


Molecular Characterization of Canine Parvovirus Variants CPV-2a and CPV-2c, Associated with Vaccinated Dogs at Libreville, Gabon
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Clinical Findings and Dog Characteristics
2.3. DNA Extraction and PCR Amplification
2.4. High-Throughput Sequencing
2.5. Phylogenetic Analysis
3. Results
3.1. Molecular Diagnosis and Sequences Analysis
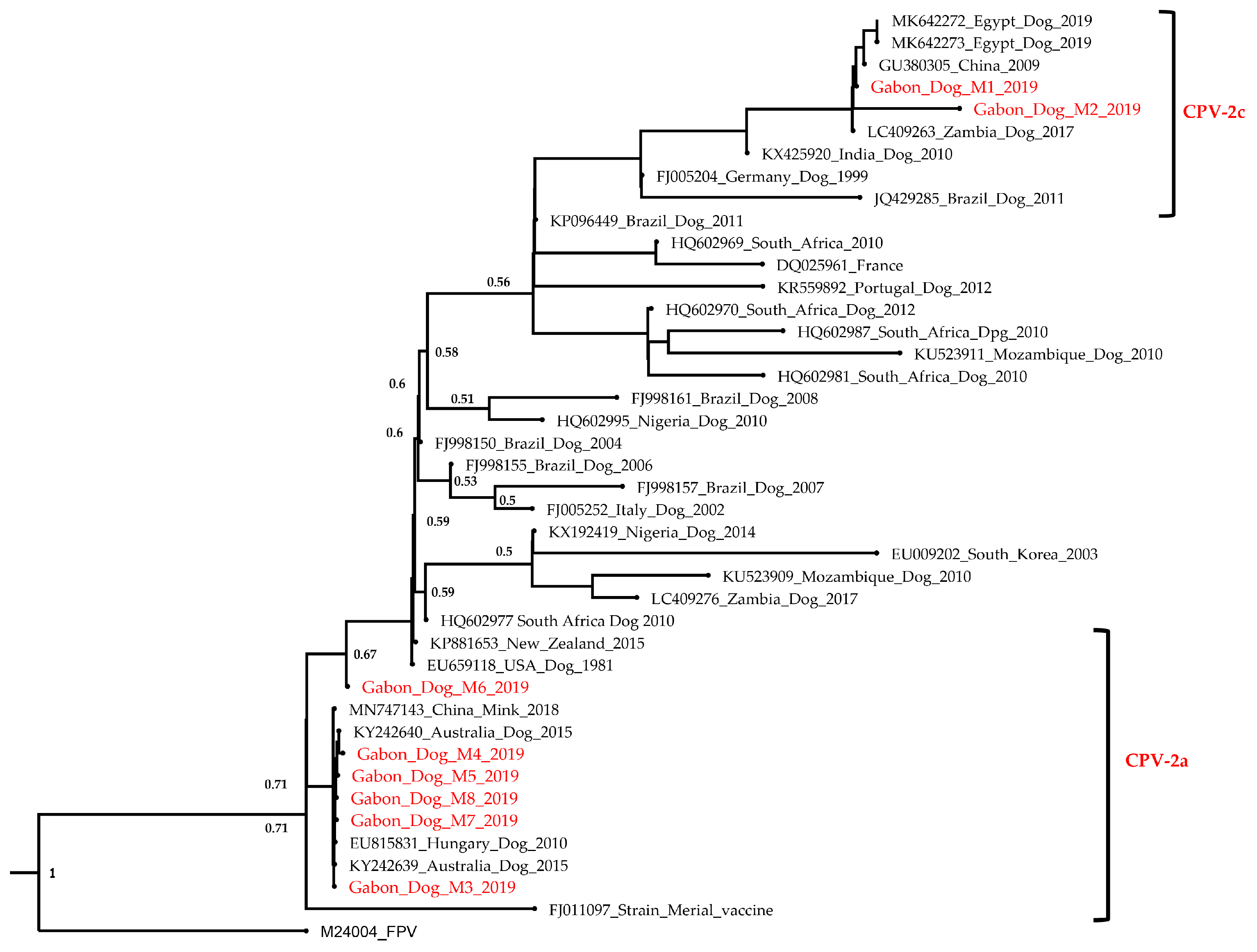
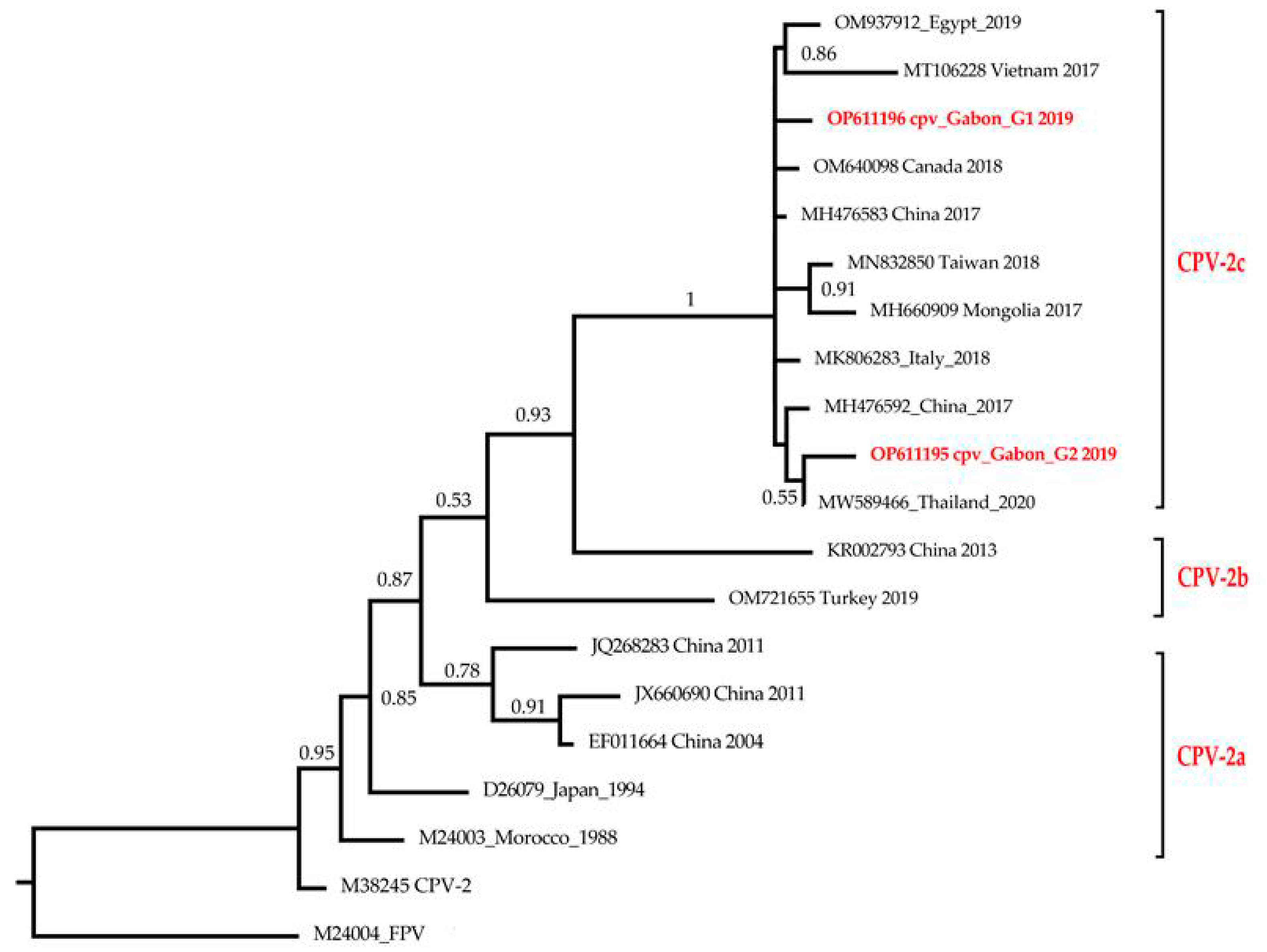
3.2. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.-M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Pénzes, J.J. ICTV virus taxonomy profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367. [Google Scholar] [CrossRef] [PubMed]
- Parrish, C.R. Host range relationships and the evolution of canine parvovirus. Vet. Microbiol. 1999, 69, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Touihri, L.; Bouzid, I.; Daoud, R.; Desario, C.; El Goulli, A.F.; Decaro, N.; Ghorbel, A.; Buonavoglia, C.; Bahloul, C. Molecular characterization of canine parvovirus-2 variants circulating in Tunisia. Virus Genes 2009, 38, 249–258. [Google Scholar] [CrossRef]
- Chollom, S.; Fyaktu, E.; Okwori, A.; Agada, G.; Hashimu, G.; Akele, R.; Voumangai, E.; Dashe, T.; Egah, D. Molecular detection of canine parvovirus in Jos, Nigeria. J. Vet. Med. Anim. Health 2013, 5, 57–59. [Google Scholar]
- Houston, D.; Ribble, C.; Head, L. Risk factors associated with parvovirus enteritis in dogs: 283 cases (1982–1991). J. Am. Vet. Med. Assoc. 1996, 208, 542–546. [Google Scholar]
- Appel, M.; Scott, F.; Carmichael, L. Isolation and immunisation studies of a canine parco-like virus from dogs with haemorrhagic enteritis. Vet. Rec. 1979, 105, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Desario, C.; Billi, M.; Lorusso, E.; Colaianni, M.; Colao, V.; Elia, G.; Ventrella, G.; Kusi, I.; Bo, S. Evaluation of an in-clinic assay for the diagnosis of canine parvovirus. Vet. J. 2013, 198, 504–507. [Google Scholar] [CrossRef]
- Johnson, R.; Spradbrow, P. Isolation from dogs with severe enteritis of a parvovirus related to feline panleucopaenia virus. Aust. Vet. J. 1979, 55, 151–152. [Google Scholar] [CrossRef]
- Decaro, N.; Buonavoglia, C. Canine parvovirus—A review of epidemiological and diagnostic aspects, with emphasis on type 2c. Vet. Microbiol. 2012, 155, 1–12. [Google Scholar] [CrossRef]
- Decaro, N.; Campolo, M.; Desario, C.; Elia, G.; Martella, V.; Lorusso, E.; Buonavoglia, C. Maternally-derived antibodies in pups and protection from canine parvovirus infection. Biologicals 2005, 33, 261–267. [Google Scholar] [CrossRef]
