der Lelie, D.; Taghavi, S. COVID-19 and the Gut Microbiome: More than a Gut Feeling. MSystems 2020, 5, e00453-20. [Google Scholar] [CrossRef]
- Robinson, C.M.; Pfeiffer, J.K. Viruses and the Microbiota. Annu. Rev. Virol. 2014, 1, 55–69. [Google Scholar] [CrossRef]
- Karst, S.M. The Influence of Commensal Bacteria on Infection with Enteric Viruses. Nat. Rev. Microbiol. 2016, 14, 197–204. [Google Scholar] [CrossRef]
- Deriu, E.; Boxx, G.M.; He, X.; Pan, C.; Benavidez, S.D.; Cen, L.; Rozengurt, N.; Shi, W.; Cheng, G. Influenza Virus Affects Intestinal Microbiota and Secondary Salmonella Infection in the Gut through Type I Interferons. PLoS Pathog. 2016, 12, e1005572. [Google Scholar] [CrossRef]
- Zuo, T.; Zhang, F.; Lui, G.C.; Yeoh, Y.K.; Li, A.Y.; Zhan, H.; Wan, Y.; Chung, A.C.; Cheung, C.P.; Chen, N. Alterations in Gut Microbiota of Patients with COVID-19 during Time of Hospitalization. Gastroenterology 2020, 159, 944–955. [Google Scholar] [CrossRef]
- Li, N.; Ma, W.-T.; Pang, M.; Fan, Q.-L.; Hua, J.-L. The Commensal Microbiota and Viral Infection: A Comprehensive Review. Front. Immunol. 2019, 10, 1551. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, Y.; Ichiki, T.; Yamakawa, T.; Tsuji, Y.; Kuronuma, K.; Takahashi, S.; Narimatsu, E.; Nakase, H. Impaired Tryptophan Metabolism in the Gastrointestinal Tract of Patients with Critical Coronavirus Disease 2019. Front. Med. 2022, 9, 941422. [Google Scholar] [CrossRef]
- Sorgdrager, F.J.; Naudé, P.J.; Kema, I.P.; Nollen, E.A.; Deyn, P.P.D. Tryptophan Metabolism in Inflammaging: From Biomarker to Therapeutic Target. Front. Immunol. 2019, 10, 2565. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, A.P.; Lopes, A.L.F.; Pacheco, G.; de Nolêto, I.R.S.G.; Nicolau, L.A.D.; Medeiros, J.V.R. Premises among SARS-CoV-2, Dysbiosis and Diarrhea: Walking through the ACE2/MTOR/Autophagy Route. Med. Hypotheses 2020, 144, 110243. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhao, X.; Patel, A.; Potru, R.; Azizi-Ghannad, S.; Dolinger, M.; Cao, J.; Bartholomew, C.; Mazurkiewicz, J.; Conti, D. Rapamycin Inhibition of MTOR Reduces Levels of the Na+/H+ Exchanger 3 in Intestines of Mice and Humans, Leading to Diarrhea. Gastroenterology 2015, 149, 151–162. [Google Scholar] [CrossRef]
- Cumhur Cure, M.; Cure, E. Effects of the Na+/H+ Ion Exchanger on Susceptibility to COVID-19 and the Course of the Disease. J. Renin-Angiotensin-Aldosterone Syst. 2021, 2021, 4754440. [Google Scholar] [CrossRef]
- He, P.; Yun, C.C. Mechanisms of the Regulation of the Intestinal Exchanger NHE3. J. Biomed. Biotechnol. 2010, 2010, 238080. [Google Scholar] [CrossRef]
- Mohandas, S.; Vairappan, B. Severe Acute Respiratory Syndrome Coronavirus-2 Infection and the Gut–Liver Axis. J. Dig. Dis. 2020, 21, 687–695. [Google Scholar] [CrossRef]
- Dean, M.J.; Ochoa, J.B.; Sanchez-Pino, M.D.; Zabaleta, J.; Garai, J.; Del Valle, L.; Wyczechowska, D.; Baiamonte, L.B.; Philbrook, P.; Majumder, R. Severe COVID-19 Is Characterized by an Impaired Type I Interferon Response and Elevated Levels of Arginase Producing Granulocytic Myeloid Derived Suppressor Cells. Front. Immunol. 2021, 12, 695972. [Google Scholar] [CrossRef]
- Johnson, B.A.; Hage, A.; Kalveram, B.; Mears, M.; Plante, J.A.; Rodriguez, S.E.; Ding, Z.; Luo, X.; Bente, D.; Bradrick, S.S. Peptidoglycan-Associated Cyclic Lipopeptide Disrupts Viral Infectivity. J. Virol. 2019, 93, e01282-19. [Google Scholar] [CrossRef]
