
Fungal Viruses Unveiled: A Comprehensive Review of Mycoviruses
Abstract
:1. Introduction
2. The History and Origins of Mycoviruses
3. Diversity and Taxonomy
3.1. Fungal Host Range
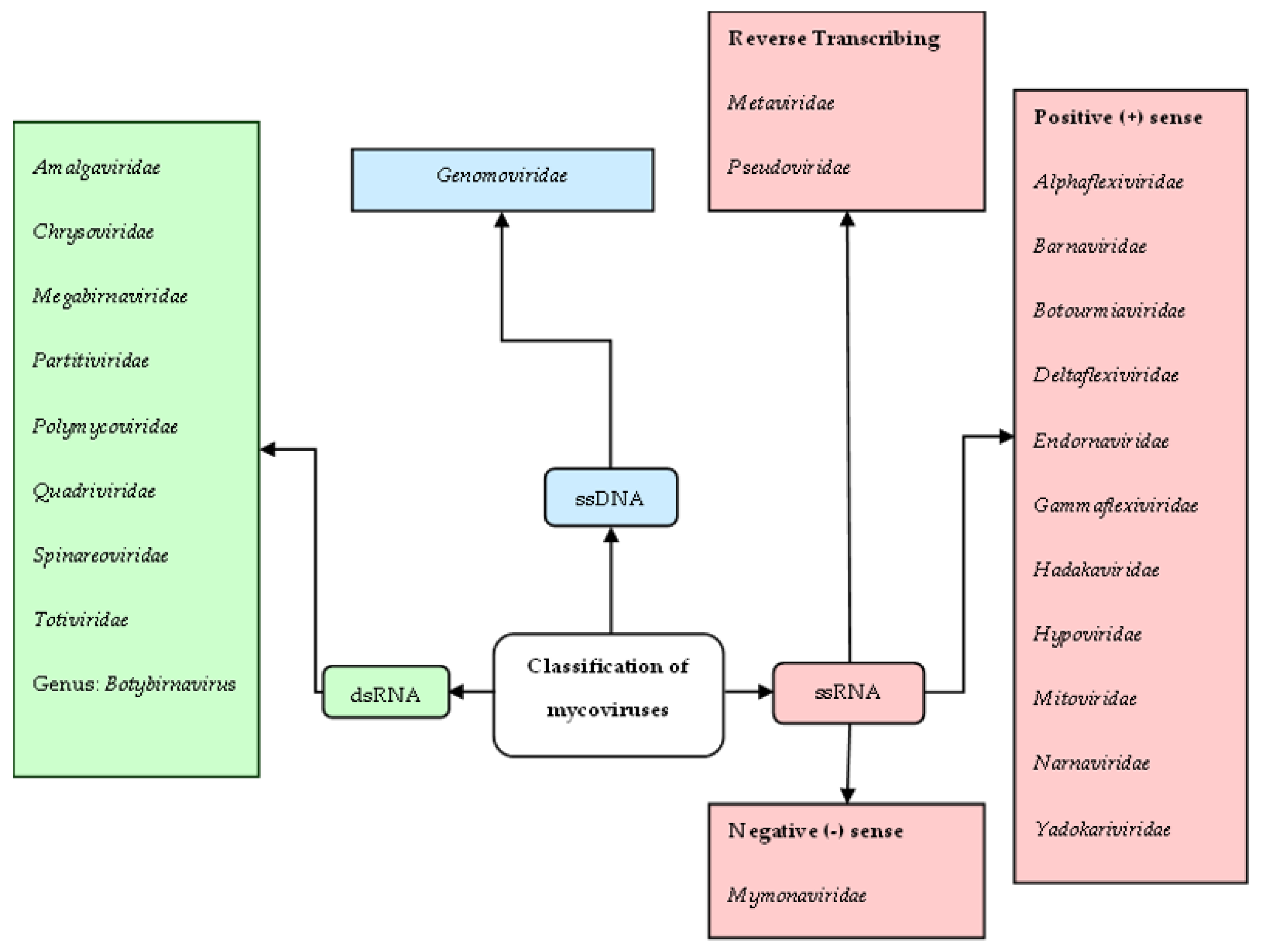
3.2. Mycoviral Taxa
3.2.1. Double-Stranded RNA (dsRNA) Mycoviruses

{kind=link}
| Families and Genera | Genome Size and Segmentation | Genome Organization | Exemplar Species |
|---|---|---|---|
| Family Amalgaviridae Mycoviral associated genera Zybavirus | Non-segmented genomes around 3.5 kb in size. | 5′ proximal ORF encodes for a CP, followed by ribosomal frameshift signal and a 3′ proximal ORF encoding for an RdRp protein. | Zygosaccharomyces bailii virus Z |
| Family Chrysoviridae Mycoviral associated genera Alphachrysovirus Betachrysovirus | Multisegmented genomes around 8.9 to 16.0 kb in size. Alphachrysovirus: 3–4 separately encapsidated segments. Betachrysovirus: 4–7 separately encapsidated genomes. | dsRNA 1: Encodes for an RdRp. dsRNA 2: Encodes for a CP. dsRNA 3: Encodes for phytoreo S7 domain in Alphachrysovirus, and an unknown protein in Betachrysovirus. dsRNA 4–7: Encodes for a hypothetical protein with unknown function. | Alphachrysovirus: Penicillium chrysogenum virus Betachrysovirus: Botryosphaeria dothidea chrysovirus 1 |
| Family Megabirnaviridae Mycoviral associated genera Megabirnavirus | Bisegmented genomes around 16 kb in size. | dsRNA 1: 5′ proximal ORF encodes for a CP, followed by a ribosomal frameshift signal and a 3′ proximal ORF encoding for an RdRp. dsRNA 2: Encodes for hypothetical protein with unknown function. | Rosellinia necatrix Megabirnavirus 1 |
| Family Partitiviridae Mycoviral associated genera Alphapartitivirus Betapartitivirus Gammapartivirus | Bisegmented genomes around 3 to 4.8 kb in size. | dsRNA 1: Encodes for an RdRp. dsRNA 2: Encodes for a CP. | Alphapartitivirus: Rosellinia necatrix partitivirus 2 Betapartitivirus: Ceratocystis resinifera virus 1 Gammapartivirus: Aspergillus ochraceous virus |
| Family Polymycoviridae Mycoviral associated genera Polymycovirus | Multisegmented genomes around 7.5 to 12.5 kb in size. | dsRNA 1: Encodes for an RdRp. dsRNA 2: Encodes for hypothetical protein with unknown function. dsRNA 3: Encodes for a Mtf. dsRNA 4: Encodes for a PASrp. dsRNA 5–8: Encodes for hypothetical proteins with unknown functions. | Aspergillus fumigatus tetramycovirus 1 |
| Family Polymycoviridae Mycoviral associated genera Polymycovirus | Multisegmented genomes around 7.5 to 12.5 kb in size. | dsRNA 1: Encodes for an RdRp. dsRNA 2: Encodes for hypothetical protein with unknown function. dsRNA 3: Encodes for a Mtf. dsRNA 4: Encodes for a PASrp. dsRNA 5–8: Encodes for hypothetical proteins with unknown functions. | Aspergillus fumigatus tetramycovirus 1 |
| Family Quadriviridae Mycoviral associated genera Quadrivirus | Quadripartite genomes around 3.5 to 5.0 kb in size. | dsRNA 1: Encodes for a hypothetical protein with unknown function. dsRNA 2 and 4: Encode for a CP. dsRNA 3: Encodes for an RdRp. | Rosellinia necatrix quadrivirus 1 |
| Family Spinareoviridae Mycoviral associated genera Mycoreovirus | Multisegmented genomes (11–12 segments) around 23 to 29 kb in size | Segments encodes for a single viral protein (VP1 to VP12) VP 1: Encodes for an RdRp VP 2: Encodes for CP VP 3 or VP10: Encodes for a capping enzyme, (guanylyltransferase) VP4 to VP12: Encodes for hypothetical proteins with unknown function | Mycoreovirus-1/Cp9B21 |
| Family Totiviridae Mycoviral associated genera Totivirus Victorivirus | Non-segmented genomes around 4.6–7.0 kb in size | 5′ proximal ORF encodes for CP and the 3′ proximal ORF encodes for an RdRp Totiviruses additionally encode for a ribosomal frameshift signal between the CP and RdRp Some totiviruses may additionally produce satellite dsRNA which encode for killer proteins | Totivirus: Saccharomyces cerevisiae virus L-A Victorivirus: Helminthosporium victoriae virus 190S |
| Family Unclassified Mycoviral associated genera Botybirnavirus | Bisegmented genomes around 5.7 to 6.3 kb in size | dsRNA 1: Encodes for an RdRp dsRNA 2: Encodes for a hypothetical protein with unknown function | Bipolaris maydis botybirnavirus 1 |
3.2.2. Single Stranded RNA (ssRNA) Mycoviruses
Positive (+) Sense
Negative (−) Sense
Reverse Transcribing (RT)
3.2.3. Single-Stranded DNA (ssDNA) Mycoviruses
4. Effect of Mycoviruses on Fungi
4.1. Host–Virus Arms Race
4.2. Hypervirulence, Hypovirulence and the Effect of Mycoviruses on the Pathogenesis of Fungi
4.3. The ‘Killer Phenotype’ in Yeasts
5. Mycoviruses as Biocontrol Agents
5.1. Limitations of Biocontrol: The Role of Mycovirus Transmission
5.1.1. Horizontal Transmission
5.1.2. Vertical Transmission
6. The Detection of Mycoviruses
6.1. In Vitro Based Detection
6.2. In Silico Based Detection
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ghabrial, S.A.; Suzuki, N. Viruses of Plant Pathogenic Fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.; Bonds, A.; Clemons, R.; Thapa, N.; Simmons, D.; Carter-House, D.; Ortanez, J.; Liu, P.; Miralles-Durán, A.; Desirò, A. Survey of early-diverging lineages of fungi reveals abundant and diverse mycoviruses. Mbio 2020, 11, e02027-20. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Yu, J.; Kim, K.-H. Five Questions about Mycoviruses. PLoS Pathog. 2015, 11, e1005172. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C. Changes to virus taxonomy and the Statutes ratified by the International Committee on Taxonomy of Viruses (2020). Arch. Virol. 2020, 165, 2737–2748. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479, 356–368. [Google Scholar] [CrossRef][Green Version]
- Kotta-Loizou, I. Mycoviruses and their role in fungal pathogenesis. Curr. Opin. Microbiol. 2021, 63, 10–18. [Google Scholar] [CrossRef]
- Sato, Y.; Turina, M.; Chiba, S.; Okada, R.; Bhatti, M.F.; Kotta-Loizou, I.; Coutts, R.H.; Kondo, H.; Sabanadzovic, S.; Suzuki, N. ICTV Virus Taxonomy Profile: Hadakaviridae 2023. J. Gen. Virol. 2023, 104, 001820. [Google Scholar] [CrossRef]
- Sato, Y.; Das, S.; Velasco, L.; Turina, M.; Osaki, H.; Kotta-Loizou, I.; Coutts, R.H.; Kondo, H.; Sabanadzovic, S.; Suzuki, N. ICTV Virus Taxonomy Profile: Yadokariviridae 2023. J. Gen. Virol. 2023, 104, 001826. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Fujita, M.; Chiba, S.; Hyodo, K.; Andika, I.B.; Suzuki, N.; Kondo, H. Two novel fungal negative-strand RNA viruses related to mymonaviruses and phenuiviruses in the shiitake mushroom (Lentinula edodes). Virology 2019, 533, 125–136. [Google Scholar] [CrossRef]
- Guo, M.; Shen, G.; Wang, J.; Liu, M.; Bian, Y.; Xu, Z. Mycoviral diversity and characteristics of a negative-stranded RNA virus LeNSRV1 in the edible mushroom Lentinula edodes. Virology 2021, 555, 89–101. [Google Scholar] [CrossRef]
- Wang, L.; He, H.; Wang, S.; Chen, X.; Qiu, D.; Kondo, H.; Guo, L. Evidence for a novel negative-stranded RNA mycovirus isolated from the plant pathogenic fungus Fusarium graminearum. Virology 2018, 518, 232–240. [Google Scholar] [CrossRef]
- Yu, X.; Li, B.; Fu, Y.; Jiang, D.; Ghabrial, S.A.; Li, G.; Peng, Y.; Xie, J.; Cheng, J.; Huang, J.; et al. A geminivirus-related DNA mycovirus that confers hypovirulence to a plant pathogenic fungus. Proc. Natl. Acad. Sci. USA 2010, 107, 8387–8392. [Google Scholar] [CrossRef][Green Version]
- Krupovic, M.; Ghabrial, S.A.; Jiang, D.; Varsani, A. Genomoviridae: A new family of widespread single-stranded DNA viruses. Arch. Virol. 2016, 161, 2633–2643. [Google Scholar] [CrossRef][Green Version]
- Varsani, A.; Krupovic, M. Family Genomoviridae: 2021 taxonomy update. Arch. Virol. 2021, 166, 2911–2926. [Google Scholar] [CrossRef]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef]
- Jiang, D.; Fu, Y.; Guoqing, L.; Ghabrial, S.A. Viruses of the plant pathogenic fungus Sclerotinia sclerotiorum. In Advances in Virus Research; Academic Press: New York, NY, USA, 2013; Volume 86, pp. 215–248. [Google Scholar]
- Hao, F.; Ding, T.; Wu, M.; Zhang, J.; Yang, L.; Chen, W.; Li, G. Two Novel Hypovirulence-Associated Mycoviruses in the Phytopathogenic Fungus Botrytis cinerea: Molecular Characterization and Suppression of Infection Cushion Formation. Viruses 2018, 10, 254. [Google Scholar] [CrossRef][Green Version]
- Wu, M.; Jin, F.; Zhang, J.; Yang, L.; Jiang, D.; Li, G. Characterization of a Novel Bipartite Double-Stranded RNA Mycovirus Conferring Hypovirulence in the Phytopathogenic Fungus Botrytis porri. Virol. J 2012, 86, 6605–6619. [Google Scholar] [CrossRef][Green Version]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal–plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef]
- Xie, J.; Jiang, D. New Insights into Mycoviruses and Exploration for the Biological Control of Crop Fungal Diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef][Green Version]
- Choi, G.H.; Dawe, A.L.; Churbanov, A.; Smith, M.L.; Milgroom, M.G.; Nuss, D.L. Molecular Characterization of Vegetative Incompatibility Genes That Restrict Hypovirus Transmission in the Chestnut Blight Fungus Cryphonectria parasitica. Genetics 2012, 190, 113–127. [Google Scholar] [CrossRef][Green Version]
- Lee, K.-M.; Yu, J.; Son, M.; Lee, Y.-W.; Kim, K.-H. Transmission of Fusarium boothii Mycovirus via Protoplast Fusion Causes Hypovirulence in Other Phytopathogenic Fungi. PLoS ONE 2011, 6, e21629. [Google Scholar] [CrossRef] [PubMed]
- Wagemans, J.; Holtappels, D.; Vainio, E.; Rabiey, M.; Marzachì, C.; Herrero, S.; Ravanbakhsh, M.; Tebbe, C.C.; Ogliastro, M.; Ayllón, M.A.; et al. Going Viral: Virus-Based Biological Control Agents for Plant Protection. Annu. Rev. Phytopathol. 2022, 60, 21–42. [Google Scholar] [CrossRef] [PubMed]
- Sinden, J.; Hauser, E. Report on two new mushroom diseases. Mushroom Sci. 1950, 1, 96–100. [Google Scholar]
- Hollings, M. Viruses Associated with A Die-Back Disease of Cultivated Mushroom. Nature 1962, 196, 962–965. [Google Scholar] [CrossRef]
- Ellis, L.; Kleinschmidt, W. Virus-like particles of a fraction of statolon, a mould product. Nature 1967, 215, 649–650. [Google Scholar] [CrossRef]
- Banks, G.; Buck, K.; Chain, E.; Himmelweit, F.; Marks, J.; Tyler, J.; Hollings, M.; Last, F.; Stone, O. Viruses in fungi and interferon stimulation. Nature 1968, 218, 542–545. [Google Scholar] [CrossRef]
- Van Alfen, N.; Jaynes, R.; Anagnostakis, S.; Day, P. Chestnut blight: Biological control by transmissible hypovirulence in Endothia parasitica. Science 1975, 189, 890–891. [Google Scholar] [CrossRef]
- Berry, E.; Bevan, E. A new species of double-stranded RNA from yeast. Nature 1972, 239, 279–280. [Google Scholar] [CrossRef]
- Maske, B.L.; Neto, D.P.D.C.; da Silva, G.B.; Lindner, J.D.D.; Soccol, C.R.; de Melo Pereira, G.V. Yeast viruses and their implications in fermented foods and beverages. Curr. Opin. Food Sci. 2022, 47, 100879. [Google Scholar] [CrossRef]
- Donaire, L.; Rozas, J.; Ayllón, M.A. Molecular characterization of Botrytis ourmia-like virus, a mycovirus close to the plant pathogenic genus Ourmiavirus. Virology 2016, 489, 158–164. [Google Scholar] [CrossRef][Green Version]
- Neupane, A.; Feng, C.; Feng, J.; Kafle, A.; Bücking, H.; Lee Marzano, S.-Y. Metatranscriptomic Analysis and In Silico Approach Identified Mycoviruses in the Arbuscular Mycorrhizal Fungus Rhizophagus spp. Viruses 2018, 10, 707. [Google Scholar] [CrossRef][Green Version]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef]
- Li, C.X.; Zhu, J.Z.; Gao, B.D.; Zhu, H.J.; Zhou, Q.; Zhong, J. Characterization of a Novel Ourmia-Like Mycovirus Infecting Magnaporthe oryzae and Implications for Viral Diversity and Evolution. Viruses 2019, 11, 223. [Google Scholar] [CrossRef][Green Version]
- Kwon, S.-J.; Lim, W.-S.; Park, S.-H.; Park, M.-R.; Kim, K.-H. Molecular characterization of a dsRNA mycovirus, Fusarium graminearum virus-DK21, which is phylogenetically related to hypoviruses but has a genome organization and gene expression strategy resembling those of plant potex-like viruses. Mol. Cell 2007, 23, 304. [Google Scholar] [CrossRef]
- Andika, I.B.; Wei, S.; Cao, C.; Salaipeth, L.; Kondo, H.; Sun, L. Phytopathogenic fungus hosts a plant virus: A naturally occurring cross-kingdom viral infection. Proc. Natl. Acad. Sci. USA 2017, 114, 12267–12272. [Google Scholar] [CrossRef][Green Version]
- Cao, X.; Liu, J.; Pang, J.; Kondo, H.; Chi, S.; Zhang, J.; Sun, L.; Andika, I.B. Common but Nonpersistent Acquisitions of Plant Viruses by Plant-Associated Fungi. Viruses 2022, 14, 2279. [Google Scholar] [CrossRef]
- Roossinck, M.J. Evolutionary and ecological links between plant and fungal viruses. New Phytol. 2019, 221, 86–92. [Google Scholar] [CrossRef][Green Version]
- Wang, Q.; Zou, Q.; Dai, Z.; Hong, N.; Wang, G.; Wang, L. Four Novel Mycoviruses from the Hypovirulent Botrytis cinerea SZ-2-3y Isolate from Paris polyphylla: Molecular Characterisation and Mitoviral Sequence Transboundary Entry into Plants. Viruses 2022, 14, 151. [Google Scholar] [CrossRef]
- Bian, R.; Andika, I.B.; Pang, T.; Lian, Z.; Wei, S.; Niu, E.; Wu, Y.; Kondo, H.; Liu, X.; Sun, L. Facilitative and synergistic interactions between fungal and plant viruses. Proc. Natl. Acad. Sci. USA 2020, 117, 3779–3788. [Google Scholar] [CrossRef]
- Kartali, T.; Zsindely, N.; Nyilasi, I.; Németh, O.; Sávai, G.N.; Kocsubé, S.; Lipinszki, Z.; Patai, R.; Spisák, K.; Nagy, G.; et al. Molecular Characterization of Novel Mycoviruses in Seven Umbelopsis Strains. Viruses 2022, 14, 2343. [Google Scholar] [CrossRef]
- Petrzik, K.; Koloniuk, I.; Sehadová, H.; Sarkisova, T. Chrysoviruses inhabited symbiotic fungi of lichens. Viruses 2019, 11, 1120. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wan, X.; Zhao, Y.; Zhang, Y.; Wei, C.; Du, H.; Zhang, H.; Chen, J.; Yang, L.; Zang, R.; Wen, C. Molecular characterization of a novel partitivirus isolated from the phytopathogenic fungus Aplosporella javeedii. Arch. Virol. 2021, 166, 1237–1240. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, M.; Zhu, H.; Zhong, J.; Liao, X.; Zhou, Q. Molecular characterization of a novel mitovirus from the plant-pathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2021, 166, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jiang, J.; Wang, Y.; Hong, N.; Zhang, F.; Xu, W.; Wang, G. Hypovirulence of the phytopathogenic fungus Botryosphaeria dothidea: Association with a coinfecting chrysovirus and a partitivirus. J. Virol. 2014, 88, 7517–7527. [Google Scholar] [CrossRef][Green Version]
- Zhai, L.; Xiang, J.; Zhang, M.; Fu, M.; Yang, Z.; Hong, N.; Wang, G. Characterization of a novel double-stranded RNA mycovirus conferring hypovirulence from the phytopathogenic fungus Botryosphaeria dothidea. Virology 2016, 493, 75–85. [Google Scholar] [CrossRef]
- Yang, M.; Wang, G.; Mu, W.; Guo, J.; Xi, J.; Hu, L.; Liang, K.; Xu, Q.; Hu, Z.; Song, J. A novel victorivirus isolated from the tobacco spot blight fungus Stagonosporopsis cucurbitacearum in China. Arch. Virol. 2022, 167, 2845–2850. [Google Scholar] [CrossRef]
- He, Y.; Zou, Q.; Li, S.; Zhu, H.; Hong, N.; Wang, G.; Wang, L. Molecular characterization of a new fusarivirus infecting Botryosphaeria dothidea, the causal agent of pear ring rot disease. Arch. Virol. 2022, 167, 1893–1897. [Google Scholar] [CrossRef]
- Khan, H.A.; Telengech, P.; Kondo, H.; Bhatti, M.F.; Suzuki, N. Mycovirus hunting revealed the presence of diverse viruses in a single isolate of the phytopathogenic fungus Diplodia seriata from Pakistan. Front. Cell. Infect. Microbiol. 2022, 12, 702. [Google Scholar] [CrossRef]
- De Wet, J.; Bihon, W.; Preisig, O.; Wingfield, B.D.; Wingfield, M.J. Characterization of a novel dsRNA element in the pine endophytic fungus Diplodia scrobiculata. Arch. Virol. 2011, 156, 1199–1208. [Google Scholar] [CrossRef][Green Version]
- Aday, A.; Lehtijarvi, A.; Doğmuş-Lehtijarvi, H. Frequency of virus in some Diplodia pinea and Gremmeniella abietina isolates originated from Turkey. J. Agric. Ext. Rural Dev. 2012, 4, 181–183. [Google Scholar] [CrossRef]
- Jo, Y.; Choi, H.; Chu, H.; Cho, W.K. Unveiling Mycoviromes Using Fungal Transcriptomes. Int. J. Mol. Sci. 2022, 23, 10926. [Google Scholar] [CrossRef]