- Brady, S.; Norris, J.M.; Kelman, M.; Ward, M.P. Canine parvovirus in Australia: The role of socio-economic factors in disease clusters. Vet. J. 2012, 193, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Desario, C.; Amorisco, F.; Losurdo, M.; Elia, G.; Parisi, A.; Ventrella, G.; Martella, V.; Buonavoglia, C. Detection of a canine parvovirus type 2c with a non-coding mutation and its implications for molecular characterisation. Vet. J. 2013, 196, 555–557. [Google Scholar] [CrossRef]
- Decaro, N.; Elia, G.; Campolo, M.; Desario, C.; Lucente, M.; Bellacicco, A.; Buonavoglia, C. New approaches for the molecular characterization of canine parvovirus type 2 strains. J. Vet. Med. Ser. B 2005, 52, 316–319. [Google Scholar] [CrossRef] [PubMed]
- Buonavoglia, C.; Martella, V.; Pratelli, A.; Tempesta, M.; Cavalli, A.; Buonavoglia, D.; Bozzo, G.; Elia, G.; Decaro, N.; Carmichael, L. Evidence for evolution of canine parvovirus type 2 in Italy. J. Gen. Virol. 2001, 82, 3021–3025. [Google Scholar] [CrossRef]
- de Oliveira, P.S.B.; Cargnelutti, J.F.; Masuda, E.K.; Weiblen, R.; Flores, E.F. New variants of canine parvovirus in dogs in southern Brazil. Arch. Virol. 2019, 164, 1361–1369. [Google Scholar] [CrossRef]
- Castillo, C.; Neira, V.; Aniñir, P.; Grecco, S.; Pérez, R.; Panzera, Y.; Zegpi, N.-A.; Sandoval, A.; Sandoval, D.; Cofre, S. First molecular identification of canine parvovirus type 2 (CPV2) in Chile reveals high occurrence of CPV2c antigenic variant. Front. Vet. Sci. 2020, 7, 194. [Google Scholar] [CrossRef]
- Miranda, C.; Thompson, G. Canine parvovirus: The worldwide occurrence of antigenic variants. J. Gen. Virol. 2016, 97, 2043–2057. [Google Scholar] [CrossRef]
- Wilson, S.; Illambas, J.; Siedek, E.; Stirling, C.; Thomas, A.; Plevová, E.; Sture, G.; Salt, J. Vaccination of dogs with canine parvovirus type 2b (CPV-2b) induces neutralising antibody responses to CPV-2a and CPV-2c. Vaccine 2014, 32, 5420–5424. [Google Scholar] [CrossRef]
- Day, M.J.; Horzinek, M.; Schultz, R.; Squires, R. WSAVA Guidelines for the vaccination of dogs and cats. J. Small Anim. Pract. 2016, 57, E1. [Google Scholar] [CrossRef] [PubMed]
- Kapiya, J.; Nalubamba, K.S.; Kaimoyo, E.; Changula, K.; Chidumayo, N.; Saasa, N.; Simuunza, M.C.; Takada, A.; Mweene, A.S.; Chitanga, S. First genetic detection and characterization of canine parvovirus from diarrheic dogs in Zambia. Arch. Virol. 2019, 164, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Zaher, K.S.; El-Dabae, W.H.; El-Sebelgy, M.M.; Aly, N.I.; Salama, Z.T. Genotyping and phylogenetic analysis of canine parvovirus circulating in Egypt. Vet. World 2020, 13, 326. [Google Scholar] [CrossRef] [PubMed]