- Roy, K.; Agarwal, S.; Banerjee, R.; Paul, M.K.; Purbey, P.K. COVID-19 and Gut Immunomodulation. World J. Gastroenterol. 2021, 27, 7925. [Google Scholar] [CrossRef] [PubMed]
- Viana, S.D.; Nunes, S.; Reis, F. ACE2 Imbalance as a Key Player for the Poor Outcomes in COVID-19 Patients with Age-Related Comorbidities–Role of Gut Microbiota Dysbiosis. Ageing Res. Rev. 2020, 62, 101123. [Google Scholar] [CrossRef] [PubMed]
- Zaim, S.; Chong, J.H.; Sankaranarayanan, V.; Harky, A. COVID-19 and Multiorgan Response. Curr. Probl. Cardiol. 2020, 45, 100618. [Google Scholar] [CrossRef] [PubMed]
- Baindara, P.; Chakraborty, R.; Holliday, Z.M.; Mandal, S.M.; Schrum, A.G. Oral Probiotics in Coronavirus Disease 2019: Connecting the Gut–Lung Axis to Viral Pathogenesis, Inflammation, Secondary Infection and Clinical Trials. New Microbes New Infect. 2021, 40, 100837. [Google Scholar] [CrossRef]
- Jin, X.; Lian, J.-S.; Hu, J.-H.; Gao, J.; Zheng, L.; Zhang, Y.-M.; Hao, S.-R.; Jia, H.-Y.; Cai, H.; Zhang, X.-L. Epidemiological, Clinical and Virological Characteristics of 74 Cases of Coronavirus-Infected Disease 2019 (COVID-19) with Gastrointestinal Symptoms. Gut 2020, 69, 1002–1009. [Google Scholar] [CrossRef]
- Luo, S.; Zhang, X.; Xu, H. Don’t Overlook Digestive Symptoms in Patients with 2019 Novel Coronavirus Disease (COVID-19). Clin. Gastroenterol. Hepatol. 2020, 18, 1636–1637. [Google Scholar] [CrossRef]
- Santacroce, L.; Inchingolo, F.; Topi, S.; Del Prete, R.; Di Cosola, M.; Charitos, I.A.; Montagnani, M. Potential Beneficial Role of Probiotics on the Outcome of COVID-19 Patients: An Evolving Perspective. Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 295–301. [Google Scholar] [CrossRef]
- Angurana, S.K.; Bansal, A. Probiotics and Coronavirus Disease 2019: Think about the Link. Br. J. Nutr. 2021, 126, 1564–1570. [Google Scholar] [CrossRef]
- Sundararaman, A.; Ray, M.; Ravindra, P.V.; Halami, P.M. Role of Probiotics to Combat Viral Infections with Emphasis on COVID-19. Appl. Microbiol. Biotechnol. 2020, 104, 8089–8104. [Google Scholar] [CrossRef]
- Santacroce, L.; Charitos, I.A.; Bottalico, L. A Successful History: Probiotics and Their Potential as Antimicrobials. Expert Rev. Anti-Infect. Ther. 2019, 17, 635–645. [Google Scholar] [CrossRef]
- Shimizu, K.; Hirata, H.; Kabata, D.; Tokuhira, N.; Koide, M.; Ueda, A.; Tachino, J.; Shintani, A.; Uchiyama, A.; Fujino, Y. Ivermectin Administration Is Associated with Lower Gastrointestinal Complications and Greater Ventilator-Free Days in Ventilated Patients with COVID-19: A Propensity Score Analysis. J. Infect. Chemother. 2022, 28, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.L.; Simonetti, B.; Klein, K.; Chen, K.-E.; Williamson, M.K.; Antón-Plágaro, C.; Shoemark, D.K.; Simón-Gracia, L.; Bauer, M.; Hollandi, R. Neuropilin-1 Is a Host Factor for SARS-CoV-2 Infection. Science 2020, 370, 861–865. [Google Scholar] [CrossRef]
- Kow, C.S.; Hasan, S.S. The Use of Antimotility Drugs in COVID-19 Associated Diarrhea. J. Infect. 2021, 82, e19. [Google Scholar] [CrossRef] [PubMed]
- Roberto Poscia, M.D. Oxygen-Ozone as Adjuvant Treatment in Early Control of COVID-19 Progression and Modulation of the Gut Microbial Flora. 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT04366089 (accessed on 17 May 2023).