- Hafez, E.E.; Aseel, D.G.; Mostafa, S. Two novel mycoviruses related to Geminivirus isolated from the soil-borne fungi Macrophomina phaseolina (Tassi) Goid. and Mucor racemosus Bull. Biotechnol. Biotechnol. Equip. 2013, 27, 4222–4226. [Google Scholar] [CrossRef][Green Version]
- Wang, J.; Xiao, Y.; Zhao, H.; Ni, Y.; Liu, X.; Zhao, X.; Wang, G.; Xiao, X.; Liu, H. A novel double-stranded RNA mycovirus that infects Macrophomina phaseolina. Arch. Virol. 2019, 164, 2411–2416. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ni, Y.; Liu, X.; Zhao, H.; Xiao, Y.; Xiao, X.; Li, S.; Liu, H. Divergent RNA viruses in Macrophomina phaseolina exhibit potential as virocontrol agents. Virus Evol. 2021, 7, veaa095. [Google Scholar] [CrossRef] [PubMed]
- Marzano, S.-Y.L.; Nelson, B.D.; Ajayi-Oyetunde, O.; Bradley, C.A.; Hughes, T.J.; Hartman, G.L.; Eastburn, D.M.; Domier, L.L. Identification of diverse mycoviruses through metatranscriptomics characterization of the viromes of five major fungal plant pathogens. J. Virol. 2016, 90, 6846–6863. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Marais, A.; Faure, C.; Comont, G.; Candresse, T.; Stempien, E.; Corio-Costet, M.-F. Characterization of the mycovirome of the phytopathogenic fungus, Neofusicoccum parvum. Viruses 2021, 13, 375. [Google Scholar] [CrossRef]
- Marais, A.; Nivault, A.; Faure, C.; Comont, G.; Theil, S.; Candresse, T.; Corio-Costet, M.-F. Molecular characterization of a novel fusarivirus infecting the plant-pathogenic fungus Neofusicoccum luteum. Arch. Virol. 2018, 163, 559–562. [Google Scholar] [CrossRef]
- Marais, A.; Nivault, A.; Faure, C.; Theil, S.; Comont, G.; Candresse, T.; Corio-Costet, M.-F. Determination of the complete genomic sequence of Neofusicoccum luteum mitovirus 1 (NLMV1), a novel mitovirus associated with a phytopathogenic Botryosphaeriaceae. Arch. Virol. 2017, 162, 2477–2480. [Google Scholar] [CrossRef][Green Version]
- Zhao, Y.; Du, H.; Liu, Y.; Zhong, R.; Guan, Z.; Wang, G.; Zhang, Y.; Wei, C.; Wang, M.; Wan, X. Molecular characterization of a novel victorivirus isolated from the phytopathogenic fungus Phaeobotryon rhois. Arch. Virol. 2023, 168, 15. [Google Scholar] [CrossRef]
- Gilbert, K.B.; Holcomb, E.E.; Allscheid, R.L.; Carrington, J.C. Hiding in plain sight: New virus genomes discovered via a systematic analysis of fungal public transcriptomes. PLoS ONE 2019, 14, e0219207. [Google Scholar] [CrossRef][Green Version]
- McHale, M.T.; Roberts, I.N.; Noble, S.M.; Beaumont, C.; Whitehead, M.P.; Seth, D.; Oliver, R.P. CfT-I: An LTR-retrotransposon in Cladosporium fulvum, a fungal pathogen of tomato. Mol. Gener. Genet. 1992, 233, 337–347. [Google Scholar] [CrossRef]
- Nerva, L.; Turina, M.; Zanzotto, A.; Gardiman, M.; Gaiotti, F.; Gambino, G.; Chitarra, W. Isolation, molecular characterization and virome analysis of culturable wood fungal endophytes in esca symptomatic and asymptomatic grapevine plants. Environ. Microbiol. 2019, 21, 2886–2904. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, M.; Yang, Y.; Liu, Q.; Zhang, Z.; Han, C.; Wang, Y. Characterization of the mycovirome from the plant-pathogenic fungus Cercospora beticola. Viruses 2021, 13, 1915. [Google Scholar] [CrossRef]
- Daudu, J.; Snowden, J.; Tubby, K.; Coutts, R.; Kotta-Loizou, I. Studying a mycovirus from Dothistroma septosporum, causative agent of pine needle blight. Access Microbiol 2019, 1, 783. [Google Scholar] [CrossRef]
- Lima, S.S.; Abadio, A.K.R.; Araujo, E.F.; Kitajima, E.W.; Sartorato, A.; Vieira de Queiroz, M. Mycovirus in Pseudocercospora griseola, the causal agent of angular leaf spot in common bean. Can. J. Microbiol. 2010, 56, 359–365. [Google Scholar] [CrossRef]
- Zelikovitch, N.; Eyal, Z.; Ben-Zvi, B.; Koltin, Y. Double-stranded RNA mycoviruses in Septoria tritici. Mycol. Res. 1990, 94, 590–594. [Google Scholar] [CrossRef]
- Cheng, J.; Jiang, D.; Fu, Y.; Li, G.; Peng, Y.; Ghabrial, S.A. Molecular characterization of a dsRNA totivirus infecting the sclerotial parasite Coniothyrium minitans. Virus Res. 2003, 93, 41–50. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Xue, C.; Xu, C.; Geng, Y.; Zang, R.; Guo, Y.; Wu, H.; Zhang, M. Complete genome sequence of a novel mycovirus isolated from the phytopathogenic fungus Corynespora cassiicola in China. Arch. Virol. 2020, 165, 2401–2404. [Google Scholar] [CrossRef]
- Petrzik, K.; Koloniuk, I.; Sarkisova, T.; Hrabáková, L. Detection and genome sequence of a new betapartitivirus associated with Cucurbitaria piceae Borthw. fungus causing bud blight of spruce in the Czech Republic. Arch. Virol. 2016, 161, 1405–1409. [Google Scholar] [CrossRef]
- Ye, L.; Shi, X.; He, Y.; Chen, J.; Xu, Q.; Shafik, K.; Xu, W. Characterization of a novel botybirnavirus with a unique dsRNA infecting Didymella theifolia from tea plants. bioRxiv 2022. [Google Scholar] [CrossRef]
- Zhou, J.; Hu, X.; Liang, X.; Wang, Y.; Xie, C.; Zheng, L. Complete genome sequence of a novel mycovirus from Phoma matteucciicola. Arch. Virol. 2021, 166, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Xu, G.; Zhou, J.; Xie, C.; Cui, H.; Miao, W.; Kang, Z.; Zheng, L. Complete genomic sequence and organization of a novel mycovirus from Phoma matteuccicola strain LG915. Arch. Virol. 2019, 164, 2209–2213. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, Y.; Liang, X.; Xie, C.; Liu, W.; Miao, W.; Kang, Z.; Zheng, L. Molecular characterization of a novel ourmia-like virus infecting Phoma matteucciicola. Viruses 2020, 12, 231. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhou, S.; Chen, D.; Fu, Y.; Zhou, J.; Yang, Y.; Xie, C.; Zheng, L. Characterization of a novel mycotombus-like virus from the plant pathogenic fungus Phoma matteucciicola. Arch. Virol. 2023, 168, 103. [Google Scholar] [CrossRef] [PubMed]
- Shah, U.A.; Kotta-Loizou, I.; Fitt, B.D.; Coutts, R.H. Identification, molecular characterization, and biology of a novel quadrivirus infecting the phytopathogenic fungus Leptosphaeria biglobosa. Viruses 2018, 11, 9. [Google Scholar] [CrossRef][Green Version]
- Shah, U.A.; Kotta-Loizou, I.; Fitt, B.D.; Coutts, R.H. Mycovirus-induced hypervirulence of Leptosphaeria biglobosa enhances systemic acquired resistance to Leptosphaeria maculans in Brassica napus. Mol. Plant Microbe Interact. 2020, 33, 98–107. [Google Scholar] [CrossRef][Green Version]
- Wang, Y.; Xu, Z.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. Characterization of a novel RNA virus from the phytopathogenic fungus Leptosphaeria biglobosa related to members of the genus Mitovirus. Arch. Virol. 2019, 164, 913–916. [Google Scholar] [CrossRef]
- Xie, J.; Havens, W.M.; Lin, Y.-H.; Suzuki, N.; Ghabrial, S.A. The victorivirus Helminthosporium victoriae virus 190S is the primary cause of disease/hypovirulence in its natural host and a heterologous host. Virus Res. 2016, 213, 238–245. [Google Scholar] [CrossRef]
- Dunkle, L.D. Double-stranded RNA mycovirus in Periconia circinata. Physiol. Plant Pathol. 1974, 4, 107–116. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, H.; Zhao, C.; Liu, S.; Guo, L. The complete genome sequence of a novel mycovirus from Alternaria longipes strain HN28. Arch. Virol. 2015, 160, 577–580. [Google Scholar] [CrossRef]
- Aoki, N.; Moriyama, H.; Kodama, M.; Arie, T.; Teraoka, T.; Fukuhara, T. A novel mycovirus associated with four double-stranded RNAs affects host fungal growth in Alternaria alternata. Virus Res. 2009, 140, 179–187. [Google Scholar] [CrossRef]
- Da Silva Xavier, A.; de Barros, A.P.O.; Godinho, M.T.; Zerbini, F.M.; de Oliveira Souza, F.; Bruckner, F.P.; Alfenas-Zerbini, P. A novel mycovirus associated to Alternaria alternata comprises a distinct lineage in Partitiviridae. Virus Res. 2018, 244, 21–26. [Google Scholar] [CrossRef]
- Okada, R.; Ichinose, S.; Takeshita, K.; Urayama, S.-i.; Fukuhara, T.; Komatsu, K.; Arie, T.; Ishihara, A.; Egusa, M.; Kodama, M. Molecular characterization of a novel mycovirus in Alternaria alternata manifesting two-sided effects: Down-regulation of host growth and up-regulation of host plant pathogenicity. Virology 2018, 519, 23–32. [Google Scholar] [CrossRef]
- Komatsu, K.; Katayama, Y.; Omatsu, T.; Mizutani, T.; Fukuhara, T.; Kodama, M.; Arie, T.; Teraoka, T.; Moriyama, H. Genome sequence of a novel victorivirus identified in the phytopathogenic fungus Alternaria arborescens. Arch. Virol. 2016, 161, 1701–1704. [Google Scholar] [CrossRef]
- Wang, W.; Wang, X.; Tu, C.; Yang, M.; Xiang, J.; Wang, L.; Hong, N.; Zhai, L.; Wang, G. Novel Mycoviruses Discovered from a Metatranscriptomics Survey of the Phytopathogenic Alternaria Fungus. Viruses 2022, 14, 2552. [Google Scholar] [CrossRef]
- Chen, Y.; Shang, H.H.; Yang, H.Q.; Da Gao, B.; Zhong, J. A mitovirus isolated from the phytopathogenic fungus Alternaria brassicicola. Arch. Virol. 2017, 162, 2869–2874. [Google Scholar] [CrossRef]
- Li, H.; Bian, R.; Liu, Q.; Yang, L.; Pang, T.; Salaipeth, L.; Andika, I.B.; Kondo, H.; Sun, L. Identification of a novel hypovirulence-inducing hypovirus from Alternaria alternata. Front. Microbiol. 2019, 10, 1076. [Google Scholar] [CrossRef][Green Version]
- Wang, H.; Li, C.; Cai, L.; Fang, S.; Zheng, L.; Yan, F.; Zhang, S.; Liu, Y. The complete genomic sequence of a novel botybirnavirus isolated from a phytopathogenic Bipolaris maydis. Virus Genes 2018, 54, 733–736. [Google Scholar] [CrossRef]
- Deng, Q.; Wang, H.; Li, C.; Li, P.; Fang, S.; Yang, S.; Yan, F.; Zhang, S.; Chen, Z. The complete genomic sequence of a novel alphapartitivirus from Bipolaris maydis, the causal agent of corn southern leaf blight. Arch. Virol. 2017, 162, 2433–2436. [Google Scholar] [CrossRef]
- Suikphopetch, P.; Suwanmanee, S.; Pumeesat, P.; Ampawong, S. In vitro characterization of Chrysovirus-1-induced hypovirulence of Bipolaris maydis. Walailak. J. Sci. Technol. 2021, 18, 6564. [Google Scholar] [CrossRef]
- Li, Q.; Huang, W.; Hai, D.; Wang, Y.; Xie, J.; Wang, M. The complete genome sequence of a novel hypovirus infecting Bipolaris oryzae. Arch. Virol. 2020, 165, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Soldevila, A.I.; Havens, W.M. Molecular genetics of the viruses infecting the plant pathogenic fungus Helminthosporium victoriae. In dsRNA Genetic Elements: Concepts and Applications in Agriculture, Forestry, and Medicine; Tavantzis, S.M., Ed.; CRC Press: Boca Raton, FL, USA, 2002; pp. 213–236. [Google Scholar]
- Márquez, L.M.; Redman, R.S.; Rodriguez, R.J.; Roossinck, M.J. A virus in a fungus in a plant: Three-way symbiosis required for thermal tolerance. Science 2007, 315, 513–515. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shepherd, H.S. Characterization and localization of a virus-like particle in a Drechslera species. Exp. Mycol. 1990, 14, 294–298. [Google Scholar] [CrossRef]
- Livore, A.B.; Grubb, P.; Magill, C.W.; Magill, J.M. A Sephadex column procedure for DNA isolation is also useful for detecting dsRNA. Nucleic Acids Res. 1988, 16, 776. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zheng, Y.; Yin, S.; Zhao, Y.; Li, S.; Lu, Z.; Li, Z.; Deng, Q.; Li, Z.; Zhang, S.; Fang, S. Molecular and biological characteristics of a novel chrysovirus infecting the fungus phytopathogenic Setosphaeria turcica f.sp. sorghi. Virus Res. 2023, 325, 199037. [Google Scholar] [CrossRef]
- Nerva, L.; Ciuffo, M.; Vallino, M.; Margaria, P.; Varese, G.; Gnavi, G.; Turina, M. Multiple approaches for the detection and characterization of viral and plasmid symbionts from a collection of marine fungi. Virus Res. 2016, 219, 22–38. [Google Scholar] [CrossRef]
- Gao, Z.; Cai, L.; Liu, M.; Wang, X.; Yang, J.; An, H.; Deng, Q.; Zhang, S.; Fang, S. A novel previously undescribed fusarivirus from the phytopathogenic fungus Setosphaeria turcica. Arch. Virol. 2021, 166, 665–669. [Google Scholar] [CrossRef]
- Gao, Z.; Zhang, M.; Yu, T.; Wang, X.; Wang, X.; An, H.; Zhang, S.; Liu, M.; Fang, S. Molecular characterization of a novel polymycovirus from the phytopathogenic fungus Setosphaeria turcica. Arch. Virol. 2021, 166, 2315–2319. [Google Scholar] [CrossRef]
- Liu, H.; Wang, H.; Liao, X.L.; Gao, B.; Lu, X.; Sun, D.; Gong, W.; Zhong, J.; Zhu, H.; Pan, X. Mycoviral gene integration converts a plant pathogenic fungus into a biocontrol agent. Proc. Natl. Acad. Sci. USA 2022, 119, e2214096119. [Google Scholar] [CrossRef]
- McGinty, R.; Buck, K.; Rawlinson, C. Transcriptase activity assoclated with a type 2 double-stranded RNA mycovirus. Biochem. Biophys. Res. Commun. 1981, 98, 501–506. [Google Scholar] [CrossRef]
- Buck, K.; McGinty, R.; Rawlinson, C. Two Serologically Unrelated Viruses Isolated from a Phialophora sp. J. Gen. Virol. 1981, 55, 235–239. [Google Scholar] [CrossRef]
- Kanhayuwa, L.; Kotta-Loizou, I.; Özkan, S.; Gunning, A.P.; Coutts, R.H.A. A novel mycovirus from Aspergillus fumigatus contains four unique dsRNAs as its genome and is infectious as dsRNA. Proc. Natl. Acad. Sci. USA 2015, 112, 9100–9105. [Google Scholar] [CrossRef][Green Version]
- Kotta-Loizou, I.; Coutts, R.H. Mycoviruses in Aspergilli: A comprehensive review. Front. Microbiol. 2017, 8, 1699. [Google Scholar] [CrossRef][Green Version]
- Zoll, J.; Verweij, P.E.; Melchers, W.J. Discovery and characterization of novel Aspergillus fumigatus mycoviruses. PLoS ONE 2018, 13, e0200511. [Google Scholar] [CrossRef]
- Kinsella, C.M.; Deijs, M.; Gittelbauer, H.; van der Hoek, L.; van Dijk, K. Human clinical isolates of pathogenic fungi are host to diverse mycoviruses. Microbiol. Spectr. 2022, 10, e01610-22. [Google Scholar] [CrossRef]
- Niu, Y.; Yuan, Y.; Mao, J.; Yang, Z.; Cao, Q.; Zhang, T.; Wang, S.; Liu, D. Characterization of two novel mycoviruses from Penicillium digitatum and the related fungicide resistance analysis. Sci. Rep. 2018, 8, 5513. [Google Scholar] [CrossRef][Green Version]
- DeMarini, D.; Kurtzman, C.; Fennell, D.; Worden, K.; Detroy, R. Transmission of PsV-F and PsV-S mycoviruses during conidiogenesis of Penicillium stoloniferum. Microbiology 1977, 100, 59–64. [Google Scholar] [CrossRef][Green Version]
- Lau, S.K.; Lo, G.C.; Chow, F.W.; Fan, R.Y.; Cai, J.J.; Yuen, K.-Y.; Woo, P.C. Novel partitivirus enhances virulence of and causes aberrant gene expression in Talaromyces marneffei. MBio 2018, 9, e00947-18. [Google Scholar] [CrossRef][Green Version]
- Jiang, Y.; Tian, X.; Liu, X.; Yang, B.; Wang, N.; Wang, Q.; Yu, W.; Qi, X.; Peng, J.; Hsiang, T. Complete genome sequence of a novel chrysovirus infecting Talaromyces neofusisporus. Arch. Virol. 2022, 167, 2789–2793. [Google Scholar] [CrossRef]
- Teng, L.; Chen, S.; Hu, Z.; Chen, J.; Liu, H.; Zhang, T. Molecular characterization and transcriptomic analysis of a novel polymycovirus in the fungus Talaromyces amestolkiae. Front. Microbiol. 2022, 13, 4249. [Google Scholar] [CrossRef]
- Kohno, S.; Fujimura, T.; Rulong, S.; Kwon-Chung, K. Double-stranded RNA virus in the human pathogenic fungus Blastomyces dermatitidis. J. Virol. 1994, 68, 7554–7558. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, Z.; Su, S.; Hamilton, M.; Yan, L.; Chen, Y. The ability to cause infection in a pathogenic fungus uncovers a new biological feature of honey bee viruses. J. Invertebr. Pathol. 2014, 120, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Zhang, L.; Luo, J.; Yang, S.; Deng, Y.; Li, J.; Hou, C. Two pathogenic fungi isolated from chalkbrood samples and honey bee viruses they carried. Front. Microbiol. 2022, 13, 806. [Google Scholar] [CrossRef] [PubMed]
- Nerva, L.; Forgia, M.; Ciuffo, M.; Chitarra, W.; Chiapello, M.; Vallino, M.; Varese, G.; Turina, M. The mycovirome of a fungal collection from the sea cucumber Holothuria polii. Virus Res. 2019, 273, 197737. [Google Scholar] [CrossRef]
- Kim, J.-M.; Jung, J.-E.; Park, J.-A.; Park, S.-M.; Cha, B.-J.; Kim, D.-H. Biological function of a novel chrysovirus, CnV1-BS122, in the Korean Cryphonectria nitschkei BS122 strain. J. Biosci. Bioeng. 2013, 115, 1–3. [Google Scholar] [CrossRef]