- Altman, K.; Kelman, M.; Ward, M. Are vaccine strain, type or administration protocol risk factors for canine parvovirus vaccine failure? Vet. Microbiol. 2017, 210, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Buonavoglia, C.; Barrs, V. Canine parvovirus vaccination and immunisation failures: Are we far from disease eradication? Vet. Microbiol. 2020, 247, 108760. [Google Scholar] [CrossRef] [PubMed]
- Amrani, N.; Desario, C.; Kadiri, A.; Cavalli, A.; Berrada, J.; Zro, K.; Sebbar, G.; Colaianni, M.L.; Parisi, A.; Elia, G. Molecular epidemiology of canine parvovirus in Morocco. Infect. Genet. Evol. 2016, 41, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Fagbohun, O.; Omobowale, T. Sequence and phylogenetic analysis of canine parvovirus-2 isolates in dogs revealed circulation of three subtypes in Nigeria. Virusdisease 2018, 29, 411–415. [Google Scholar] [CrossRef]
- Steinel, A.; Parrish, C.; Truyen, U.; Van Vuuren, M.; Venter, E.H. Antigenic and genetic analysis of canine parvoviruses in southern Africa. Onderstepoort J. Vet. Res. 1998, 65, 239–242. [Google Scholar]
- Dogonyaro, B.B.; Bosman, A.-M.; Sibeko, K.P.; Venter, E.H.; van Vuuren, M. Genetic analysis of the VP2-encoding gene of canine parvovirus strains from Africa. Vet. Microbiol. 2013, 165, 460–465. [Google Scholar] [CrossRef]
- Figueiredo, J.; Miranda, C.; Souto, R.; Silva, E.; Fafetine, J.; Thompson, G. Genetic characterization of canine parvovirus type 2 subtypes in Maputo, Mozambique. Arch. Microbiol. 2017, 199, 543–549. [Google Scholar] [CrossRef]
- Ngwa, V.N.; Bayanga, H.G.E.; Kouamo, J. Epidemiology of Canine parvovirus Enteritis in Dogs of the Metropolitan City of Yaounde, Cameroon. J. Anim. Vet. Adv. 2020, 19, 129–136. [Google Scholar]
- Chollom, S.; Ularamu, H.; Woma, T.; Owolodun, O.; Luka, P.; Anvou, A.; Nwankiti, O.; Egah, D. Detection of canine parvovirus 2: A comparison of conventional polymerase chain reaction and haemagglutination test. Afr. J. Microbiol. Res. 2012, 6, 4592–4596. [Google Scholar] [CrossRef]
- Antipov, D.; Raiko, M.; Lapidus, A.; Pevzner, P.A. Metaviral SPAdes: Assembly of viruses from metagenomic data. Bioinformatics 2020, 36, 4126–4129. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Decaro, N.; Elia, G.; Buonavoglia, C. Surveillance activity for canine parvovirus in Italy. J. Vet. Med. Ser. B 2005, 52, 312–315. [Google Scholar] [CrossRef]