- Wu, C.; Xu, Q.; Cao, Z.; Pan, D.; Zhu, Y.; Wang, S.; Liu, D.; Song, Z.; Jiang, W.; Ruan, Y. The Volatile and Heterogeneous Gut Microbiota Shifts of COVID-19 Patients over the Course of a Probiotics-Assisted Therapy. Clin. Transl. Med. 2021, 11, e643. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Han, H.; Li, X.; Chen, C.; Xie, X.; Su, G.; Ye, S.; Wang, C.; He, Q.; Wang, F. Probiotics Use Is Associated with Improved Clinical Outcomes among Hospitalized Patients with COVID-19. Ther. Adv. Gastroenterol. 2021, 14, 17562848211035670. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Cheng, F.; Xu, Q.; Su, Y.; Cai, X.; Zeng, F.; Zhang, Y. The Role of Probiotics in Coronavirus Disease-19 Infection in Wuhan: A Retrospective Study of 311 Severe Patients. Int. Immunopharmacol. 2021, 95, 107531. [Google Scholar] [CrossRef]
- Gutiérrez-Castrellón, P.; Gandara-Martí, T.; Abreu, Y.; Abreu, A.T.; Nieto-Rufino, C.D.; López-Orduña, E.; Jiménez-Escobar, I.; Jiménez-Gutiérrez, C.; López-Velazquez, G.; Espadaler-Mazo, J. Probiotic Improves Symptomatic and Viral Clearance in Covid19 Outpatients: A Randomized, Quadruple-Blinded, Placebo-Controlled Trial. Gut Microbes 2022, 14, 2018899. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durairajan, S.S.K.; Singh, A.K.; Saravanan, U.B.; Namachivayam, M.; Radhakrishnan, M.; Huang, J.-D.; Dhodapkar, R.; Zhang, H. Gastrointestinal Manifestations of SARS-CoV-2: Transmission, Pathogenesis, Immunomodulation, Microflora Dysbiosis, and Clinical Implications. Viruses 2023, 15, 1231. https://doi.org/10.3390/v15061231
Durairajan SSK, Singh AK, Saravanan UB, Namachivayam M, Radhakrishnan M, Huang J-D, Dhodapkar R, Zhang H. Gastrointestinal Manifestations of SARS-CoV-2: Transmission, Pathogenesis, Immunomodulation, Microflora Dysbiosis, and Clinical Implications. Viruses. 2023; 15(6):1231. https://doi.org/10.3390/v15061231
Chicago/Turabian StyleDurairajan, Siva Sundara Kumar, Abhay Kumar Singh, Udhaya Bharathy Saravanan, Mayurikaa Namachivayam, Moorthi Radhakrishnan, Jian-Dong Huang, Rahul Dhodapkar, and Hongjie Zhang. 2023. "Gastrointestinal Manifestations of SARS-CoV-2: Transmission, Pathogenesis, Immunomodulation, Microflora Dysbiosis, and Clinical Implications" Viruses 15, no. 6: 1231. https://doi.org/10.3390/v15061231