- Deng, F.; Allen, T.D.; Hillman, B.I.; Nuss, D.L. Comparative Analysis of Alterations in Host Phenotype and Transcript Accumulation following Hypovirus and Mycoreovirus Infections of the Chestnut Blight Fungus Cryphonectria parasitica. Eukaryot. Cell. 2007, 6, 1286–1298. [Google Scholar] [CrossRef][Green Version]
- Cornejo, C.; Hisano, S.; Bragança, H.; Suzuki, N.; Rigling, D. A new double-stranded RNA mycovirus in Cryphonectria naterciae is able to cross the species barrier and is deleterious to a new host. J. Fungi 2021, 7, 861. [Google Scholar] [CrossRef]
- Nuskern, L.; Stojanović, M.; Milanović-Litre, M.; Šibenik, T.; Ježić, M.; Poljak, I.; Ćurković-Perica, M. Filling the Gap in Southern Europe—Diversity of Cryphonectria parasitica and Associated Mycovirus (Cryphonectria hypovirus 1) in Montenegro. J. Fungi 2022, 8, 552. [Google Scholar] [CrossRef]
- Rumbou, A.; Vainio, E.J.; Büttner, C. Towards the forest virome: High-throughput sequencing drastically expands our understanding on virosphere in temperate forest ecosystems. Microorganisms 2021, 9, 1730. [Google Scholar] [CrossRef]
- Hansen, D.R.; Van Alfen, N.K.; Gillies, K.; Powell, W.A. Naked dsRNA associated with hypovirulence of Endothia parasitica is packaged in fungal vesicles. J. Gen. Virol. 1985, 66, 2605–2614. [Google Scholar] [CrossRef]
- Preisig, O.; Moleleki, N.; Smit, W.; Wingfield, B.; Wingfield, M. A novel RNA mycovirus in a hypovirulent isolate of the plant pathogen Diaporthe ambigua. J. Gen. Virol. 2000, 81, 3107–3114. [Google Scholar] [CrossRef]
- Xu, G.; Zhang, X.; Liang, X.; Chen, D.; Xie, C.; Kang, Z.; Zheng, L. A novel hexa-segmented dsRNA mycovirus confers hypovirulence in the phytopathogenic fungus Diaporthe pseudophoenicicola. Environ. Microbiol. 2022, 24, 4274–4284. [Google Scholar] [CrossRef]
- Shafik, K.; Umer, M.; You, H.; Aboushedida, H.; Wang, Z.; Ni, D.; Xu, W. Characterization of a novel mitovirus infecting Melanconiella theae isolated from tea plants. Front. Microbiol. 2021, 12, 757556. [Google Scholar] [CrossRef]
- Peyambari, M.; Habibi, M.K.; Fotouhifar, K.-B.; Dizadji, A.; Roossinck, M.J. Molecular characterization of a novel putative partitivirus infecting Cytospora sacchari, a plant pathogenic fungus. Plant Pathol. J. 2014, 30, 151. [Google Scholar] [CrossRef][Green Version]
- Hrabáková, L.; Koloniuk, I.; Petrzik, K. Phomopsis longicolla RNA virus 1–Novel virus at the edge of myco-and plant viruses. Virology 2017, 506, 14–18. [Google Scholar] [CrossRef]
- Zhang, C.J.; Zhou, X.Y.; Zhong, J.; Guo, J.; Yang, X.P.; Zhu, H.J. Complete nucleotide sequence of a novel partitivirus infecting the plant-pathogenic fungus Phomopsis vexans. Arch. Virol. 2021, 166, 291–294. [Google Scholar] [CrossRef]
- Zhang, R.J.; Zhong, J.; Shang, H.H.; Pan, X.T.; Zhu, H.J.; Da Gao, B. The complete nucleotide sequence and genomic organization of a novel victorivirus with two non-overlapping ORFs, identified in the plant-pathogenic fungus Phomopsis vexans. Arch. Virol. 2015, 160, 1805–1809. [Google Scholar] [CrossRef]
- Xie, F.L.; Zhou, X.Y.; Xiao, R.; Zhang, C.J.; Zhong, J.; Zhou, Q.; Liu, F.; Zhu, H.J. Discovery and exploration of widespread infection of mycoviruses in Phomopsis vexans, the causal agent of phomopsis blight of eggplant in China. Front. Plant Sci. 2022, 13, 996862. [Google Scholar] [CrossRef]
- Yaegashi, H.; Kanematsu, S.; Ito, T. Molecular characterization of a new hypovirus infecting a phytopathogenic fungus, Valsa ceratosperma. Virus Res. 2012, 165, 143–150. [Google Scholar] [CrossRef]
- Xu, X.; Hai, D.; Li, J.; Huang, F.; Wang, Y. Molecular characterization of a novel penoulivirus from the phytopathogenic fungus Colletotrichum camelliae. Arch. Virol. 2022, 167, 641–644. [Google Scholar] [CrossRef]
- Guo, J.; Zhu, J.Z.; Zhou, X.Y.; Zhong, J.; Li, C.H.; Zhang, Z.G.; Zhu, H.J. A novel ourmia-like mycovirus isolated from the plant pathogenic fungus Colletotrichum gloeosporioides. Arch. Virol. 2019, 164, 2631–2635. [Google Scholar] [CrossRef] [PubMed]
- Suharto, A.R.; Jirakkakul, J.; Eusebio-Cope, A.; Salaipeth, L. Hypovirulence of Colletotrichum gloesporioides Associated with dsRNA Mycovirus Isolated from a Mango Orchard in Thailand. Viruses 2022, 14, 1921. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Zhang, H.; Yin, M.; Han, Z.; Bai, Q.; Peng, Y.; Shafik, K.; Zhai, L.; Hong, N.; Xu, W. A novel heptasegmented positive-sense single-stranded RNA virus from the phytopathogenic fungus Colletotrichum fructicola. J. Virol. 2022, 96, e00318-22. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.Z.; Guo, J.; Hu, Z.; Zhang, X.T.; Li, X.G.; Zhong, J. A novel partitivirus that confer hypovirulence to the plant pathogenic fungus Colletotrichum liriopes. Front. Microbiol. 2021, 12, 653809. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Chen, D.; Lei, X.H.; Zhu, H.J.; Zhu, J.Z.; Da Gao, B. Detection and characterization of a novel Gammapartitivirus in the phytopathogenic fungus Colletotrichum acutatum strain HNZJ001. Virus Res. 2014, 190, 104–109. [Google Scholar] [CrossRef]
- Li, C.; Li, M.; Gao, Z.; Gong, D.; Hong, X.; Jiang, Y.; Chang, S.; Hu, M. Mycoviruses of Colletotrichum spp.: A review. South. J. Agric. 2020, 51, 123–132. [Google Scholar]
- Hrabáková, L.; Grum-Grzhimaylo, A.A.; Koloniuk, I.; Debets, A.J.; Sarkisova, T.; Petrzik, K. The alkalophilic fungus Sodiomyces alkalinus hosts beta-and gammapartitiviruses together with a new fusarivirus. PLoS ONE 2017, 12, e0187799. [Google Scholar] [CrossRef][Green Version]
- Cañizares, M.C.; López-Escudero, F.J.; Pérez-Artés, E.; García-Pedrajas, M.D. Characterization of a novel single-stranded RNA mycovirus related to invertebrate viruses from the plant pathogen Verticillium dahliae. Arch. Virol. 2018, 163, 771–776. [Google Scholar] [CrossRef]
- Feng, Z.; Zhu, H.; Li, Z.; Shi, Y.; Zhao, L.; Liu, L.; Jiang, D. Complete genome sequence of a novel dsRNA mycovirus isolated from the phytopathogenic fungus Verticillium dahliae Kleb. Arch. Virol. 2013, 158, 2621–2623. [Google Scholar] [CrossRef]
- Cañizares, M.C.; Pérez-Artés, E.; García-Pedrajas, M.D. The complete nucleotide sequence of a novel partitivirus isolated from the plant pathogenic fungus Verticillium albo-atrum. Arch. Virol. 2014, 159, 3141–3144. [Google Scholar] [CrossRef][Green Version]
- Gao, J.; Wang, N.; Jia, J.; Xie, J.; Jiang, D.; Zhang, L. Characterization of a novel magoulivirus isolated from the phytopathogenic fungus Verticillium dahlia. Arch. Virol. 2022, 167, 2387–2390. [Google Scholar] [CrossRef]
- Khalifa, M.E.; MacDiarmid, R.M. A novel totivirus naturally occurring in two different fungal genera. Front. Microbiol. 2019, 10, 2318. [Google Scholar] [CrossRef]
- Oh, C.-S.; Hillman, B.I. Genome organization of a partitivirus from the filamentous ascomycete Atkinsonella hypoxylon. J. Gen. Virol. 1995, 76, 1461–1470. [Google Scholar] [CrossRef]
- Romo, M.; Leuchtmann, A.; García, B.; Zabalgogeazcoa, I. Double-stranded RNA viruses infecting Epichloe festucae. N. Z. J. Agric. Res. 2006, 13, 207–209. [Google Scholar] [CrossRef]
- Romo, M.; Leuchtmann, A.; García, B.; Zabalgogeazcoa, I. A totivirus infecting the mutualistic fungal endophyte Epichloë festucae. Virus Res. 2007, 124, 38–43. [Google Scholar] [CrossRef]
- De la Paz Giménez-Pecci, M.; Bogo, M.R.; Santi, L.; Moraes, C.K.D.; Corrêa, C.T.; Henning Vainstein, M.; Schrank, A. Characterization of mycoviruses and analyses of chitinase secretion in the biocontrol fungus Metarhizium anisopliae. Curr. Microbiol. 2002, 45, 334–339. [Google Scholar] [CrossRef]
- Wang, P.; Yang, G.; Shi, N.; Huang, B. Molecular characterization of a novel double-stranded RNA virus infecting the entomopathogenic fungus Metarhizium brunneum. Arch. Microbiol. 2022, 204, 606. [Google Scholar] [CrossRef]
- Wang, P.; Yang, G.; Shi, N.; Huang, B. Molecular characterization of a new partitivirus, MbPV1, isolated from the entomopathogenic fungus Metarhizium brunneum in China. Arch. Virol. 2020, 165, 765–769. [Google Scholar] [CrossRef]
- He, L.; Wang, P.; Yang, G.; Chen, X.; Huang, B. A novel polymycovirus infecting the entomopathogenic fungus Metarhizium brunneum. Arch. Virol. 2023, 168, 6. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, T.; Luo, C.; Jiang, D.; Li, G.; Li, Q.; Hsiang, T.; Huang, J. Prevalence and diversity of mycoviruses infecting the plant pathogen Ustilaginoidea virens. Virus Res. 2015, 195, 47–56. [Google Scholar] [CrossRef]
- Zhong, J.; Lei, X.H.; Zhu, J.Z.; Song, G.; Zhang, Y.D.; Chen, Y.; Gao, B.D. Detection and sequence analysis of two novel co-infecting double-strand RNA mycoviruses in Ustilaginoidea virens. Arch. Virol. 2014, 159, 3063–3070. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.J.; Chen, D.; Zhong, J.; Zhang, S.Y.; Gao, B.D. A novel mycovirus identified from the rice false smut fungus Ustilaginoidea virens. Virus Genes 2015, 51, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Jiang, Y.; Dong, W. A novel monopartite dsRNA virus isolated from the phytopathogenic fungus Ustilaginoidea virens and ancestrally related to a mitochondria-associated dsRNA in the green alga Bryopsis. Virology 2014, 462, 227–235. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhong, J.; Zhou, Q.; Lei, X.H.; Chen, D.; Shang, H.H.; Zhu, H.J. The nucleotide sequence and genome organization of two victoriviruses from the rice false smut fungus Ustilaginoidea virens. Virus Genes 2014, 48, 570–573. [Google Scholar] [CrossRef] [PubMed]
- Shi, N.; Hu, F.; Wang, P.; Zhang, Y.; Zhu, Q.; Yang, G.; Huang, B. Molecular characterization of two dsRNAs that could correspond to the genome of a new mycovirus that infects the entomopathogenic fungus Beauveria bassiana. Arch. Virol. 2021, 166, 3233–3237. [Google Scholar] [CrossRef]
- Filippou, C.; Diss, R.M.; Daudu, J.O.; Coutts, R.H.; Kotta-Loizou, I. The polymycovirus-mediated growth enhancement of the entomopathogenic fungus Beauveria bassiana is dependent on carbon and nitrogen metabolism. Front. Microbiol. 2021, 12, 606366. [Google Scholar] [CrossRef]
- Koloniuk, I.; Hrabáková, L.; Petrzik, K. Molecular characterization of a novel amalgavirus from the entomopathogenic fungus Beauveria bassiana. Arch. Virol. 2015, 160, 1585–1588. [Google Scholar] [CrossRef]
- Kotta-Loizou, I.; Coutts, R.H.A. Studies on the Virome of the Entomopathogenic Fungus Beauveria bassiana Reveal Novel dsRNA Elements and Mild Hypervirulence. PLoS Pathog. 2017, 13, e1006183. [Google Scholar] [CrossRef][Green Version]
- Zhang, Y.; Shi, N.; Wang, P.; Zhu, Q.; Yang, G.; Huang, B. Molecular characterization of a novel alternavirus infecting the entomopathogenic fungus Cordyceps chanhua. Arch. Virol. 2022, 167, 1467–1470. [Google Scholar] [CrossRef]
- Zhu, Q.; Shi, N.; Wang, P.; Zhang, Y.; Peng, F.; Yang, G.; Huang, B. A Novel Gammapartitivirus That Causes Changes in Fungal Development and Multi-Stress Tolerance to Important Medicinal Fungus Cordyceps chanhua. J. Fungi 2022, 8, 1309. [Google Scholar] [CrossRef]
- Herrero, N. Identification and sequence determination of a new chrysovirus infecting the entomopathogenic fungus Isaria javanica. Arch. Virol. 2017, 162, 1113–1117. [Google Scholar] [CrossRef]
- Petrzik, K.; Siddique, A.B. A mycoparasitic and opportunistic fungus is inhabited by a mycovirus. Arch. Virol. 2019, 164, 2545–2549. [Google Scholar] [CrossRef]
- Liu, C.; Li, M.; Redda, E.T.; Mei, J.; Zhang, J.; Wu, B.; Jiang, X. A novel double-stranded RNA mycovirus isolated from Trichoderma harzianum. Virol. J. 2019, 16, 113. [Google Scholar] [CrossRef]
- Chun, J.; Yang, H.-E.; Kim, D.-H. Identification and Molecular Characterization of a Novel Partitivirus from Trichoderma atroviride NFCF394. Viruses 2018, 10, 578. [Google Scholar] [CrossRef][Green Version]
- Lee, S.H.; Yun, S.-H.; Chun, J.; Kim, D.-H. Characterization of a novel dsRNA mycovirus of Trichoderma atroviride NFCF028. Arch. Virol. 2017, 162, 1073–1077. [Google Scholar] [CrossRef]
- Yun, S.-H.; Lee, S.H.; So, K.-K.; Kim, J.-M.; Kim, D.-H. Incidence of diverse dsRNA mycoviruses in Trichoderma spp. causing green mold disease of shiitake Lentinula edodes. FEMS Microbiol. Lett. 2016, 363, fnw220. [Google Scholar] [CrossRef][Green Version]
- Li, P.; Bhattacharjee, P.; Wang, S.; Zhang, L.; Ahmed, I.; Guo, L. Mycoviruses in Fusarium species: An update. Front. Cell. Infect. Microbiol. 2019, 9, 257. [Google Scholar] [CrossRef][Green Version]
- Zhao, Y.; Zhang, Y.; Wan, X.; She, Y.; Li, M.; Xi, H.; Xie, J.; Wen, C. A novel ourmia-like mycovirus confers hypovirulence-associated traits on Fusarium oxysporum. Front. Microbiol. 2020, 11, 569869. [Google Scholar] [CrossRef]
- Wang, J.; Li, C.; Song, P.; Qiu, R.; Song, R.; Li, X.; Ni, Y.; Zhao, H.; Liu, H.; Li, S. Molecular and Biological Characterization of the First Mymonavirus Identified in Fusarium oxysporum. Front. Microbiol. 2022, 13, 870204. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, B.; Qi, K.; Zhang, Y.; Ma, L.; Jiang, H.; Qin, S.; Qi, J. Characterization of a novel mitovirus isolated from the phytopathogenic fungus Fusarium pseudograminearum. Arch. Virol. 2022, 167, 1369–1373. [Google Scholar] [CrossRef]
- Jacquat, A.G.; Theumer, M.G.; Cañizares, M.C.; Debat, H.J.; Iglesias, J.; García Pedrajas, M.D.; Dambolena, J.S. A survey of mycoviral infection in Fusarium spp. isolated from maize and sorghum in argentina identifies the first mycovirus from Fusarium verticillioides. Viruses 2020, 12, 1161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, X.; Bhattacharjee, P.; Shi, Y.; Guo, L.; Wang, S. Molecular characterization of a novel strain of Fusarium graminearum virus 1 infecting Fusarium graminearum. Viruses 2020, 12, 357. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, P.; Wang, S.; Zhang, L.; Qiu, D.; Zhou, X.; Guo, L. A tripartite ssDNA mycovirus from a plant pathogenic fungus is infectious as cloned DNA and purified virions. Sci. Adv. 2020, 6, eaay9634. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lutz, T.; Langer, G.; Heinze, C. Complete genome sequence of a novel alternavirus infecting the fungus Ilyonectria crassa. Arch. Virol. 2023, 168, 34. [Google Scholar] [CrossRef]
- Pielhop, T.P.; Popp, C.; Knierim, D.; Margaria, P.; Maiß, E. First report of a chrysovirus infecting a member of the fungal genus Ilyonectria. Arch. Virol. 2022, 167, 2411–2415. [Google Scholar] [CrossRef]
- Ahn, I.-P.; Lee, Y.-H. A viral double-stranded RNA up regulates the fungal virulence of Nectria radicicola. Mol. Plant Microbe Interact. 2001, 14, 496–507. [Google Scholar] [CrossRef][Green Version]
- Pielhop, T.P.; Popp, C.; Knierim, D.; Margaria, P.; Mai, E. Three new mycoviruses identified in the apple replant disease (ARD)-associated fungus Rugonectria rugulosa. Virus Genes 2022, 58, 423–435. [Google Scholar] [CrossRef]
- Lutz, T.; Langer, G.; Heinze, C. Complete genome sequence of a new quadrivirus infecting a member of the genus Thelonectria. Arch. Virol. 2022, 167, 691–694. [Google Scholar] [CrossRef]
- Herrero, N.; Zabalgogeazcoa, I. Mycoviruses infecting the endophytic and entomopathogenic fungus Tolypocladium cylindrosporum. Virus Res. 2011, 160, 409–413. [Google Scholar] [CrossRef][Green Version]
- Herrero, N.; Sánchez Márquez, S.; Zabalgogeazcoa, I. Mycoviruses are common among different species of endophytic fungi of grasses. Arch. Virol. 2009, 154, 327–330. [Google Scholar] [CrossRef]
- Chen, H.; Jin, L.; Jiang, X.; Yu, Z.; Duns, G.J.; Shao, R.; Xu, W.; Chen, J. A novel mitovirus from Buergenerula spartinae infecting the invasive species Spartina alterniflora. Virol. Sin. 2014, 29, 257–260. [Google Scholar] [CrossRef]
- Jamil, N.; Buck, K.; Carlile, M. Sequence relationships between virus double-stranded RNA from isolates of Gaeumannomyces graminis in different vegetative compatibility groups. J. Gen. Virol. 1984, 65, 1741–1747. [Google Scholar] [CrossRef]
- Chun, S.J.; Lee, Y.-H. Inheritance of dsRNAs in the rice blast fungus, Magnaporthe grisea. FEMS Microbiol. Lett. 1997, 148, 159–162. [Google Scholar] [CrossRef]
- Aihara, M.; Urayama, S.-I.; Le, M.T.; Katoh, Y.; Higashiura, T.; Fukuhara, T.; Arie, T.; Teraoka, T.; Komatsu, K.; Moriyama, H. Infection by Magnaporthe oryzae chrysovirus 1 strain A triggers reduced virulence and pathogenic race conversion of its host fungus, Magnaporthe oryzae. J. Gen. Plant Pathol. 2018, 84, 92–103. [Google Scholar] [CrossRef]