- Unemo, M.; Golparian, D.; Skogen, V.; Olsen, A.O.; Moi, H.; Syversen, G.; Hjelmevoll, S.O. Neisseria gonorrhoeae strain with high-level resistance to spectinomycin due to a novel resistance mechanism (mutated ribosomal protein S5) verified in Norway. Antimicrob. Agents Chemother. 2013, 57, 1057–1061. [Google Scholar] [CrossRef]
- Lin, C.-N.; Chiang, S.-Y. Canine Parvovirus Type 2. In Canine Medicine—Recent Topics and Advanced Research; Kaoud, H.A.E., Ed.; Tech Publications: Rijeka, Croatia, 2016; pp. 1–17. [Google Scholar]
- Rabbani, A.H.; Ullah, Q.; Naseer, O.; Raza, A.I.; Shahid, M.; Ali, S.; Hussain, K.; Ali, A.; Khan, Y.R. Canine Parvo Virus: A Review on Current Perspectives in Seroprevalence, Diagnostics and Therapeutics. Glob. Vet. 2021, 23, 113–126. [Google Scholar]
- Prittie, J. Canine parvoviral enteritis: A review of diagnosis, management, and prevention. J. Vet. Emerg. Crit. Care 2004, 14, 167–176. [Google Scholar] [CrossRef]
- Otto, C.M.; Drobatz, K.J.; Soter, C. Endotoxemia and tumor necrosis factor activity in dogs with naturally occurring parvoviral enteritis. J. Vet. Intern. Med. 1997, 11, 65–70. [Google Scholar] [CrossRef]
- Turk, J.; Miller, M.; Brown, T.; Fales, W.; Fischer, J.; Gosser, H.; Nelson, S.; Shaw, D.; Solorzano, R. Coliform septicemia and pulmonary disease associated with canine parvoviral enteritis: 88 cases (1987-1988). J. Am. Vet. Med. Assoc. 1990, 196, 771–773. [Google Scholar]
- Umar, S.; Ali, A.; Younus, M.; Maan, M.K.; Shahzad, A.; Khan, W.A.; Irfan, M. Prevalence of canine parvovirus infection at different pet clinics in Lahore, Pakistan. Pak. J. Zool. 2015, 47, 657–663. [Google Scholar]
- Franzo, G.; De Villiers, L.; De Villiers, M.; Ravandi, A.; Gyani, K.; Van Zyl, L.; Coetzee, L.M.; Khaiseb, S.; Molini, U. Molecular epidemiology of canine parvovirus in Namibia: Introduction pathways and local persistence. Prev. Vet. Med. 2022, 209, 105780. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, D.; Banu, K.; Mondal, B. Canine parvovirology—A brief updated review on structural biology, occurrence, pathogenesis, clinical diagnosis, treatment and prevention. Comp. Immunol. Microbiol. Infect. Dis. 2022, 82, 101765. [Google Scholar] [CrossRef] [PubMed]
- Maya, L.; Calleros, L.; Francia, L.; Hernández, M.; Iraola, G.; Panzera, Y.; Sosa, K.; Pérez, R. Phylodynamics analysis of canine parvovirus in Uruguay: Evidence of two successive invasions by different variants. Arch. Virol. 2013, 158, 1133–1141. [Google Scholar] [CrossRef]