- Lin, Y.; Zhou, J.; Zhou, X.; Shuai, S.; Zhou, R.; An, H.; Fang, S.; Zhang, S.; Deng, Q. A novel narnavirus from the plant-pathogenic fungus Magnaporthe oryzae. Arch. Virol. 2020, 165, 1235–1240. [Google Scholar] [CrossRef]
- Urayama, S.; Kato, S.; Suzuki, Y.; Aoki, N.; Le, M.T.; Arie, T.; Teraoka, T.; Fukuhara, T.; Moriyama, H. Mycoviruses related to chrysovirus affect vegetative growth in the rice blast fungus Magnaporthe oryzae. J. Gen. Virol. 2010, 91, 3085–3094. [Google Scholar] [CrossRef]
- Park, Y.; Chen, X.; Punja, Z.K. Molecular and biological characterization of a mitovirus in Chalara elegans (Thielaviopsis basicola). Phytopathology 2006, 96, 468–479. [Google Scholar] [CrossRef][Green Version]
- Park, Y.; James, D.; Punja, Z.K. Co-infection by two distinct totivirus-like double-stranded RNA elements in Chalara elegans (Thielaviopsis basicola). Virus Res. 2005, 109, 71–85. [Google Scholar] [CrossRef]
- Espinal, R.B.A.; de Santana, S.F.; Santos, V.C.; Lizardo, G.N.R.; Silva, R.J.S.; Corrêa, R.X.; Loguercio, L.L.; Neto, A.G.; Pirovani, C.P.; Fonseca, P.L.C. Uncovering a Complex Virome Associated with the Cacao Pathogens Ceratocystis cacaofunesta and Ceratocystis fimbriata. Pathogens 2023, 12, 287. [Google Scholar] [CrossRef]
- Deng, F.; Boland, G.J. Natural occurrence of a partitivirus in the sapstaining fungus Ceratocystis resinifera. Can. J. Plant Pathol. 2007, 29, 182–189. [Google Scholar] [CrossRef]
- Ohkita, S.; Lee, Y.; Nguyen, Q.; Ikeda, K.; Suzuki, N.; Nakayashiki, H. Three ourmia-like viruses and their associated RNAs in Pyricularia oryzae. Virology 2019, 534, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Hunst, P.L.; Latterell, F.M.; Rossi, A.E. Variation in double-stranded RNA from isolates of Pyricularia oryzae. Phytopathology 1986, 76, 674–678. [Google Scholar] [CrossRef]
- Cole, T.E.; McLler, B.M.; Hong, Y.; Brasier, C.M.; Buck, K.W. Complexity of Virus-like Double-stranded RN A Elements in a Diseased Isolate of the Dutch Elm Disease Fungus, Ophiostoma novo-ulmi. J. Phytopathol. 1998, 146, 593–598. [Google Scholar] [CrossRef]
- Zhu, Y.; Lu, A.; Wang, Z.; Lu, Q.; Yu, C.; Liu, H. Molecular characterization of a novel endornavirus isolated from Ophiostoma bicolor associated with bark beetles. Arch. Virol. 2022, 167, 2839–2843. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Dover, S.L.; Cole, T.E.; Brasier, C.M.; Buck, K.W. Multiple Mitochondrial Viruses in an Isolate of the Dutch Elm Disease Fungus Ophiostoma Novo-Ulmi. Virology 1999, 258, 118–127. [Google Scholar] [CrossRef][Green Version]
- Honda, S.; Eusebio-Cope, A.; Miyashita, S.; Yokoyama, A.; Aulia, A.; Shahi, S.; Kondo, H.; Suzuki, N. Establishment of Neurospora crassa as a model organism for fungal virology. Nat. Commun. 2020, 11, 5627. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhong, J.; Hu, Y.; Da Gao, B. A novel nonsegmented double-stranded RNA mycovirus identified in the phytopathogenic fungus Nigrospora oryzae shows similarity to partitivirus-like viruses. Arch. Virol. 2016, 161, 229–232. [Google Scholar] [CrossRef]
- Zhong, J.; Zhao, S.Q.; Li, G.F.; Pang, X.D.; Deng, X.J.; Zhu, H.J.; Da Gao, B.; Zhou, Q. A novel fusarivirus isolated from the phytopathogenic fungus Nigrospora oryzae. Virus Genes 2016, 52, 891–895. [Google Scholar] [CrossRef]
- Liu, H.; Liu, R.; Li, C.X.; Wang, H.; Zhu, H.J.; Gao, B.D.; Zhou, Q.; Zhong, J. A victorivirus and two novel mitoviruses co-infected the plant pathogen Nigrospora oryzae. Viruses 2019, 11, 83. [Google Scholar] [CrossRef][Green Version]
- Zhong, J.; Sui, W.W.; Li, P.; Tang, Q.J.; Liu, T.B.; Xiao, Y.S. Characterization of a novel victorivirus from Nigrospora chinensis, a fungus isolated from tobacco. Arch. Virol. 2022, 167, 2851–2855. [Google Scholar] [CrossRef]
- Armengol, J.; Alaniz, S.; Vicent, A.; Beltrán, R.; Abad-Campos, P.; Pérez-Sierra, A.; García-Jiménez, J.; Salem, I.B.; Souli, M.; Boughalleb, N. Effect of dsRNA on growth rate and reproductive potential of Monosporascus cannonballus. Fungal Biol. 2011, 115, 236–244. [Google Scholar] [CrossRef]
- Sahin, E.; Akata, I. Viruses infecting macrofungi. Virusdisease 2018, 29, 1–18. [Google Scholar] [CrossRef]
- De Rezende, R.R.; de Oliveira Souza, F.; Leal, L.L.; Morgan, T.; Bermudez, J.M.M.; Cascardo, R.d.S.; de Barros, D.R.; Alfenas-Zerbini, P. Characterization of a new mitovirus infecting the phytopathogenic fungus Microdochium albescens. Arch. Virol. 2021, 166, 3455–3459. [Google Scholar] [CrossRef]
- Chen, F.; Pu, Z.; Ni, H.; Wang, Y.; Yan, B. Multiple mycoviruses identified in Pestalotiopsis spp. from Chinese bayberry. Virol. J. 2021, 18, 43. [Google Scholar] [CrossRef]
- Zhou, L.; Li, X.; Kotta-Loizou, I.; Dong, K.; Li, S.; Ni, D.; Hong, N.; Wang, G.; Xu, W. A mycovirus modulates the endophytic and pathogenic traits of a plant associated fungus. ISME J. 2021, 15, 1893–1906. [Google Scholar] [CrossRef]
- Velasco, L.; Arjona-Girona, I.; Cretazzo, E.; López-Herrera, C. Viromes in Xylariaceae fungi infecting avocado in Spain. Virology 2019, 532, 11–21. [Google Scholar] [CrossRef]
- Velasco, L.; López-Herrera, C.; Cretazzo, E. Two novel partitiviruses that accumulate differentially in Rosellinia necatrix and Entoleuca sp. infecting avocado. Virus Res. 2020, 285, 198020. [Google Scholar] [CrossRef]
- Arjona-López, J.M.; López-Herrera, C.J. Entoleuca sp. infected by mycoviruses as potential biocontrol agents of avocado white root rot. Eur. J. Plant Pathol. 2021, 159, 409–420. [Google Scholar] [CrossRef]
- Chiba, S.; Salaipeth, L.; Lin, Y.-H.; Sasaki, A.; Kanematsu, S.; Suzuki, N. A novel bipartite double-stranded RNA mycovirus from the white root rot fungus Rosellinia necatrix: Molecular and biological characterization, taxonomic considerations, and potential for biological control. J. Virol. 2009, 83, 12801–12812. [Google Scholar] [CrossRef][Green Version]
- Lin, Y.-H.; Chiba, S.; Tani, A.; Kondo, H.; Sasaki, A.; Kanematsu, S.; Suzuki, N. A novel quadripartite dsRNA virus isolated from a phytopathogenic filamentous fungus, Rosellinia necatrix. Virology 2012, 426, 42–50. [Google Scholar] [CrossRef][Green Version]
- Arjona-López, J.M.; Telengech, P.; Suzuki, N.; López-Herrera, C.J. Coinfection of Rosellinia necatrix by a partitivirus and a virga-like virus is associated with hypovirulence. Eur. J. Plant Pathol. 2020, 158, 111–119. [Google Scholar] [CrossRef]
- Sasaki, A.; Nakamura, H.; Suzuki, N.; Kanematsu, S. Characterization of a new megabirnavirus that confers hypovirulence with the aid of a co-infecting partitivirus to the host fungus, Rosellinia necatrix. Virus Res. 2016, 219, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Arjona-López, J.M.; Telengech, P.; Suzuki, N.; López-Herrera, C.J. A moderate level of hypovirulence conferred by a hypovirus in the avocado white root rot fungus, Rosellinia necatrix. Fungal Biol. 2021, 125, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Kanematsu, S.; Arakawa, M.; Oikawa, Y.; Onoue, M.; Osaki, H.; Nakamura, H.; Ikeda, K.; Kuga-Uetake, Y.; Nitta, H.; Sasaki, A.; et al. A Reovirus Causes Hypovirulence of Rosellinia necatrix. Phytopathology 2004, 94, 561–568. [Google Scholar] [CrossRef][Green Version]
- Yaegashi, H.; Kanematsu, S. Natural infection of the soil-borne fungus Rosellinia necatrix with novel mycoviruses under greenhouse conditions. Virus Res. 2016, 219, 83–91. [Google Scholar] [CrossRef]
- Sutela, S.; Forgia, M.; Vainio, E.J.; Chiapello, M.; Daghino, S.; Vallino, M.; Martino, E.; Girlanda, M.; Perotto, S.; Turina, M. The virome from a collection of endomycorrhizal fungi reveals new viral taxa with unprecedented genome organization. Virus Evol. 2020, 6, veaa076. [Google Scholar] [CrossRef]
- Ren, P.; Rajkumar, S.S.; Zhang, T.; Sui, H.; Masters, P.S.; Martinkova, N.; Kubátová, A.; Pikula, J.; Chaturvedi, S.; Chaturvedi, V. A common partitivirus infection in United States and Czech Republic isolates of bat white-nose syndrome fungal pathogen Pseudogymnoascus destructans. Sci. Rep. 2020, 10, 13893. [Google Scholar] [CrossRef]
- Galán-Cubero, R.; Córdoba, L.; Rodríguez-Romero, J.; Chiapello, M.; Turina, M.; Ayllón, M.A. Molecular Data of a Novel Penoulivirus Associated with the Plant-Pathogenic Fungus Erysiphe necator. Phytopathology 2022, 112, 1587–1591. [Google Scholar] [CrossRef]
- Pandey, B.; Naidu, R.; Grove, G. Detection and analysis of mycovirus-related RNA viruses from grape powdery mildew fungus Erysiphe necator. Arch. Virol. 2018, 163, 1019–1030. [Google Scholar] [CrossRef]
- Botella, L.; Hantula, J. Description, distribution, and relevance of viruses of the forest pathogen Gremmeniella abietina. Viruses 2018, 10, 654. [Google Scholar] [CrossRef][Green Version]
- Botella, L.; Tuomivirta, T.T.; Vervuurt, S.; Diez, J.J.; Hantula, J. Occurrence of two different species of mitoviruses in the European race of Gremmeniella abietina var. abietina, both hosted by the genetically unique Spanish population. Fungal Biol. 2012, 116, 872–882. [Google Scholar] [CrossRef]
- Schoebel, C.N.; Zoller, S.; Rigling, D. Detection and genetic characterisation of a novel mycovirus in Hymenoscyphus fraxineus, the causal agent of ash dieback. Infect. Genet. Evol. 2014, 28, 78–86. [Google Scholar] [CrossRef]
- Shamsi, W.; Kondo, H.; Ulrich, S.; Rigling, D.; Prospero, S. Novel RNA viruses from the native range of Hymenoscyphus fraxineus, the causal fungal agent of ash dieback. Virus Res. 2022, 320, 198901. [Google Scholar] [CrossRef]
- Castro, M.; Kramer, K.; Valdivia, L.; Ortiz, S.; Castillo, A. A double-stranded RNA mycovirus confers hypovirulence-associated traits to Botrytis cinerea. FEMS Microbiol. Lett. 2003, 228, 87–91. [Google Scholar] [CrossRef]
- Howitt, R.L.J.; Beever, R.E.; Pearson, M.N.; Forster, R.L.S. Genome characterization of a flexuous rod-shaped mycovirus, Botrytis virus X, reveals high amino acid identity to genes from plant ‘potex-like’ viruses. Arch. Virol. 2006, 151, 563–579. [Google Scholar] [CrossRef]
- Pearson, M.N.; Bailey, A.M. Viruses of botrytis. In Advances in Virus Research; Academic Press: New York, NY, USA, 2013; Volume 86, pp. 249–272. [Google Scholar]
- Donaire, L.; Pagán, I.; Ayllón, M.A. Characterization of Botrytis cinerea negative-stranded RNA virus 1, a new mycovirus related to plant viruses, and a reconstruction of host pattern evolution in negative-sense ssRNA viruses. Virology 2016, 499, 212–218. [Google Scholar] [CrossRef]
- Hao, F.; Wu, M.; Li, G. Molecular characterization and geographic distribution of a mymonavirus in the population of Botrytis cinerea. Viruses 2018, 10, 432. [Google Scholar] [CrossRef][Green Version]
- Potgieter, C.A.; Castillo, A.; Castro, M.; Cottet, L.; Morales, A. A wild-type Botrytis cinerea strain co-infected by double-stranded RNA mycoviruses presents hypovirulence-associated traits. Virol. J. 2013, 10, 220. [Google Scholar] [CrossRef][Green Version]
- Pi-Fang, T.; Pearson, M.N.; Beever, R.E. Mycoviruses in Monilinia fructicola. Mycol. Res. 2004, 108, 907–912. [Google Scholar]
- De Miccolis Angelini, R.M.; Raguseo, C.; Rotolo, C.; Gerin, D.; Faretra, F.; Pollastro, S. The Mycovirome in a Worldwide Collection of the Brown Rot Fungus Monilinia fructicola. J. Fungi 2022, 8, 481. [Google Scholar] [CrossRef]
- Hamid, M.; Xie, J.; Wu, S.; Maria, S.; Zheng, D.; Assane Hamidou, A.; Wang, Q.; Cheng, J.; Fu, Y.; Jiang, D. A Novel Deltaflexivirus that Infects the Plant Fungal Pathogen, Sclerotinia sclerotiorum, Can Be Transmitted Among Host Vegetative Incompatible Strains. Viruses 2018, 10, 295. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, K.; Zheng, D.; Cheng, J.; Chen, T.; Fu, Y.; Jiang, D.; Xie, J. Characterization of a novel Sclerotinia sclerotiorum RNA virus as the prototype of a new proposed family within the order Tymovirales. Virus Res. 2016, 219, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Mu, F.; Jia, J.; Xue, Y.; Jiang, D.; Fu, Y.; Cheng, J.; Lin, Y.; Xie, J. Characterization of a novel botoulivirus isolated from the phytopathogenic fungus Sclerotinia sclerotiorum. Arch. Virol. 2021, 166, 2859–2863. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wei, D.; Jiang, D.; Fu, Y.; Li, G.; Ghabrial, S.; Peng, Y. Characterization of debilitation-associated mycovirus infecting the plant-pathogenic fungus Sclerotinia sclerotiorum. J. Gen. Virol. 2006, 87, 241–249. [Google Scholar] [CrossRef]
- Liu, L.; Xie, J.; Cheng, J.; Fu, Y.; Li, G.; Yi, X.; Jiang, D. Fungal negative-stranded RNA virus that is related to bornaviruses and nyaviruses. Proc. Natl. Acad. Sci. USA 2014, 111, 12205–12210. [Google Scholar] [CrossRef][Green Version]
- Zhong, J.; Chen, D.; Zhu, H.J.; Gao, B.D.; Zhou, Q. Hypovirulence of Sclerotium rolfsii caused by associated RNA mycovirus. Front. Microbiol. 2016, 7, 1798. [Google Scholar] [CrossRef][Green Version]
- Zhu, J.Z.; Zhu, H.J.; Gao, B.D.; Zhou, Q.; Zhong, J. Diverse, novel mycoviruses from the virome of a hypovirulent Sclerotium rolfsii strain. Front. Plant Sci. 2018, 9, 1738. [Google Scholar] [CrossRef][Green Version]
- Sahin, E.; Keskin, E.; Akata, I. Full-length genome characterization of a new fusagravirus hosted by the spring orange peel fungus Caloscypha fulgens. Acta Virol. 2021, 65, 313–319. [Google Scholar] [CrossRef]
- Sahin, E.; Keskin, E.; Akata, I. Molecular characterization of a novel partitivirus hosted by the false morel mushroom Gyromitra esculenta. Arch. Virol. 2021, 166, 1247–1251. [Google Scholar] [CrossRef]
- Sahin, E.; Keskin, E.; Akata, I. The unique genome organization of two novel fusariviruses hosted by the true morel mushroom Morchella esculenta. Virus Res. 2021, 302, 198486. [Google Scholar] [CrossRef]
- Sahin, E.; Akata, I.; Keskin, E. Novel and divergent bipartite mycoviruses associated with the ectomycorrhizal fungus Sarcosphaera coronaria. Virus Res. 2020, 286, 198071. [Google Scholar] [CrossRef]
- Sahin, E.; Ozbey Saridogan, B.G.; Keskin, E.; Akata, I. Identification and complete genome sequencing of a novel betapartitivirus naturally infecting the mycorrhizal desert truffle Terfezia claveryi. Virus Genes 2023, 59, 254–259. [Google Scholar] [CrossRef]
- Sahin, E.; Akata, I. Complete genome sequence of a novel mitovirus from the ectomycorrhizal fungus Geopora sumneriana. Arch. Virol. 2019, 164, 2853–2857. [Google Scholar] [CrossRef]
- Sahin, E.; Keskin, E.; Akata, I. Novel and diverse mycoviruses co-inhabiting the hypogeous ectomycorrhizal fungus Picoa juniperi. Virology 2021, 552, 10–19. [Google Scholar] [CrossRef]
- Stielow, B.; Klenk, H.-P.; Winter, S.; Menzel, W. A novel Tuber aestivum (Vittad.) mitovirus. Arch. Virol. 2011, 156, 1107–1110. [Google Scholar] [CrossRef]
- Stielow, B.; Menzel, W. Complete nucleotide sequence of TaV1, a novel totivirus isolated from a black truffle ascocarp (Tuber aestivum Vittad.). Arch. Virol. 2010, 155, 2075–2078. [Google Scholar] [CrossRef]
- Riccioni, C.; Rubini, A.; Belfiori, B.; Passeri, V.; Paolocci, F.; Arcioni, S. Tmt1: The first LTR-retrotransposon from a Tuber spp. Curr. Genet. 2008, 53, 23–34. [Google Scholar] [CrossRef]
- Taylor, D.J.; Ballinger, M.J.; Bowman, S.M.; Bruenn, J.A. Virus-host co-evolution under a modified nuclear genetic code. PeerJ 2013, 1, e50. [Google Scholar] [CrossRef][Green Version]
- Pospíšek, M.; Palkova, Z.; Korb, J.; Vaněk, D. Isolation and characterization of a new dsRNA virus from Wickerhamia fluorescens. Folia Microbiol. 1996, 41, 223–227. [Google Scholar] [CrossRef]
- Khan, H.A.; Kondo, H.; Shahi, S.; Bhatti, M.F.; Suzuki, N. Identification of novel totiviruses from the ascomycetous fungus Geotrichum candidum. Arch. Virol. 2022, 167, 2833–2838. [Google Scholar] [CrossRef]
- Matte, O.; Chabalier, C.; Ratomahenina, R.; Bossy, J.; Galzy, P. Isolation of a double-stranded RNA and a virus-like particle from Geotrichum candidum. J. Basic Microbiol. 1991, 31, 447–452. [Google Scholar] [CrossRef]
- Pospíšek, M.; Palková, Z.; Janderová, B.; Korb, J. Isolation and characterization of the dsRNA virus from the yeast Endomyces magnusii. FEMS Microbiol. Lett. 1994, 116, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Groves, D.P.; Clare, J.J.; Oliver, S.G. Isolation and characterisation of a double-stranded RNA virus-like particle from the yeast Yarrowia lipolytica. Curr. Genet. 1983, 7, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Abu-Mejdad, N.M.J.A.; Al-Badran, A.I.; Al-Saadoon, A.H.; Minati, M.H. A new report on gene expression of three killer toxin genes with antimicrobial activity of two killer toxins in Iraq. Bull. Natl. Res. Cent. 2020, 44, 162. [Google Scholar] [CrossRef]
- Lee, M.D.; Creagh, J.W.; Fredericks, L.R.; Crabtree, A.M.; Patel, J.S.; Rowley, P.A. The Characterization of a Novel Virus Discovered in the Yeast Pichia membranifaciens. Viruses 2022, 14, 594. [Google Scholar] [CrossRef]
- Forbes, E.M.; Nieduszynska, S.R.; Brunton, F.K.; Gibson, J.; Glover, L.A.; Stansfield, I. Control of gag-pol gene expression in the Candida albicans retrotransposon Tca2. BMC Mol. Biol. 2007, 8, 94. [Google Scholar] [CrossRef][Green Version]
- Schmitt, M.J.; Breinig, F. Yeast viral killer toxins: Lethality and self-protection. Nat. Rev. Microbiol. 2006, 4, 212–221. [Google Scholar] [CrossRef]
- Wickner, R.B.; Fujimura, T.; Esteban, R. Viruses and Prions of Saccharomyces cerevisiae. In Advances in Virus Research; Academic Press: New York, NY, USA, 2013; Volume 86, pp. 1–36. [Google Scholar]
- Rodríguez-Cousiño, N.; Gómez, P.; Esteban, R. Variation and distribution of LA helper totiviruses in Saccharomyces sensu stricto yeasts producing different killer toxins. Toxins 2017, 9, 313. [Google Scholar] [CrossRef][Green Version]
- Müller, F.; Brühl, K.-H.; Freidel, K.; Kowallik, K.V.; Ciriacy, M. Processing of TY1 proteins and formation of Ty1 virus-like particles in Saccharomyces cerevisiae. Mol. Gener. Genet. 1987, 207, 421–429. [Google Scholar] [CrossRef]
- Schmitt, M.J.; Neuhausen, F. Killer toxin-secreting double-stranded RNA mycoviruses in the yeasts Hanseniaspora uvarum and Zygosaccharomyces bailii. J. Virol. 1994, 68, 1765–1772. [Google Scholar] [CrossRef][Green Version]
- Depierreux, D.; Vong, M.; Nibert, M.L. Nucleotide sequence of Zygosaccharomyces bailii virus Z: Evidence for+ 1 programmed ribosomal frameshifting and for assignment to family Amalgaviridae. Virus Res. 2016, 217, 115–124. [Google Scholar] [CrossRef][Green Version]
- Schmitt, M.J.; Poravou, O.; Trenz, K.; Rehfeldt, K. Unique double-stranded RNAs responsible for the anti-Candida activity of the yeast Hanseniaspora uvarum. J. Virol. 1997, 71, 8852–8855. [Google Scholar] [CrossRef][Green Version]
- Pfeiffer, I.; Farkas, Z.; Golubev, W.I. dsRNA viruses in Nadsonia fulvescens. J. Gen. Appl. Microbiol. 2004, 50, 97–100. [Google Scholar] [CrossRef][Green Version]
- Crucitti, D.; Chiapello, M.; Oliva, D.; Forgia, M.; Turina, M.; Carimi, F.; La Bella, F.; Pacifico, D. Identification and molecular characterization of novel mycoviruses in Saccharomyces and non-Saccharomyces yeasts of oenological interest. Viruses 2021, 14, 52. [Google Scholar] [CrossRef]
- Tavantzis, S.M.; Romaine, C.P.; Smith, S.H. Purification and partial characterization of a bacilliform virus from Agaricus bisporus: A single-stranded RNA mycovirus. Virology 1980, 105, 94–102. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Park, S.-H.; Yie, S.-W.; Kim, K.-H. RT-PCR detection of dsRNA Mycoviruses infecting Pleurotus ostreatus and Agaricus blazei Murrill. Plant Pathol. J. 2005, 21, 343–348. [Google Scholar] [CrossRef]
- Deakin, G.; Dobbs, E.; Bennett, J.M.; Jones, I.M.; Grogan, H.M.; Burton, K.S. Multiple viral infections in Agaricus bisporus-Characterisation of 18 unique RNA viruses and 8 ORFans identified by deep sequencing. Sci. Rep. 2017, 7, 2469. [Google Scholar] [CrossRef][Green Version]
- Heinze, C. A novel mycovirus from Clitocybe odora. Arch. Virol. 2012, 157, 1831–1834. [Google Scholar] [CrossRef]
- Shamoun, S.; Varga, A.; Valverde, R.; Ramsfield, T.; Sumampong, G.; Elliott, M.; Masri, S.; James, D. Identification and molecular characterization of a new double-stranded RNA virus infecting Chondrostereum purpureum. Can. J. Plant Pathol. 2008, 30, 604–613. [Google Scholar] [CrossRef]
- Sahin, E.; Akata, I. Full-length genome characterization of a novel alphapartitivirus detected in the ectomycorrhizal fungus Hygrophorus penarioides. Virus Genes 2021, 57, 94–99. [Google Scholar] [CrossRef]
- Sahin, E.; Akata, I.; Keskin, E. Molecular characterization of a new endornavirus inhabiting the ectomycorrhizal fungus Hygrophorus penarioides. Braz. J. Microbiol. 2021, 52, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- Akata, I.; Edis, G.; Keskin, E.; Sahin, E. Diverse partitiviruses hosted by the ectomycorrhizal agaric Hebeloma mesophaeum and the natural transmission of a partitivirus between phylogenetically distant, sympatric fungi. Virology 2023, 581, 63–70. [Google Scholar] [CrossRef]
- Sahin, E.; Keskin, E.; Akata, I. Molecular characterization of the complete genome of a novel partitivirus hosted by the saprobic mushroom Leucocybe candicans. Arch. Microbiol. 2021, 203, 5825–5830. [Google Scholar] [CrossRef] [PubMed]
- Reyes, B.M.D.; Fonseca, P.L.C.; Heming, N.M.; de Amorim Conceição, L.B.; de Souza Nascimento, K.T.; Gramacho, K.P.; Arevalo-Gardini, E.; Pirovani, C.P.; Aguiar, E.R.G.R. Characterization of the microbiota dynamics associated with Moniliophthora roreri, causal agent of cocoa frosty pod rot disease, reveals new viral species. Front. Microbiol. 2022, 13, 1053562. [Google Scholar] [CrossRef] [PubMed]
- Linnakoski, R.; Sutela, S.; Coetzee, M.P.; Duong, T.A.; Pavlov, I.N.; Litovka, Y.A.; Hantula, J.; Wingfield, B.D.; Vainio, E.J. Armillaria root rot fungi host single-stranded RNA viruses. Sci. Rep. 2021, 11, 7336. [Google Scholar] [CrossRef]
- Tonka, T.; Walterová, L.; Hejna, O.; Čurn, V. Molecular characterization of a ssRNA mycovirus isolated from the forest pathogenic fungus Armillaria ostoyae. Acta virologica 2022, 66, 290–294. [Google Scholar] [CrossRef]
- Zhao, Y.-j.; Shirouzu, T.; Chiba, Y.; Hosaka, K.; Moriyama, H.; Urayama, S.-i.; Hagiwara, D. Identification of novel RNA mycoviruses from wild mushroom isolates in Japan. Virus Res. 2023, 325, 199045. [Google Scholar] [CrossRef]
- Magae, Y.; Sunagawa, M. Characterization of a mycovirus associated with the brown discoloration of edible mushroom, Flammulina velutipes. Virol. J. 2010, 7, 342. [Google Scholar] [CrossRef][Green Version]
- Yu, H.J.; Lim, D.; Lee, H.-S. Characterization of a novel single-stranded RNA mycovirus in Pleurotus ostreatus. Virology 2003, 314, 9–15. [Google Scholar] [CrossRef][Green Version]
- Ro, H.-S.; Kang, E.-J.; Yu, J.-S.; Lee, T.-S.; Lee, C.-W.; Lee, H.-S. Isolation and characterization of a novel mycovirus, PeSV, in Pleurotus eryngii and the development of a diagnostic system for it. Biotechnol. Lett. 2007, 29, 129–135. [Google Scholar] [CrossRef]
- Li, X.; Li, S.; Yin, W.; Sossah, F.L.; Song, B.; Li, Y.; Zhang, B. Complete genome sequence of a novel mycovirus from Pleurotus citrinopileatus. Arch. Virol. 2023, 168, 66. [Google Scholar] [CrossRef]
- Chen, K.; Liang, P.; Yu, M.; Chang, S. A new double-stranded RNA virus from Volvariella volvacea. Mycologia 1988, 80, 849–853. [Google Scholar] [CrossRef]
- Barroso, G.; Labarère, J. Evidence for viral and naked double-stranded RNAs in the basidiomycete Agrocybe aegerita. Curr. Genet. 1990, 18, 231–237. [Google Scholar] [CrossRef]
- Li, X.; Sui, K.; Xie, J.; Hai, D.; Yin, W.; Sossah, F.L.; Jiang, D.; Song, B.; Li, Y. Molecular characterization of a novel fusarivirus infecting the edible fungus Auricularia heimuer. Arch. Virol. 2020, 165, 2689–2693. [Google Scholar] [CrossRef]
- Li, X.; Liu, Q.; Li, S.; Sossah, F.L.; Han, X.; Zhu, G.; Li, Y.; Li, C.; Fu, Y. A novel fungal negative-stranded RNA virus related to mymonaviruses in Auricularia heimuer. Arch. Virol. 2022, 167, 2223–2227. [Google Scholar] [CrossRef]
- Li, X.; Xie, J.; Hai, D.; Sui, K.; Yin, W.; Sossah, F.L.; Jiang, D.; Song, B.; Li, Y. Molecular characteristics of a novel ssRNA virus isolated from Auricularia heimuer in China. Arch. Virol. 2020, 165, 1495–1499. [Google Scholar] [CrossRef]
- Peter, M.; Courty, P.E.; Kohler, A.; Delaruelle, C.; Martin, D.; Tagu, D.; Frey-Klett, P.; Duplessis, S.; Chalot, M.; Podila, G. Analysis of expressed sequence tags from the ectomycorrhizal basidiomycetes Laccaria bicolor and Pisolithus microcarpus. New Phytol. 2003, 159, 117–129. [Google Scholar] [CrossRef]
- Cao, C.; Li, H.; Jones, M.G.; Wylie, S.J. Challenges to elucidating how endornaviruses influence fungal hosts: Creating mycovirus-free isogenic fungal lines and testing them. J. Virol. Methods 2019, 274, 113745. [Google Scholar] [CrossRef]
- Cao, C.T.; Derbyshire, M.C.; Regmi, R.; Li, H.; Jones, M.G.; Wylie, S.J. Small RNA Analyses of a Ceratobasidium Isolate Infected with Three Endornaviruses. Viruses 2022, 14, 2276. [Google Scholar] [CrossRef]
- Zhong, J.; Chen, C.-Y.; Gao, B.-D. Genome sequence of a novel mycovirus of Rhizoctonia solani, a plant pathogenic fungus. Virus Genes 2015, 51, 167–170. [Google Scholar] [CrossRef]
- Li, Y.; Sun, Y.; Yu, L.; Chen, W.; Liu, H.; Yin, L.; Guang, Y.; Yang, G.; Mo, X. Complete genome sequence of a novel mitovirus from binucleate Rhizoctonia AG-K strain FAS2909W. Arch. Virol. 2022, 167, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Shu, C.; Zhang, M.; Yang, M.; Zhou, E. Molecular characterization of a novel endornavirus conferring hypovirulence in rice sheath blight fungus Rhizoctonia solani AG-1 IA strain GD-2. Viruses 2019, 11, 178. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Abdoulaye, A.H.; Foda, M.F.; Kotta-Loizou, I. Viruses infecting the plant pathogenic fungus Rhizoctonia solani. Viruses 2019, 11, 1113. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Marzano, S.-Y.L.; Domier, L.L. Novel mycoviruses discovered from metatranscriptomics survey of soybean phyllosphere phytobiomes. Virus Res. 2016, 213, 332–342. [Google Scholar] [CrossRef][Green Version]
- Komatsu, A.; Kondo, H.; Sato, M.; Kurahashi, A.; Nishibori, K.; Suzuki, N.; Fujimori, F. Isolation and characterization of a novel mycovirus infecting an edible mushroom, Grifola frondosa. Mycoscience 2019, 60, 211–220. [Google Scholar] [CrossRef]
- Kozlakidis, Z.; Hacker, C.V.; Bradley, D.; Jamal, A.; Phoon, X.; Webber, J.; Brasier, C.M.; Buck, K.W.; Coutts, R.H. Molecular characterisation of two novel double-stranded RNA elements from Phlebiopsis gigantea. Virus Genes 2009, 39, 132–136. [Google Scholar] [CrossRef]
- Drenkhan, T.; Sutela, S.; Veeväli, V.; Vainio, E.J. Phlebiopsis gigantea strains from Estonia show potential as native biocontrol agents against Heterobasidion root rot and contain diverse dsRNA and ssRNA viruses. Biol. Control. 2022, 167, 104837. [Google Scholar] [CrossRef]
- Akata, I.; Keskin, E.; Sahin, E. Molecular characterization of a new mitovirus hosted by the ectomycorrhizal fungus Albatrellopsis flettii. Arch. Virol. 2021, 166, 3449–3454. [Google Scholar] [CrossRef]
- Vainio, E.J.; Sutela, S. Mixed infection by a partitivirus and a negative-sense RNA virus related to mymonaviruses in the polypore fungus Bondarzewia berkeleyi. Virus Res. 2020, 286, 198079. [Google Scholar] [CrossRef]
- Sutela, S.; Piri, T.; Vainio, E.J. Discovery and community dynamics of novel ssRNA mycoviruses in the conifer pathogen Heterobasidion parviporum. Front. Microbiol. 2021, 12, 770787. [Google Scholar] [CrossRef]
- Vainio, E.J.; Korhonen, K.; Tuomivirta, T.T.; Hantula, J. A novel putative partitivirus of the saprotrophic fungus Heterobasidion ecrustosum infects pathogenic species of the Heterobasidion annosum complex. Fungal Biol. 2010, 114, 955–965. [Google Scholar] [CrossRef]
- Vainio, E.J.; Hantula, J. Taxonomy, biogeography and importance of Heterobasidion viruses. Virus Res. 2016, 219, 2–10. [Google Scholar] [CrossRef]
- Petrzik, K.; Sarkisova, T.; Starý, J.; Koloniuk, I.; Hrabáková, L.; Kubešová, O. Molecular characterization of a new monopartite dsRNA mycovirus from mycorrhizal Thelephora terrestris (Ehrh.) and its detection in soil oribatid mites (Acari: Oribatida). Virology 2016, 489, 12–19. [Google Scholar] [CrossRef][Green Version]
- Teng, L.; Li, X.; Cai, X.; Yang, S.; Liu, H.; Zhang, T. The complete genome sequence of a novel mycovirus in the plant-pathogenic fungus Exobasidium gracile. Arch. Virol. 2022, 167, 1343–1347. [Google Scholar] [CrossRef]
- Zhang, T.; Cai, X.; Teng, L.; Li, X.; Zhong, N.; Liu, H. Molecular characterization of three novel mycoviruses in the plant pathogenic fungus Exobasidium. Virus Res. 2022, 307, 198608. [Google Scholar] [CrossRef]
- Beck, R.J.; Smith, O.P.; Tooley, P.W.; Peterson, G.L.; Bonde, M.R. Characterization of double-stranded RNA from Tilletia indica. Mycologia 1994, 86, 656–659. [Google Scholar] [CrossRef]
- Applen Clancey, S.; Ruchti, F.; LeibundGut-Landmann, S.; Heitman, J.; Ianiri, G. A novel mycovirus evokes transcriptional rewiring in the fungus Malassezia and stimulates beta interferon production in macrophages. Mbio 2020, 11, e01534-20. [Google Scholar] [CrossRef]
- Zhang, T.; Cai, X.; Zeng, Z.; Du, H.; Liu, H. A novel totivirus isolated from the phytopathogenic fungus Rhodosporidiobolus odoratus strain GZ2017. Arch. Virol. 2020, 165, 1911–1914. [Google Scholar] [CrossRef]
- Liu, J.-J.; Xiang, Y.; Sniezko, R.A.; Schoettle, A.W.; Williams, H.; Zamany, A. Characterization of Cronartium ribicola dsRNAs reveals novel members of the family Totiviridae and viral association with fungal virulence. Virol. J. 2019, 16, 118. [Google Scholar] [CrossRef]
- Liu, J.-J.; Chan, D.; Xiang, Y.; Williams, H.; Li, X.-R.; Sniezko, R.A.; Sturrock, R.N. Characterization of five novel mitoviruses in the white pine blister rust fungus Cronartium ribicola. PLoS ONE 2016, 11, e0154267. [Google Scholar] [CrossRef][Green Version]
- Lawrence, G.; Boelen, M.; Pryor, A. Transmission of double-stranded RNAs in flax rust, Melampsora lini. Canad. J. Bot. 1988, 66, 61–66. [Google Scholar] [CrossRef]
- Zheng, L.; Zhao, J.; Liang, X.; Zhuang, H.; Qi, T.; Kang, Z. Complete genome sequence of a novel mitovirus from the wheat stripe rust fungus Puccinia striiformis. Arch. Virol. 2019, 164, 897–901. [Google Scholar] [CrossRef] [PubMed]
- Pryor, A.; Boelen, M. A double-stranded RNA mycovirus from the maize rust Puccinia sorghi. Canad. J. Bot. 1987, 65, 2380–2383. [Google Scholar] [CrossRef]
- Zheng, L.; Lu, X.; Liang, X.; Jiang, S.; Zhao, J.; Zhan, G.; Liu, P.; Wu, J.; Kang, Z. Molecular characterization of novel totivirus-like double-stranded RNAs from Puccinia striiformis f. sp. tritici, the causal agent of wheat stripe rust. Front. Microbiol. 2017, 8, 1960. [Google Scholar] [CrossRef][Green Version]
- Dickinson, M.J.; Pryor, A. Isometric virus-like particles encapsidate the double-stranded RNA found in Puccinia striiformis, Puccinia recondita, and Puccinia sorghi. Canad. J. Bot. 1989, 67, 3420–3425. [Google Scholar] [CrossRef]
- Castillo, A.; Cifuentes, V. Presence of double-stranded RNA and virus-like particles in Phaffia rhodozyma. Curr. Genet. 1994, 26, 364–368. [Google Scholar] [CrossRef]
- Baeza Cancino, M.; Bravo, N.; Sanhueza, M.; Flores, O.; Villarreal, P.; Cifuentes Guzmán, V. Molecular characterization of totiviruses in Xanthophyllomyces dendrorhous. Virol. J. 2012, 9, 140. [Google Scholar] [CrossRef][Green Version]
- Golubev, W.I.; Pfeiffer, I.; Churkina, L.G.; Golubeva, E.W. Double-stranded RNA viruses in a mycocinogenic strain of Cystofilobasidium infirmominiatum. FEMS Yeast Res. 2003, 3, 63–68. [Google Scholar] [CrossRef]
- Gácser, A.; Hamari, Z.; Pfeiffer, I.; Varga, J.; Kevei, F.; Kucsera, J. Genetic diversity in the red yeast Cryptococcus hungaricus and its phylogenetic relationship to some related basidiomycetous yeasts. FEMS Yeast Res. 2001, 1, 213–220. [Google Scholar]
- Golubev, W.I.; Pfeiffer, I.; Golubeva, E. Mycocin production in Trichosporon pullulans populations colonizing tree exudates in the spring. FEMS Microbiol. Ecol. 2002, 40, 151–157. [Google Scholar] [CrossRef]