- Ohneiser, S.; Hills, S.; Cave, N.; Passmore, D.; Dunowska, M. Canine parvoviruses in New Zealand form a monophyletic group distinct from the viruses circulating in other parts of the world. Vet. Microbiol. 2015, 178, 190–200. [Google Scholar] [CrossRef]
- Bull, J. Evolutionary reversion of live viral vaccines: Can genetic engineering subdue it? Virus Evol. 2015, 1, vev005. [Google Scholar] [CrossRef]
- Packianathan, R.; Hodge, A.; Wright, J.; Lavidis, L.; Ameiss, K.; Yip, H.Y.E.; Akbarzadeh, M.; Sharifian, M.; Amanollahi, R.; Khabiri, A. Cross-Neutralization of Vanguard C4 Vaccine Against Australian Isolates of Canine Parvovirus Variants CPV-2a, CPV-2b, and CPV-2c. Viral Immunol. 2022, 35, 553–558. [Google Scholar] [CrossRef]
- Calderon, M.G.; Mattion, N.; Bucafusco, D.; Fogel, F.; Remorini, P.; La Torre, J. Molecular characterization of canine parvovirus strains in Argentina: Detection of the pathogenic variant CPV2c in vaccinated dogs. J. Virol. Methods 2009, 159, 141–145. [Google Scholar] [CrossRef]
- Larson, L.J.; Schultz, R. Do two current canine parvovirus type 2 and 2b vaccines provide protection against the new type 2c variant? Vet. Ther. 2008, 9, 94. [Google Scholar]
- Carmichael, L. An annotated historical account of canine parvovirus. J. Vet. Med. Ser. B 2005, 52, 303–311. [Google Scholar] [CrossRef]
- Goddard, A.; Leisewitz, A.L. Canine parvovirus. Vet. Clin. Small Anim. Pract. 2010, 40, 1041–1053. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A. Small Animal Pathology for Veterinary Technicians; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Pérez, R.; Calleros, L.; Marandino, A.; Sarute, N.; Iraola, G.; Grecco, S.; Blanc, H.; Vignuzzi, M.; Isakov, O.; Shomron, N. Phylogenetic and genome-wide deep-sequencing analyses of canine parvovirus reveal co-infection with field variants and emergence of a recent recombinant strain. PLoS ONE 2014, 9, e111779. [Google Scholar] [CrossRef] [PubMed]
- Battilani, M.; Gallina, L.; Vaccari, F.; Morganti, L. Co-infection with multiple variants of canine parvovirus type 2 (CPV-2). Vet. Res. Commun. 2007, 31, 209. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.J.; Silva, E.; Desario, C.; Decaro, N.; Carvalheira, J.; Buonavoglia, C.; Thompson, G. Natural coinfection with 2 parvovirus variants in dog. Emerg. Infect. Dis. 2008, 14, 678. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| N | Age (Months) | Sex | Clinical Signs |