- Khandjian, E.; Turian, G.; Eisen, H. Characterization of the RNA mycovirus infecting Allomyces arbuscula. J. Gen. Virol. 1977, 35, 415–424. [Google Scholar] [CrossRef]
- Khandjian, E.; Turian, G. In vitro RNA synthesis by double-stranded RNA mycovirus from Allomyces arbuscula. FEMS Microbiol. Lett. 1977, 2, 121–124. [Google Scholar] [CrossRef]
- Papp, T.; Nyilasi, I.; Fekete, C.; Ferenczy, L.; Vágvölgyi, C. Presence of double-stranded RNA and virus-like particles in Rhizopus isolates. Can. J. Microbiol. 2001, 47, 443–447. [Google Scholar] [CrossRef]
- Turina, M.; Ghignone, S.; Astolfi, N.; Silvestri, A.; Bonfante, P.; Lanfranco, L. The virome of the arbuscular mycorrhizal fungus Gigaspora margarita reveals the first report of DNA fragments corresponding to replicating non-retroviral RNA viruses in fungi. Environ. Microbiol. 2018, 20, 2012–2025. [Google Scholar] [CrossRef]
- Ikeda, Y.; Shimura, H.; Kitahara, R.; Masuta, C.; Ezawa, T. A novel virus-like double-stranded RNA in an obligate biotroph arbuscular mycorrhizal fungus: A hidden player in mycorrhizal symbiosis. Mol. Plant Microbe Interact. 2012, 25, 1005–1012. [Google Scholar] [CrossRef][Green Version]
- Kitahara, R.; Ikeda, Y.; Shimura, H.; Masuta, C.; Ezawa, T. A unique mitovirus from Glomeromycota, the phylum of arbuscular mycorrhizal fungi. Arch. Virol. 2014, 159, 2157–2160. [Google Scholar] [CrossRef][Green Version]
- Kartali, T.; Nyilasi, I.; Kocsubé, S.; Patai, R.; Polgár, T.F.; Zsindely, N.; Nagy, G.; Bodai, L.; Lipinszki, Z.; Vágvölgyi, C. Characterization of four novel dsRNA viruses isolated from Mucor hiemalis strains. Viruses 2021, 13, 2319. [Google Scholar] [CrossRef]
- Barata, R.R.; Vianez-Júnior, J.L.; Nunes, M.R. Transcriptomic analysis of Mucor irregularis containing a negative single-stranded RNA mycovirus. Microbiol. Resour. Announc. 2019, 8, e00503-19. [Google Scholar] [CrossRef][Green Version]
- Kartali, T.; Nyilasi, I.; Szabó, B.; Kocsubé, S.; Patai, R.; Polgár, T.F.; Nagy, G.; Vágvölgyi, C.; Papp, T. Detection and molecular characterization of novel dsRNA viruses related to the Totiviridae family in Umbelopsis ramanniana. Front. Cell. Infect. Microbiol. 2019, 9, 249. [Google Scholar] [CrossRef][Green Version]
- Qu, J.; Shi, N.; Yang, G.; Huang, B. Molecular characterization of a novel totivirus infecting the basal fungus Conidiobolus heterosporus. Arch. Virol. 2021, 166, 1801–1804. [Google Scholar] [CrossRef]
- Shi, N.; Xie, T.; Yang, G.; Wang, P.; Huang, B. Molecular characterization of two novel totiviruses coinfecting the basal fungus Conidiobolus adiaeretus. Arch. Virol. 2023, 168, 7. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Hu, F.; Shi, N.; Wang, P.; Huang, B. A novel non-segmented double-stranded RNA virus isolated from the basal fungus Conidiobolus sp. Arch. Virol. 2020, 165, 1919–1923. [Google Scholar] [CrossRef] [PubMed]
- Krupovic, M.; Dolja, V.V.; Koonin, E.V. Plant viruses of the Amalgaviridae family evolved via recombination between viruses with double-stranded and negative-strand RNA genomes. Biol. Direct 2015, 10, 12. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yang, Y.; Gaspard, G.; McMullen, N.; Duncan, R. Polycistronic Genome Segment Evolution and Gain and Loss of FAST Protein Function during Fusogenic Orthoreovirus Speciation. Viruses 2020, 12, 702. [Google Scholar] [CrossRef] [PubMed]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M. ICTV Virus Taxonomy Profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef]
- Chiba, S.; Castón, J.R.; Ghabrial, S.A.; Suzuki, N. ICTV Virus Taxonomy Profile: Quadriviridae. J. Gen. Virol. 2018, 99, 1480–1481. [Google Scholar] [CrossRef]
- Sato, Y.; Miyazaki, N.; Kanematsu, S.; Xie, J.; Ghabrial, S.A.; Hillman, B.I.; Suzuki, N. ICTV Virus Taxonomy Profile: Megabirnaviridae. J. Gen. Virol. 2019, 100, 1269–1270. [Google Scholar] [CrossRef]
- Kotta-Loizou, I.; Castón, J.R.; Coutts, R.H.A.; Hillman, B.I.; Jiang, D.; Kim, D.-H.; Moriyama, H.; Suzuki, N. ICTV Virus Taxonomy Profile: Chrysoviridae. J. Gen. Virol. 2020, 101, 143–144. [Google Scholar] [CrossRef]
- Kotta-Loizou, I.; Coutts, R.H.; Consortium, I.R. ICTV Virus Taxonomy Profile: Polymycoviridae 2022. J. Gen. Virol. 2022, 103, 1747. [Google Scholar] [CrossRef]
- Supyani, S.; Hillman, B.I.; Suzuki, N. Baculovirus expression of the 11 mycoreovirus-1 genome segments and identification of the guanylyltransferase-encoding segment. J. Gen. Virol. 2007, 88, 342–350. [Google Scholar] [CrossRef]
- Lin, Y.H.; Hisano, S.; Yaegashi, H.; Kanematsu, S.; Suzuki, N. A second quadrivirus strain from the phytopathogenic filamentous fungus Rosellinia necatrix. Arch. Virol. 2013, 158, 1093–1098. [Google Scholar] [CrossRef]
- Li, L.; Liu, J.; Xu, A.; Wang, T.; Chen, J.; Zhu, X. Molecular characterization of a trisegmented chrysovirus isolated from the radish Raphanus sativus. Virus Res. 2013, 176, 169–178. [Google Scholar] [CrossRef]
- Valverde, R.A.; Khalifa, M.E.; Okada, R.; Fukuhara, T.; Sabanadzovic, S. ICTV Virus Taxonomy Profile: Endornaviridae. J. Gen. Virol. 2019, 100, 1204–1205. [Google Scholar] [CrossRef]
- Gilbert, K.B.; Holcomb, E.E.; Allscheid, R.L.; Carrington, J.C. Discovery of new mycoviral genomes within publicly available fungal transcriptomic datasets. PLoS ONE 2019, 14, e0219207. [Google Scholar]
- Li, P.; Zhang, H.; Chen, X.; Qiu, D.; Guo, L. Molecular characterization of a novel hypovirus from the plant pathogenic fungus Fusarium graminearum. Virology 2015, 481, 151–160. [Google Scholar] [CrossRef][Green Version]
- Park, D.; Goh, C.; Lee, J.; Sebastiani, F.; Hahn, Y. Identification of Pistacia-associated flexivirus 1, a putative mycovirus of the family Gammaflexiviridae, in the mastic tree (Pistacia lentiscus) transcriptome. Acta Virol. 2020, 64, 28–35. [Google Scholar] [CrossRef]
- Chen, X.; He, H.; Yang, X.; Zeng, H.; Qiu, D.; Guo, L. The complete genome sequence of a novel Fusarium graminearum RNA virus in a new proposed family within the order Tymovirales. Arch. Virol. 2016, 161, 2899–2903. [Google Scholar] [CrossRef]
- Jiāng, D.; Ayllón, M.A.; Marzano, S.-Y.L. ICTV virus taxonomy profile: Mymonaviridae. J. Gen. Virol. 2019, 100, 1343–1344. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Cao, J.; Yin, X.; Guo, Y.; Guo, L.; Wu, H.; Zhang, M. Characterization of a Novel Mycovirus from the Phytopathogenic Fungus Botryosphaeria dothidea. Viruses 2022, 14, 331. [Google Scholar] [CrossRef]
- Llorens, C.; Soriano, B.; Krupovic, M. ICTV Virus Taxonomy Profile: Metaviridae. J. Gen. Virol. 2020, 101, 1131–1132. [Google Scholar] [CrossRef]
- Llorens, C.; Soriano, B.; Krupovic, M. ICTV Virus Taxonomy Profile: Pseudoviridae. J. Gen. Virol. 2021, 102, 1563. [Google Scholar] [CrossRef] [PubMed]
- Dodonova, S.O.; Prinz, S.; Bilanchone, V.; Sandmeyer, S.; Briggs, J.A. Structure of the Ty3/Gypsy retrotransposon capsid and the evolution of retroviruses. Proc. Natl. Acad. Sci. USA 2019, 116, 10048–10057. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ejmal, M.A.; Holland, D.J.; MacDiarmid, R.M.; Pearson, M.N. A novel chrysovirus from a clinical isolate of Aspergillus thermomutatus affects sporulation. PLoS ONE 2018, 13, e0209443. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.; Li, H.; Nguyen, D.; Jones, M.; Wylie, S. Co-Infection with Three Mycoviruses Stimulates Growth of a Monilinia fructicola Isolate on Nutrient Medium, but Does Not Induce Hypervirulence in a Natural Host. Viruses 2019, 11, 89. [Google Scholar] [CrossRef][Green Version]
- Nuss, D.L. Mycoviruses, RNA Silencing, and Viral RNA Recombination. In Advances in Virus Research; Maramorosch, K., Shatkin, A.J., Murphy, F.A., Eds.; Academic Press: New York, NY, USA, 2011; Volume 80, pp. 25–48. [Google Scholar]
- Zhang, D.-X.; Spiering, M.J.; Nuss, D.L. Characterizing the Roles of Cryphonectria parasitica RNA-Dependent RNA Polymerase-Like Genes in Antiviral Defense, Viral Recombination and Transposon Transcript Accumulation. PLoS ONE 2014, 9, e108653. [Google Scholar] [CrossRef][Green Version]
- Segers, G.C.; Zhang, X.; Deng, F.; Sun, Q.; Nuss, D.L. Evidence that RNA silencing functions as an antiviral defense mechanism in fungi. Proc. Natl. Acad. Sci. USA 2007, 104, 12902–12906. [Google Scholar] [CrossRef][Green Version]
- Sun, Q.; Choi, G.H.; Nuss, D.L. A single Argonaute gene is required for induction of RNA silencing antiviral defense and promotes viral RNA recombination. Proc. Natl. Acad. Sci. USA 2009, 106, 17927–17932. [Google Scholar] [CrossRef][Green Version]
- Chang, S.-S.; Zhang, Z.; Liu, Y. RNA Interference Pathways in Fungi: Mechanisms and Functions. Annu. Rev. Microbiol. 2012, 66, 305–323. [Google Scholar] [CrossRef][Green Version]
- Zhao, J.H.; Guo, H.S. RNA silencing: From discovery and elucidation to application and perspectives. J. Integr. Plant Biol. 2022, 64, 476–498. [Google Scholar] [CrossRef]
- Segers, G.C.; van Wezel, R.; Zhang, X.; Hong, Y.; Nuss, D.L. Hypovirus papain-like protease p29 suppresses RNA silencing in the natural fungal host and in a heterologous plant system. Eukaryot. Cell. 2006, 5, 896–904. [Google Scholar] [CrossRef][Green Version]
- Aulia, A.; Hyodo, K.; Hisano, S.; Kondo, H.; Hillman, B.I.; Suzuki, N. Identification of an RNA Silencing Suppressor Encoded by a Symptomless Fungal Hypovirus, Cryphonectria Hypovirus 4. Biology 2021, 10, 100. [Google Scholar] [CrossRef]
- Yaegashi, H.; Yoshikawa, N.; Ito, T.; Kanematsu, S. A mycoreovirus suppresses RNA silencing in the white root rot fungus, Rosellinia necatrix. Virology 2013, 444, 409–416. [Google Scholar] [CrossRef][Green Version]
- Gaitanis, G.; Magiatis, P.; Hantschke, M.; Bassukas, I.D.; Velegraki, A. The Malassezia Genus in Skin and Systemic Diseases. Clin. Microbiol. Rev. 2012, 25, 106–141. [Google Scholar] [CrossRef][Green Version]
- Limon, J.J.; Tang, J.; Li, D.; Wolf, A.J.; Michelsen, K.S.; Funari, V.; Gargus, M.; Nguyen, C.; Sharma, P.; Maymi, V.I. Malassezia is associated with Crohn’s disease and exacerbates colitis in mouse models. Cell Host Microbe 2019, 25, 377–388.e6. [Google Scholar] [CrossRef][Green Version]
- Aykut, B.; Pushalkar, S.; Chen, R.; Li, Q.; Abengozar, R.; Kim, J.I.; Shadaloey, S.A.; Wu, D.; Preiss, P.; Verma, N. The fungal mycobiome promotes pancreatic oncogenesis via activation of MBL. Nature 2019, 574, 264–267. [Google Scholar] [CrossRef]
- Kotta-Loizou, I.; Sipkova, J.; Coutts, R.H. Identification and sequence determination of a novel double-stranded RNA mycovirus from the entomopathogenic fungus Beauveria bassiana. Arch. Virol. 2015, 160, 873–875. [Google Scholar] [CrossRef]
- Chen, B.; Gao, S.; Choi, G.H.; Nuss, D.L. Extensive alteration of fungal gene transcript accumulation and elevation of G-protein-regulated cAMP levels by a virulence-attenuating hypovirus. Proc. Natl. Acad. Sci. USA 1996, 93, 7996–8000. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Choi, G.H.; Chen, B.; Nuss, D.L. Virus-mediated or transgenic suppression of a G-protein alpha subunit and attenuation of fungal virulence. Proc. Natl. Acad. Sci. USA 1995, 92, 305–309. [Google Scholar] [CrossRef][Green Version]
- Gao, S.; Nuss, D.L. Distinct roles for two G protein α subunits in fungal virulence, morphology, and reproduction revealed by targeted gene disruption. Proc. Natl. Acad. Sci. USA 1996, 93, 14122–14127. [Google Scholar] [CrossRef][Green Version]
- Craven, M.; Pawlyk, D.; Choi, G.; Nuss, D. Papain-like protease p29 as a symptom determinant encoded by a hypovirulence-associated virus of the chestnut blight fungus. J. Virol. 1993, 67, 6513–6521. [Google Scholar] [CrossRef][Green Version]
- Shi, L.; Wang, J.; Quan, R.; Yang, F.; Shang, J.; Chen, B. CpATG8, a homolog of yeast autophagy protein ATG8, is required for pathogenesis and hypovirus accumulation in the chest blight fungus. Front. Cell. Infect. Microbiol. 2019, 9, 222. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.-H.; So, K.-K.; Chun, J.; Kim, D.-H. Distinct roles of two DNA methyltransferases from Cryphonectria parasitica in fungal virulence, responses to hypovirus infection, and viral clearance. MBio 2021, 12, e02890-20. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.-M.; Jeon, J.-J.; Yea, S.-J.; Kim, Y.-H.; Yun, S.-H.; Lee, Y.-W.; Kim, K.-H. Double-stranded RNA mycovirus from Fusarium graminearum. Appl. Environ. Microbiol. 2002, 68, 2529–2534. [Google Scholar] [CrossRef][Green Version]
- Lemus-Minor, C.G.; Cañizares, M.C.; García-Pedrajas, M.D.; Pérez-Artés, E. Horizontal and vertical transmission of the hypovirulence-associated mycovirus Fusarium oxysporum f. sp. dianthi virus 1. Eur. J. Plant Pathol. 2019, 153, 645–650. [Google Scholar] [CrossRef]
- Sasaki, A.; Kanematsu, S.; Onoue, M.; Oyama, Y.; Yoshida, K. Infection of Rosellinia necatrix with purified viral particles of a member of Partitiviridae (RnPV1-W8). Arch. Virol. 2006, 151, 697–707. [Google Scholar] [CrossRef]
- Ran, H.; Liu, L.; Li, B.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. Co-infection of a hypovirulent isolate of Sclerotinia sclerotiorum with a new botybirnavirus and a strain of a mitovirus. Virol. J. 2016, 13, 92. [Google Scholar] [CrossRef][Green Version]
- Urayama, S.-i.; Sakoda, H.; Takai, R.; Katoh, Y.; Le, T.M.; Fukuhara, T.; Arie, T.; Teraoka, T.; Moriyama, H. A dsRNA mycovirus, Magnaporthe oryzae chrysovirus 1-B, suppresses vegetative growth and development of the rice blast fungus. Virology 2014, 448, 265–273. [Google Scholar] [CrossRef][Green Version]
- Wu, M.; Zhang, L.; Li, G.; Jiang, D.; Hou, M.; Huang, H.-C. Hypovirulence and double-stranded RNA in Botrytis cinerea. Phytopathology 2007, 97, 1590–1599. [Google Scholar] [CrossRef][Green Version]
- Wu, M.; Zhang, L.; Li, G.; Jiang, D.; Ghabrial, S.A. Genome characterization of a debilitation-associated mitovirus infecting the phytopathogenic fungus Botrytis cinerea. Virology 2010, 406, 117–126. [Google Scholar] [CrossRef][Green Version]
- Zhong, J.; Li, P.; Gao, B.D.; Zhong, S.Y.; Li, X.G.; Hu, Z.; Zhu, J.Z. Novel and diverse mycoviruses co-infecting a single strain of the phytopathogenic fungus Alternaria dianthicola. Front. Cell. Infect. Microbiol. 2022, 12, 1427. [Google Scholar] [CrossRef]
- Tebbi, C.K.; Badiga, A.; Sahakian, E.; Powers, J.J.; Achille, A.N.; Patel, S.; Migone, F. Exposure to a mycovirus containing Aspergillus Flavus reproduces acute lymphoblastic leukemia cell surface and genetic markers in cells from patients in remission and not controls. Cancer Treat. Res. Commun. 2021, 26, 100279. [Google Scholar] [CrossRef] [PubMed]
- Rowley, P.A. The frenemies within: Viruses, retrotransposons and plasmids that naturally infect Saccharomyces yeasts. Yeast 2017, 34, 279–292. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Aitmanaitė, L.; Konovalovas, A.; Medvedevas, P.; Servienė, E.; Serva, S. Specificity Determination in Saccharomyces cerevisiae Killer Virus Systems. Microorganisms 2021, 9, 236. [Google Scholar] [CrossRef]
- Schmitt, M.J.; Klavehn, P.; Wang, J.; Schnig, I.; Tipper, D.J. Cell cycle studies on the mode of action of yeast K28 killer toxin. Microbiology 1996, 142, 2655–2662. [Google Scholar] [CrossRef][Green Version]
- Martinac, B.; Zhu, H.; Kubalski, A.; Zhou, X.; Culbertson, M.; Bussey, H.; Kung, C. Yeast K1 killer toxin forms ion channels in sensitive yeast spheroplasts and in artificial liposomes. Proc. Natl. Acad. Sci. USA 1990, 87, 6228–6232. [Google Scholar] [CrossRef][Green Version]
- Orentaite, I.; Poranen, M.M.; Oksanen, H.M.; Daugelavicius, R.; Bamford, D.H. K2 killer toxin-induced physiological changes in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2016, 16, fow003. [Google Scholar] [CrossRef][Green Version]
- Liu, S.-Q.; Tsao, M. Inhibition of spoilage yeasts in cheese by killer yeast Williopsis saturnus var. saturnus. Int. J. Food Microbiol. 2009, 131, 280–282. [Google Scholar] [CrossRef]
- Mannazzu, I.; Domizio, P.; Carboni, G.; Zara, S.; Zara, G.; Comitini, F.; Budroni, M.; Ciani, M. Yeast killer toxins: From ecological significance to application. Crit. Rev. Biotechnol. 2019, 39, 603–617. [Google Scholar] [CrossRef]
- Hutzler, M.; Riedl, R.; Koob, J.; Jacob, F. Fermentation and spoilage yeasts and their relevance for the beverage industry-a review. Brew Sci. 2012, 65, 33–52. [Google Scholar]
- Mehlomakulu, N.; Setati, M.; Divol, B. Non-Saccharomyces killer toxins: Possible biocontrol agents against Brettanomyces in wine? S. Afr. J. Enol. Vitic. 2015, 36, 94–104. [Google Scholar] [CrossRef][Green Version]
- Santos, A.; Navascués, E.; Bravo, E.; Marquina, D. Ustilago maydis killer toxin as a new tool for the biocontrol of the wine spoilage yeast Brettanomyces bruxellensis. Int. J. Food Microbiol. 2011, 145, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Weiler, F.; Schmitt, M.J. Zygocin, a secreted antifungal toxin of the yeast Zygosaccharomyces bailii, and its effect on sensitive fungal cells. FEMS Yeast Res. 2003, 3, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Bissegger, M.; Rigling, D.; Heiniger, U. Population Structure and Disease Development of Cryphonectria parasitica in European Chestnut Forests in the Presence of Natural Hypovirulence. Phytopathology 1997, 87, 50–59. [Google Scholar] [CrossRef][Green Version]
- Heiniger, U.; Rigling, D. Biological control of chestnut blight in Europe. Annu. Rev. Phytopathol. 1994, 32, 581–599. [Google Scholar] [CrossRef]
- Rigling, D.; Prospero, S. Cryphonectria parasitica, the causal agent of chestnut blight: Invasion history, population biology and disease control. Mol. Plant Pathol. 2018, 19, 7–20. [Google Scholar] [CrossRef][Green Version]
- Milgroom, M.G.; Cortesi, P. Biological control of chestnut blight with hypovirulence: A critical analysis. Annu. Rev. Phytopathol. 2004, 42, 311–338. [Google Scholar] [CrossRef][Green Version]
- Heiniger, U.; Rigling, D. Application of the cryphonectria hypovirus (chv-1) to control the chestnut blight, experience from switzerland. Acta Hortic. 2009, 815, 233–246. [Google Scholar] [CrossRef]
- Prospero, S.; Rigling, D. Using molecular markers to assess the establishment and spread of a mycovirus applied as a biological control agent against chestnut blight. BioControl 2016, 61, 313–323. [Google Scholar] [CrossRef]
- Yu, X.; Li, B.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Extracellular transmission of a DNA mycovirus and its use as a natural fungicide. Proc. Natl. Acad. Sci. USA 2013, 110, 1452–1457. [Google Scholar] [CrossRef][Green Version]
- Refos, J.M.; Vonk, A.G.; Eadie, K.; Lo-Ten-Foe, J.R.; Verbrugh, H.A.; Van Diepeningen, A.D.; Van De Sande, W.W.J. Double-Stranded RNA Mycovirus Infection of Aspergillus fumigatus Is Not Dependent on the Genetic Make-Up of the Host. PLoS ONE 2013, 8, e77381. [Google Scholar] [CrossRef][Green Version]
- Glass, N.L.; Jacobson, D.J.; Shiu, P.K.T. The genetics of hyphal fusion and vegetative incompatibility in filamentous ascomycete fungi. Annu. Rev. Genet. 2000, 34, 165–186. [Google Scholar] [CrossRef] [PubMed][Green Version]
- García-Pedrajas, M.; Cañizares, M.; Sarmiento-Villamil, J.L.; Jacquat, A.G.; Dambolena, J.S. Mycoviruses in biological control: From basic research to field implementation. Phytopathology 2019, 109, 1828–1839. [Google Scholar] [CrossRef] [PubMed]
- Anagnostakis, S.L. Biological control of chestnut blight. Science 1982, 215, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Attanayake, R.N.; Carter, P.A.; Jiang, D.; del Río-Mendoza, L.; Chen, W. Sclerotinia sclerotiorum populations infecting canola from China and the United States are genetically and phenotypically distinct. Phytopathology 2013, 103, 750–761. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kohn, L.M.; Carbone, I.; Anderson, J.B. Mycelial interactions in Sclerotinia sclerotiorum. Exp. Mycol. 1990, 14, 255–267. [Google Scholar] [CrossRef]
- Zhang, D.-X.; Nuss, D.L. Engineering super mycovirus donor strains of chestnut blight fungus by systematic disruption of multilocus vic genes. Proc. Natl. Acad. Sci. USA 2016, 113, 2062–2067. [Google Scholar] [CrossRef][Green Version]
- Hutchison, E.; Brown, S.; Tian, C.; Glass, N.L. Transcriptional profiling and functional analysis of heterokaryon incompatibility in Neurospora crassa reveals that reactive oxygen species, but not metacaspases, are associated with programmed cell death. Microbiology 2009, 155, 3957–3970. [Google Scholar] [CrossRef][Green Version]
- Ikeda, K.; Inoue, K.; Kida, C.; Uwamori, T.; Sasaki, A.; Kanematsu, S.; Park, P. Potentiation of mycovirus transmission by zinc compounds via attenuation of heterogenic incompatibility in Rosellinia necatrix. Appl. Environ. Microbiol. 2013, 79, 3684–3691. [Google Scholar] [CrossRef][Green Version]
- Brusini, J.; Robin, C. Mycovirus transmission revisited by in situ pairings of vegetatively incompatible isolates of Cryphonectria parasitica. J. Virol. Methods 2013, 187, 435–442. [Google Scholar] [CrossRef]
- Yaegashi, H.; Nakamura, H.; Sawahata, T.; Sasaki, A.; Iwanami, Y.; Ito, T.; Kanematsu, S. Appearance of mycovirus-like double-stranded RNAs in the white root rot fungus, Rosellinia necatrix, in an apple orchard. FEMS Microbiol. Ecol. 2013, 83, 49–62. [Google Scholar] [CrossRef][Green Version]
- Bryner, S.F.; Rigling, D. Virulence not only costs but also benefits the transmission of a fungal virus. Evolution 2012, 66, 2540–2550. [Google Scholar] [CrossRef]
- Biella, S.; Smith, M.L.; Aist, J.R.; Cortesi, P.; Milgroom, M.G. Programmed cell death correlates with virus transmission in a filamentous fungus. Proc. R. Soc. Lond. B Biol. Sci. 2002, 269, 2269–2276. [Google Scholar] [CrossRef]
- Shang, J.; Wu, X.; Lan, X.; Fan, Y.; Dong, H.; Deng, Y.; Nuss, D.L.; Chen, B. Large-scale expressed sequence tag analysis for the chestnut blight fungus Cryphonectria parasitica. Fungal Genet. Biol. 2008, 45, 319–327. [Google Scholar] [CrossRef]
- Wu, S.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Ghabrial, S.A.; Xie, J. Virus-mediated suppression of host non-self recognition facilitates horizontal transmission of heterologous viruses. PLoS Pathog. 2017, 13, e1006234. [Google Scholar] [CrossRef][Green Version]
- Sande, W.W.J.; Lo-Ten-Foe, J.R.; Belkum, A.; Netea, M.G.; Kullberg, B.J.; Vonk, A.G. Mycoviruses: Future therapeutic agents of invasive fungal infections in humans? Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 755–763. [Google Scholar] [CrossRef][Green Version]
- Marzano, S.-Y.L.; Hobbs, H.A.; Nelson, B.D.; Hartman, G.L.; Eastburn, D.M.; McCoppin, N.K.; Domier, L.L. Transfection of Sclerotinia sclerotiorum with in vitro transcripts of a naturally occurring interspecific recombinant of Sclerotinia sclerotiorum hypovirus 2 significantly reduces virulence of the fungus. J. Virol. 2015, 89, 5060–5071. [Google Scholar] [CrossRef][Green Version]
- Chen, B.; Choi, G.H.; Nuss, D.L. Attenuation of fungal virulence by synthetic infectious hypovirus transcripts. Science 1994, 264, 1762–1764. [Google Scholar] [CrossRef]
- Choi, G.H.; Nuss, D.L. Hypovirulence of chestnut blight fungus conferred by an infectious viral cDNA. Science 1992, 257, 800–803. [Google Scholar] [CrossRef]
- Sasaki, A.; Onoue, M.; Kanematsu, S.; Suzaki, K.; Miyanishi, M.; Suzuki, N.; Nuss, D.L.; Yoshida, K. Extending Chestnut Blight Hypovirus Host Range Within Diaporthales by Biolistic Delivery of Viral cDNA. Mol. Plant Microbe Interact. 2002, 15, 780–789. [Google Scholar] [CrossRef][Green Version]
- Hillman, B.I.; Supyani, S.; Kondo, H.; Suzuki, N. A reovirus of the fungus Cryphonectria parasitica that is infectious as particles and related to the Coltivirus genus of animal pathogens. J. Virol. 2004, 78, 892–898. [Google Scholar] [CrossRef][Green Version]
- Clausen, M.; Kräuter, R.; Schachermayr, G.; Potrykus, I.; Sautter, C. Antifungal activity of a virally encoded gene in transgenic wheat. Nat. Biotechnol. 2000, 18, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Polashock, J.; Bedker, P.; Hillman, B. Movement of a small mitochondrial double-stranded RNA element of Cryphonectria parasitica: Ascospore inheritance and implications for mitochondrial recombination. Mol. Gener. Genet. 1997, 256, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Van Diepeningen, A.D.; Debets, A.J.; Hoekstra, R.F. Dynamics of dsRNA mycoviruses in black Aspergillus populations. Fungal Genet. Biol. 2006, 43, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wu, M.; Zhang, J.; Chen, W.; Li, G.; Yang, L. Sclerotinia minor Endornavirus 1, a Novel Pathogenicity Debilitation-Associated Mycovirus with a Wide Spectrum of Horizontal Transmissibility. Viruses 2018, 10, 589. [Google Scholar] [CrossRef][Green Version]
- Hillman, B.I.; Suzuki, N. Viruses in the chestnut blight fungus. In Advances in Virus Research; Academic Press: New York, NY, USA, 2004; Volume 63, pp. 423–473. [Google Scholar]
- Lee, K.-M.; Cho, W.K.; Yu, J.; Son, M.; Choi, H.; Min, K.; Lee, Y.-W.; Kim, K.-H. A Comparison of Transcriptional Patterns and Mycological Phenotypes following Infection of Fusarium graminearum by Four Mycoviruses. PLoS ONE 2014, 9, e100989. [Google Scholar] [CrossRef]
- Ihrmark, K.; Stenström, E.; Stenlid, J. Double-stranded RNA transmission through basidiospores of Heterobasidion annosum. Mycol. Res. 2004, 108, 149–153. [Google Scholar] [CrossRef]
- Varga, J.; Rinyu, E.; Kevei, É.; Tóth, B.; Kozakiewicz, Z. Double-stranded RNA mycoviruses in species of Aspergillus sections Circumdati and Fumigati. Can. J. Microbiol. 1998, 44, 569–574. [Google Scholar] [CrossRef]
- Morris, T.; Dodds, J. Isolation and analysis of double-stranded RNA from virus-infected plant and fungal tissue. Phytopathology 1979, 69, 854–858. [Google Scholar] [CrossRef][Green Version]
- Okada, R.; Kiyota, E.; Moriyama, H.; Fukuhara, T.; Natsuaki, T. A simple and rapid method to purify viral dsRNA from plant and fungal tissue. J. Gen. Plant Pathol. 2015, 81, 103–107. [Google Scholar] [CrossRef]
- Dawe, A.L.; Nuss, D.L. Hypovirus Molecular Biology: From Koch’s Postulates to Host Self-Recognition Genes that Restrict Virus Transmission. In Advances in Virus Research; Ghabrial, S.A., Ed.; Academic Press: New York, NY, USA, 2013; Volume 86, pp. 109–147. [Google Scholar]
- Liu, R.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. Molecular Characterization of a Novel Positive-Sense, Single-Stranded RNA Mycovirus Infecting the Plant Pathogenic Fungus Sclerotinia sclerotiorum. Viruses 2015, 7, 2470–2484. [Google Scholar] [CrossRef][Green Version]
- Xie, J.; Xiao, X.; Fu, Y.; Liu, H.; Cheng, J.; Ghabrial, S.A.; Li, G.; Jiang, D. A novel mycovirus closely related to hypoviruses that infects the plant pathogenic fungus Sclerotinia sclerotiorum. Virology 2011, 418, 49–56. [Google Scholar] [CrossRef][Green Version]

| Ascomycota | ||||
|---|---|---|---|---|
| Class | Order | Family | Genera | Mycoviruses |
| Arthoniomycetes | Arthoniales | Chrysotrichaceae | Chrysothrix | dsRNA [42] |
| Dothideomycetes | Botryosphaeriales | Aplosporellaceae | Aplosporella | dsRNA [43] |
| Botryosphaeriaceae | Botryosphaeria | dsRNA and +ssRNA [44,45,46,47,48] | ||
| Diplodia | dsRNA and +ssRNA [49,50,51] | |||
| Eutiarosporella | +ssRNA [52] | |||
| Macrophomina | dsRNA, +ssRNA, −ssRNA and ssDNA [53,54,55,56] | |||
| Neofusicoccum | dsRNA and +ssRNA [57,58,59] | |||
| Phaeobotryon | dsRNA [60] | |||
| Phyllostictaceae | Phyllosticta | dsRNA [61] | ||
| Capnodiales | Cladosporiaceae | Cladosporium | +ssRNA, −ssRNA and RT ssRNA [62,63] | |
| Mycosphaerellaceae | Cercospora | +ssRNA and −ssRNA [64] | ||
| Dothistroma | dsRNA [65] | |||
| Mycosphaerella | +ssRNA [52,66] | |||
| Pseudocercospora | dsRNA [66] | |||
| Zymoseptoria | dsRNA and +ssRNA [61,64,67] | |||
| Dothideales | Saccotheciaceae | Aureobasidium | +ssRNA [52] | |
| Mycosphaerellales | Teratosphaeriaceae | Acidomyces | +ssRNA [61] | |
| Hortaea | dsRNA [61] | |||
| Pleosporales | Coniothyriaceae | Coniothyrium | dsRNA and −ssRNA [63,68] | |
| Corynesporascaceae | Corynespora | dsRNA [67,69] | ||
| Cucurbitariaceae | Cucurbitaria | dsRNA [69,70] | ||
| Delitschiaceae | Delitschia | dsRNA [61] | ||
| Didymellaceae | Didymella | dsRNA [71] | ||
| Epicoccum | +ssRNA [63] | |||
| Leptosphaerulina | +ssRNA [63] | |||
| Phoma | dsRNA and +ssRNA [71,72,73,74,75] | |||
| Stagonosporopsis | dsRNA [47,72,73,74,75] | |||
| Leptosphaeriaceae | Leptosphaeria | dsRNA and + ssRNA [76,77,78] | ||
| Lindgomycetaceae | Clohesyomyces | dsRNA [47,61] | ||
| Massarinaceae | Helminthosporium | dsRNA [79] | ||
| Periconiaceae | Periconia | dsRNA and +ssRNA [61,80] | ||
| Pleosporaceae | Alternaria | dsRNA, +ssRNA, and −ssRNA [63,80,81,82,83,84,85,86,87,88] | ||
| Bipolaris | dsRNA and +ssRNA [89,90,91,92] | |||
| Cochliobolus | dsRNA [93] | |||
| Curvularia | dsRNA [94] | |||
| Drechslera | dsRNA [61,95] | |||
| Exserohilum | dsRNA [95,96,97] | |||
| Pleospora | dsRNA [96,97,98] | |||
| Setosphaeria | dsRNA and +ssRNA [97,99,100] | |||
| Stemphylium | +ssRNA [101] | |||
| Eurotiomycetes | Chaetothyriales | Herpotrichiellaceae | Phialophora | dsRNA [81,82,83,84,85,86,87,88,102,103] |
| Eurotiales | Aspergillaceae | Aspergillus | dsRNA and +ssRNA [93,104,105,106,107] | |
| Penicillium | dsRNA, +ssRNA and −ssRNA [15,63,89,90,91,92,108,109] | |||
| Trichocomaceae | Talaromyces | dsRNA [94,110,111,112] | ||
| Onygenales | Ajellomycetaceae | Blastomyces | dsRNA [98,113] | |
| Ascosphaeraceae | Ascosphaera | +ssRNA [97,99,100,114,115] | ||
| Onygenales incertae sedis | Myriodontium | dsRNA [116] | ||
| Lecanoromycetes | Lecanorales | Stereocaulaceae | Lepraria | dsRNA [42,101] |
| Sordariomycetes | Diaporthales | Cryphonectriaceae | Cryphonectria | dsRNA, +ssRNA and −ssRNA [117,118,119,120,121] |
| Endothia | dsRNA [122] | |||
| Diaporthaceae | Diaporthe | dsRNA and +ssRNA [56,123,124] | ||
| Melanconiellaceae | Melanconiella | +ssRNA [125] | ||
| Valsaceae | Cytospora | dsRNA [126] | ||
| Phomopsis | dsRNA and +ssRNA [127,128,129,130] | |||
| Valsa | +ssRNA [131] | |||
| Glomerellales | Glomerellaceae | Colletotrichum | dsRNA and +ssRNA [132,133,134,135,136,137,138] | |
| Plectosphaerellaceae | Sodiomyces | dsRNA and +ssRNA [139] | ||
| Verticillium | dsRNA and +ssRNA [140,141,142,143] | |||
| Hypocreales | Bionectriaceae | Clonostachys | dsRNA [144] | |
| Clavicipitaceae | Atkinsonella | dsRNA [145] | ||
| Epichloë | dsRNA [146,147] | |||
| Metarhizium | dsRNA [148,149,150,151] | |||
| Ustilaginoidea | dsRNA [152,153,154,155,156] | |||
| Cordycipitaceae | Beauveria | dsRNA [61,157,158,159,160] | ||
| Cordyceps | dsRNA [161,162] | |||
| Isaria | dsRNA [163] | |||
| Hypocreaceae | Hypomyces | dsRNA [164] | ||
| Trichoderma | dsRNA and +ssRNA [61,165,166,167,168] | |||
| Fusarium | dsRNA, +ssRNA, −ssRNA, and ssDNA [169,170,171,172,173,174,175] | |||
| Ilyonectria | dsRNA [176,177] | |||
| Nectria | dsRNA [178] | |||
| Rugonectria | dsRNA and +ssRNA [179] | |||
| Thelonectria | dsRNA [180] | |||
| Hypocreales incertae sedis | Acremonium | +ssRNA [63] | ||
| Ophiocordycipitaceae | Ophiocordyceps | +ssRNA [61] | ||
| Tolypocladium | dsRNA [181,182] | |||
| Magnaporthales | Magnaporthaceae | Buergenerula | +ssRNA [183] | |
| Gaeumannomyces | dsRNA and +ssRNA [61,184] | |||
| Magnaporthe | dsRNA and +ssRNA [185,186,187,188] | |||
| Microascales | Ceratocystidaceae | Berkeleyomyces | dsRNA and +ssRNA [189,190] | |
| Ceratocystis | dsRNA and +ssRNA [191] | |||
| Endoconidiophora | dsRNA [192] | |||
| Pyriculariaceae | Pyriculariaceae | dsRNA [193,194] | ||
| Ophiostomatales | Ophiostomataceae | Grosmannia | dsRNA and +ssRNA [61] | |
| Ophiostoma | dsRNA and +ssRNA [195,196,197] | |||
| Sordariales | Sordariaceae | Neurospora | dsRNA and +ssRNA [198] | |
| Togniniales | Togniniaceae | Phaeoacremonium | dsRNA and +ssRNA [63] | |
| Xylariales | Apiosporaceae | Nigrospora | dsRNA and +ssRNA [199,200,201,202] | |
| Diatrypaceae | Monosporascus | dsRNA [203] | ||
| Hypoxylaceae | Annulohypoxylon | +ssRNA [204] | ||
| Microdochiaceae | Microdochium | +ssRNA [205] | ||
| Sporocadaceae | Pestalotiopsis | dsRNA, +ssRNA and −ssRNA [206] | ||
| Pseudopestalotiopsis | dsRNA [207] | |||
| Xylariaceae | Entoleuca | dsRNA and +ssRNA [208,209,210] | ||
| Rosellinia | dsRNA and +ssRNA [18,208,211,212,213,214,215,216,217] | |||