|---|---|---|---|
| 1 | 6 | F | Diarrhea; weakness; fever; vomiting |
| 2 | 5 | M | Bloody diarrhea; weakness; fever |
| 3 | 8 | F | Diarrhea; loss of appetite; fever; vomiting |
| 4 | 6 | M | Bloody diarrhea; weakness |
| 5 | 8 | F | Diarrhea; weakness; vomiting |
| 6 | 12 | F | Diarrhea; weakness |
| 7 | 6 | F | Diarrhea; weakness; vomiting |
| 8 | 5 | F | Bloody diarrhea, weakness, vomiting |
| Accession Numbers | 80 | 87 | 93 | 101 | 103 | 297 | 300 | 305 | 323 | 426 | 555 | 564 | 568 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OP611197 | - | - | - | - | - | - | - | - | - | E | V | - | - |
| OP611198 | - | - | - | - | - | - | - | - | - | E | V | - | - |
| OP611199 | - | - | - | - | - | - | - | - | - | N | V | - | - |
| OP611200 | - | - | - | - | - | - | - | - | - | N | V | - | - |
| OP611201 | - | - | - | - | - | - | - | - | - | N | V | - | - |
| OP611202 | - | - | - | - | - | - | - | - | - | N | V | - | - |
| OP611203 | - | - | - | - | - | - | - | - | - | N | V | - | - |
| OP611204 | - | - | - | - | - | - | - | - | - | N | V | - | - |
| CPV-2 | R | M | N | I | V | S | A | D | N | N | V | S | G |
| CPV-2a | R | L | N | T | A | S | G | Y | N | N | I | S | G |
| CPV-2b | R | L | N | T | A | S | G | Y | N | D | V | S | G |
| CPV-2c | R | L | N | T | A | A | G | Y | N | E | V | S | G |
| FPV | K | M | K | I | V | S | A | D | D | N | V | N | A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maganga, G.D.; Labouba, I.; Milendz Ikapi, S.Z.; Nkili-Meyong, A.A.; Ngonga Dikongo, A.M.; Boundenga, L.; Ngoubangoye, B.; Memvie, C.; Kumulungui, B.S. Molecular Characterization of Canine Parvovirus Variants CPV-2a and CPV-2c, Associated with Vaccinated Dogs at Libreville, Gabon. Viruses 2023, 15, 1169. https://doi.org/10.3390/v15051169
Maganga GD, Labouba I, Milendz Ikapi SZ, Nkili-Meyong AA, Ngonga Dikongo AM, Boundenga L, Ngoubangoye B, Memvie C, Kumulungui BS. Molecular Characterization of Canine Parvovirus Variants CPV-2a and CPV-2c, Associated with Vaccinated Dogs at Libreville, Gabon. Viruses. 2023; 15(5):1169. https://doi.org/10.3390/v15051169
Chicago/Turabian StyleMaganga, Gael Darren, Ingrid Labouba, Serda Zita Milendz Ikapi, Andriniaina Andy Nkili-Meyong, Audrey Michel Ngonga Dikongo, Larson Boundenga, Barthelemy Ngoubangoye, Christelle Memvie, and Brice Serge Kumulungui. 2023. "Molecular Characterization of Canine Parvovirus Variants CPV-2a and CPV-2c, Associated with Vaccinated Dogs at Libreville, Gabon" Viruses 15, no. 5: 1169. https://doi.org/10.3390/v15051169