| Leotiomycetes | Leotiomycetes incertae sedis | Myxotrichaceae | Oidiodendron | +ssRNA [70,218] |
| Pseudeurotiaceae | Pseudogymnoascus | dsRNA [219] | ||
| Erysiphales | Erysiphaceae | Erysiphe | dsRNA and +ssRNA [68,220,221] | |
| Podosphaera | dsRNA and +ssRNA [77] | |||
| Helotiales | Godroniaceae | Gremmeniella | dsRNA and +ssRNA [51,61,222,223] | |
| Helotiaceae | Hymenoscyphus | dsRNA, +ssRNA and −ssRNA [121,224,225] | ||
| Mollisiaceae | Loramyces | +ssRNA [61] | ||
| Rutstroemiaceae | Rutstroemia | +ssRNA [61] | ||
| Sclerotiniaceae | Botrytis | dsRNA and +ssRNA [226,227,228,229,230,231] | ||
| Monilinia | dsRNA, +ssRNA and ssDNA [232,233] | |||
| Sclerotinia | dsRNA, +ssRNA, −ssRNA and dsRNA [16,175,234,235,236,237,238] | |||
| Sclerotium | dsRNA and +ssRNA [239,240] | |||
| Thelebolales | Thelebolaceae | Thelebolus | dsRNA [61] | |
| Pezizomycetes | Pezizales | Caloscyphaceae | Caloscypha | dsRNA [241] |
| Discinaceae | Gyromitra | dsRNA [61,242] | ||
| Morchellaceae | Morchella | dsRNA and +ssRNA [61,243] | ||
| Pezizaceae | Peziza | +ssRNA [204] | ||
| Sarcosphaera | dsRNA [244] | |||
| Terfezia | dsRNA [245] | |||
| Pyronemataceae | Geopora | dsRNA and +ssRNA [121,246] | ||
| Picoa | dsRNA and +ssRNA [247] | |||
| Tuberaceae | Tuber | dsRNA, +ssRNA and RT-ssRNA [121,248,249,250] | ||
| Saccharomycetes | Saccharomycetales | Debaryomycetaceae | Scheffersomyces | dsRNA [251] |
| Wickerhamia | dsRNA [252] | |||
| Dipodascaceae | Geotrichum | dsRNA [253,254] | ||
| Magnusiomyces | dsRNA [255] | |||
| Yarrowia | dsRNA [256] | |||
| Phaffomycetaceae | Wickerhamomyces | dsRNA [257] | ||
| Pichiaceae | Pichia | dsRNA [258] | ||
| Saccharomycetaceae | Candida | RT-ssRNA [259] | ||
| Saccharomyces | dsRNA, +ssRNA and RT-ssRNA [260,261,262,263] | |||
| Torulaspora | dsRNA [257] | |||
| Zygosaccharomyces | dsRNA [264,265] | |||
| Saccharomycodaceae | Hanseniaspora | dsRNA [264,266] | ||
| Saccharomycetales incertae sedis | Ambrosiozyma | dsRNA [258] | ||
| Nadsonia | dsRNA [267] | |||
| Starmerella | dsRNA [268] | |||
| Magnaporthe | dsRNA and +ssRNA [185,186,187,188] | |||
| Basidiomycota | ||||
|---|---|---|---|---|
| Class | Order | Family | Genera | Mycoviruses |
| Agaricomycetes | Agaricales | Agaricaceae | Agaricus | dsRNA and +ssRNA [269,270,271] |
| Leucocoprinus | +ssRNA [52] | |||
| Clitocybaceae | Clitocybe | +ssRNA [272] | ||
| Cyphellaceae | Chondrostereum | dsRNA [204,273] | ||
| Hydnangiaceae | Laccaria | +ssRNA [204] | ||
| Hygrophoraceae | Hygrophorus | dsRNA [121,274,275] | ||
| Hymenogastraceae | Hebeloma | dsRNA [276] | ||
| Lyophyllaceae | Leucocybe | dsRNA [277] | ||
| Marasmiaceae | Moniliophthora | +ssRNA [278] | ||
| Nidulariaceae | Cyathus | +ssRNA [52] | ||
| Omphalotaceae | Collybiopsis | +ssRNA [204] | ||
| Lentinula | dsRNA and +ssRNA [204] | |||
| Physalacriaceae | Armillaria | +ssRNA and −ssRNA [121,279,280] | ||
| Flammulina | dsRNA [281,282] | |||
| Pleurotaceae | Pleurotus | dsRNA and +ssRNA [270,283,284,285] | ||
| Pluteaceae | Volvariella | dsRNA [204,286] | ||
| Porotheleaceae | Megacollybia | dsRNA [204] | ||
| Psathyrellaceae | Coprinopsi | dsRNA and +ssRNA [204,281] | ||
| Schizophyllaceae | Schizophyllum | +ssRNA [204] | ||
| Strophariaceae | Agrocybe | dsRNA [281,287] | ||
| Squamanitaceae | Phaeolepiota | +ssRNA [204] | ||
| Auriculariales | Auriculariaceae | Auricularia | dsRNA, +ssRNA and −ssRNA [288,289,290] | |
| Boletales | Boletaceae | Boletus | +ssRNA [204] | |
| Pisolithaceae | Pisolithus | dsRNA [291] | ||
| Cantharellales | Cantharellaceae | Craterellus | +ssRNA [204] | |
| Ceratobasidiaceae | Ceratobasidium | +ssRNA [292,293] | ||
| Rhizoctonia | dsRNA, +ssRNA and −ssRNA [294,295,296,297] | |||
| Thanatephorus | +ssRNA [298] | |||
| Tulasnellaceae | Tulasnella | −ssRNA [218] | ||
| Gloeophyllales | Gloeophyllaceae | Neolentinus | +ssRNA [204] | |
| Hymenochaetales | Hymenochaetaceae | Fomitiporia | +ssRNA [63] | |
| Polyporales | Grifolaceae | Grifola | dsRNA [299] | |
| Phanerochaetaceae | Phlebiopsis | dsRNA and +ssRNA [300,301] | ||
| Russulales | Albatrellaceae | Albatrellopsis | +ssRNA [302] | |
| Bondarzewiaceae | Bondarzewia | dsRNA and −ssRNA [303] | ||
| Heterobasidion | dsRNA and +ssRNA [304,305,306] | |||
| Russulaceae | Lactarius | dsRNA [121] | ||
| Thelephorales | Thelephoraceae | Thelephora | dsRNA [204,307] | |
| Exobasidiomycetes | Exobasidiales | Exobasidiaceae | Exobasidium | dsRNA [304,308,309] |
| Tilletiales | Tilletiaceae | Tilletia | dsRNA [310] | |
| Malasseziomycetes | Malasseziales | Malasseziaceae | Malassezia | dsRNA [311] |
| Microbotryomycetes | Sporidiobolales | Sporidiobolaceae | Rhodosporidiobolus | dsRNA [312] |
| Pucciniales | Coleosporiaceae | Cronartium | dsRNA and +ssRNA [121,313,314] | |
| Melampsoraceae | Melampsora | dsRNA [315] | ||
| Pucciniaceae | Puccinia | dsRNA and +ssRNA [316,317,318,319] | ||
| Uromyces | dsRNA [52] | |||
| Phakopsoraceae | Phakopsora | dsRNA [52] | ||
| Tremellomycetes | Cystofilobasidiales | Mrakiaceae | Phaffia | dsRNA [320,321] |
| Cystofilobasidiaceae | Cystofilobasidium | dsRNA [322] | ||
| Tremellales | Cryptococcaceae | Cryptococcus | dsRNA [323] | |
| Trichosporonales | Trichosporonaceae | Trichosporon | dsRNA [324] | |
| Wallemiomycetes | Wallemiales | Wallemiaceae | Wallemia | dsRNA [98] |
| Blastocladiomycota | ||||
|---|---|---|---|---|
| Class | Order | Family | Genera | Mycoviruses |
| Blastocladiomycetes | Blastocladiales | Blastocladiaceae | Allomyces | dsRNA [2,325,326] |
| Chytridiomycota | ||||
| Class | Order | Family | Genera | Mycoviruses |
| Chytridiomycetes | Chytridiales | Chytridiaceae | Zopfochytrium | dsRNA [2] |
| Cladochytriales | Cladochytriaceae | Cladochytrium | dsRNA [2] | |
| Rhizophydiales | Rhizophydiales incertae sedis | Operculomyces | dsRNA and +ssRNA [2] | |
| Rhizopodaceae | Rhizopus | dsRNA and +ssRNA [2,327] | ||
| Mucoromycota | ||||
| Class | Order | Family | Genera | Mycoviruses |
| Glomeromycetes | Archaeosporales | Geosiphonaceae | Geosiphon | +ssRNA [2] |
| Diversisporales | Gigasporaceae | Gigaspora | dsRNA, +ssRNA and ssDNA [2,52,328] | |
| Glomerales | Glomeraceae | Glomus | dsRNA [329] | |
| Racocetra | dsRNA and +ssRNA [52] | |||
| Rhizophagus | +ssRNA [2,32,330] | |||
| Paraglomerales | Paraglomeraceae | Paraglomus | + ssRNA [52] | |
| Mortierellomycetes | Mortierellales | Mortierellaceae | Dissophora | dsRNA [2] |
| Lobosporangium | dsRNA [2] | |||
| Mortierella | dsRNA, +ssRNA and −ssRNA [2] | |||
| Mucoromycetes | Mucorales | Choanephoraceae | Blakeslea | dsRNA [2] |
| Choanephora | dsRNA and +ssRNA [2] | |||
| Cunninghamellaceae | Absidia | dsRNA [2] | ||
| Mucoraceae | Mucor | dsRNA, −ssRNA and ssDNA [52,53,331,332] | ||
| Phycomycetaceae | Phycomyces | [2] | ||
| Syncephalastraceae | Syncephalastrum | dsRNA [107] | ||
| Umbelopsidomycetes | Umbelopsidales | Umbelopsidaceae | Umbelopsis | dsRNA [2,41,333] |
| Neocallimastigomycota | ||||
| Class | Order | Family | Genera | Mycoviruses |
| Neocallimastigomycetes | Neocallimastigales | Neocallimastigaceae | Anaeromyces | +ssRNA [2] |
| Neocallimastix | dsRNA [2] | |||
| Pecoramyces | ssDNA [52] | |||
| Zoopagomycota | ||||
| Class | Order | Family | Genera | Mycoviruses |
| Entomophthoromycetes | Entomophthorales | Ancylistaceae | Conidiobolus | dsRNA and +ssRNA [2,334,335,336] |
| Entomophthoraceae | Entomophaga | dsRNA [2] | ||
| Entomophthora | +ssRNA [2,52] | |||
| Zoophthora | dsRNA and +ssRNA [2] | |||
| Kickxellomycetes | Kickxellales | Kickxellaceae | Kickxella | dsRNA and +ssRNA [2] |
| Zoopagomycetes | Zoopagales | Piptocephalidaceae | Syncephalis | +ssRNA [2] |
| Families and Genera | Genome Size and Segmentation | Genome Organization | Exemplar Species |
|---|---|---|---|
| Family Alphaflexiviridae Mycovirus associated genera Botrexvirus Sclerodarnavirus | Non-segmented genomes around 5.5 to 9 kb in size. | Botrexvirus: Genome consists of up to 5 ORFs. ORF1: Encodes for an RdRp, Hel, and Mtf. ORF3: Encodes for a CP. All remaining ORFs encode for putative proteins with unknown function. Sclerodarnavirus: Genome consists of a single ORF encoding for an RdRp, Mtf, and Hel. | Botrexvirus: Botrytis virus X Sclerodarnavirus: Sclerotinia sclerotiorum debilitation-associated RNA virus |
| Family Barnaviridae Mycovirus associated genera Barnavirus | Non-segmented with genomes around 4.0 kb in size. | Genome consists of 4 ORFs. ORF1: Encodes for a hypothetical protein with an unknown function. ORF2: Encodes for a putative serine protease. ORF3: Encodes for an RdRp. ORF4: Encodes for a CP. | Mushroom bacilliform virus |
| Family Botourmiaviridae Mycovirus associated genera Botoulivirus Betabotoulivirus Magoulivirus Scleroulivirus Betascleroulivirus Deltascleroulivirus Penoulivirus Rhizoulivirus Betarhizoulivirus | Non-segmented genomes around 3 to 5.3 kbp in size | Genome consists of a single ORF encoding for an RdRp | Botoulivirus: Botrytis cinerea ourmia-like virus 4 Betabotoulivirus: Entoleuca ourmia-like virus 1 Magoulivirus: Magnaporthe oryzae ourmia-like virus 1 Scleroulivirus: Sclerotinia sclerotiorum ourmia-like virus 1 Betascleroulivirus: Botrytis cinerea ourmia-like virus 10 Deltascleroulivirus: Botrytis cinerea ourmia-like virus 5 Penoulivirus: Aspergillus neoniger ourmia-like virus 1 Rhizoulivirus: Rhizoctonia solani ourmia-like virus 1Rs Betarhizoulivirus: Rhizoctonia solani ourmia-like virus 5 |
| Family Deltaflexiviridae Mycovirus associated genera Deltaflexivirus | Non-segmented with genomes around 6 to 8 kbp in size | Genome consists of 1–5 ORFs ORF 1: Encodes for polyprotein with Mt, Gtf, Hel and RdRp domains ORF 2–5: Encode for hypothetical proteins with unknown functions | Sclerotinia sclerotiorum deltaflexivirus 1 |
| Family Endornaviridae Mycovirus associated genera Alphaendornavirus Betaendornavirus | Non segmented genomes around 9.5 to 17.6 kb in size. | Genome consists of a single ORF encoding for a large polyprotein. The polyprotein always contains an RdRp domain, but may also include domains for Hel, Mt, Gtf, CPS, and phytoreo S7 domains. | Alphaendornavirus: Oryza sativa alphaendornavirus Betaendornavirus: Sclerotinia sclerotiorum endornavirus |
| Family Gammaflexiviridae Mycovirus associated genera Mycoflexivirus | Non-segmented genomes around 6.8 to 9.2 kb in size. | Consists of 2–3 ORFs ORF 1: Encodes for a replicase (REP) with an upstream Mtf domain, a Hel domain, and a downstream RdRp domain. ORF 2: Encodes for either a CP or proteins which resemble movement proteins. ORF 3: Encodes for either a CP or a hypothetical protein with an unknown function. | Botrytis virus F |
| Family Hadakaviridae Mycovirus associated genera Hadakavirus | Multisegmented genomes around 14 to 15 kb in total size. | RNA 1: Encodes for an RdRp. RNA3: Encodes for a Mtf. RNA 2, 4–7, 9 and 10: Encode for hypothetical proteins with an unknown function. RNA8: May encode for a C2H2-type zinc finger protein | hadaka virus 1 |
| Family Hypoviridae Mycovirus associated genera Alphahypovirus Betahypovirus Epsilonhypovirus Etahypovirus Thetahypovirus | Non-segmented genomes around 9.1 to 12.7 kb in size | Consist of 1–2 ORFs Encodes for a polyprotein containing RdRp and sometimes protease, Hel, and Gtf domains. Some hypoviruses also have short, internally deleted, defective interfering replicative forms of dsRNA molecules, while others have replicative forms of satellite like RNAs | Alphahypovirus Cryphonectria hypovirus 2 Betahypovirus: Cryphonectria hypovirus 4 Epsilonhypovirus: Agaricus bisporus virus 2 Etahypovirus: Sclerotium rolfsii hypovirus 8 Thetahypovirus: Botrytis cinerea hypovirus 4 |
| Family Mitoviridae Mycovirus associated genera Unuamitovirus Duamitovirus Triamitovirus Kvaramitovirus | Non-segmented genomes around 2.0 kbp to 4.5 kb in size | Consists of 1 ORF encoding for an RdRp | Unuamitovirus: Botrytis cinerea mitovirus 2 Duamitovirus: Alternaria alternata mitovirus 1 Triamitovirus: Rhizoctonia solani mitovirus 30 Kvaramitovirus: Ophiostoma mitovirus 7 |
| Family Narnaviridae Mycovirus associated genera Narnavirus | Non-segmented genomes around 2.3 to 3.6 kb in size | Consists of 1 ORF encoding for an RdRp | Saccharomyces 20S RNA narnavirus |
| Family Yadokariviridae Mycovirus associated genera Alphayadokarivirus Betayadokarivirus | Non-segmented genomes around 3.6 to 6.3 kb in size | Genome consists of a 1–2 ORFs encoding for a polyprotein. Alphayadokarivirus Consist of 1 ORF which encodes for a polyprotein with an RdRp domain, 2A-like self-cleaving peptide and a hypothetical protein domain with an unknown function. Betayadokarivirus Consist of 1 or 2 ORFs encoding for a polyprotein, which may or may not include a 2A-like self-cleaving peptide. The polyprotein contains an RdRp domain (5′ proximal ORF), and a hypothetical protein domain (3′ proximal ORF) with an unknown function. | yado-kari virus 2 |
| Families and Genera | Genome Size and Segmentation | Genome Organization | Exemplar Species |
|---|---|---|---|
| Family Mymonaviridae Mycovirus associated genera Auricularimonavirus Botrytimonavirus Lentimonavirus Penicillimonavirus Sclerotimonavirus | Non-segmented genomes around 6 to 10 kb in size | Consists of 1–7 ORFs All mymonaviruses encode for an RdRp near the C-terminus, and some may encode for a NP near the 5′ terminus. Some also encode for hypothetical proteins with unknown functions. | Auricularimonavirus: Auricularia heimuer negative-stranded RNA virus 1 Botrytimonavirus: Botrytis cinerea negative-stranded RNA virus 5 Lentimonavirus: Lentinula edodes negative-strand RNA virus 1 Penicillimonavirus: Penicillium adametzioides negative-stranded RNA virus 1 Sclerotimonavirus: Botrytis cinerea negative-stranded RNA virus 3 |
| Families and Genera | Genome Size and Segmentation | Genome Organization | Exemplar Species |
|---|---|---|---|
| Family Metaviridae Mycovirus associated genera Metavirus | Non-segmented genomes around 3 to 15 kb in size | Genome consists of an intragenic region which may code for up to 2 genes (gag and pol), which is flanked by long terminal repeat sequences (LTR) The gag gene usually encodes for the CP or NC protein while the pol gene encodes a polyprotein with PR, RT, INT and RH domains | Cladosporium fulvum T-1 virus |
| Family Pseudoviridae Mycovirus associated genera Hemivirus Pseudovirus | Non-segmented genomes that range in length from 4 to 9 kb in size | Genome consists of an internal region with 1 (gag-pol) or 2 ORFs (gag and pol), which are flanked by long terminal repeat sequences (LTR) The gag protein typically contains the CP and NC domains, while the pol protein contains PR, RT, INT, and RH domains | Hemivirus: Candida albicans Tca2 virus Pseudovirus: Saccharomyces cerevisiae Ty1 virus |
| Families and Genera | Genome Size and Segmentation | Genome Organization | Exemplar Species |
|---|---|---|---|
| Family Genomoviridae Mycovirus associated genera Gemycircularvirus Gemytripvirus | Gemycircularvirus Monopartite genomes around 1.8 to 2.4 kb in size Gemytripvirus Tripartite genome with individual segments around 1.3 kb | Gemycircularvirus Genome encodes for a rolling circle REP protein as well as a CP in ambisense orientation Gemytripvirus DNA-A: Encodes for the REP protein DNA-B: Encodes for the CP DNA-C: Encodes for p26, a protein with an unknown function | Gemycircularvirus Sclerotinia sclerotiorum hypovirulence-associated DNA virus 1 Gemytripvirus Fusarium graminearum gemytripvirus 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hough, B.; Steenkamp, E.; Wingfield, B.; Read, D. Fungal Viruses Unveiled: A Comprehensive Review of Mycoviruses. Viruses 2023, 15, 1202. https://doi.org/10.3390/v15051202
Hough B, Steenkamp E, Wingfield B, Read D. Fungal Viruses Unveiled: A Comprehensive Review of Mycoviruses. Viruses. 2023; 15(5):1202. https://doi.org/10.3390/v15051202
Chicago/Turabian StyleHough, Bianca, Emma Steenkamp, Brenda Wingfield, and David Read. 2023. "Fungal Viruses Unveiled: A Comprehensive Review of Mycoviruses" Viruses 15, no. 5: 1202. https://doi.org/10.3390/v15051202