
Clustered Cases of Waterborne Hepatitis E Virus Infection, France

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Descriptive Epidemiology
2.2. Clinical Laboratory Investigations
2.3. Environmental Investigations
2.4. Ethical Approval
3. Results
3.1. Descriptive Epidemiology
3.2. Clinical Laboratory Investigations
3.3. Environmental Investigations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kupferschmidt, K. Europe’s new hepatitis problem. Science 2016, 353, 862–863. [Google Scholar] [CrossRef] [PubMed]
- Lhomme, S.; Marion, O.; Abravanel, F.; Izopet, J.; Kamar, N. Clinical Manifestations, Pathogenesis and Treatment of Hepatitis E Virus Infections. J. Clin. Med. 2020, 9, 331. [Google Scholar] [CrossRef] [PubMed]
- Purdy, M.A.; Drexler, J.F.; Meng, X.J.; Norder, H.; Okamoto, H.; Van der Poel, W.H.M.; Reuter, G.; de Souza, W.M.; Ulrich, R.G.; Smith, D.B. ICTV Virus Taxonomy Profile: Hepeviridae 2022. J. Gen. Virol. 2022, 103, 001778. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.E.; Labrique, A.B.; Kmush, B.L. Epidemiology of Genotype 1 and 2 Hepatitis E Virus Infections. Cold Spring Harb. Perspect. Med. 2019, 9, a031732. [Google Scholar] [CrossRef]
- Dalton, H.R.; Izopet, J. Transmission and Epidemiology of Hepatitis E Virus Genotype 3 and 4 Infections. Cold Spring Harb. Perspect. Med. 2018, 8, a032144. [Google Scholar] [CrossRef]
- Renou, C.; Roque-Afonso, A.M.; Pavio, N. Foodborne transmission of hepatitis E virus from raw pork liver sausage, France. Emerg. Infect. Dis. 2014, 20, 1945–1947. [Google Scholar] [CrossRef]
- Guillois, Y.; Abravanel, F.; Miura, T.; Pavio, N.; Vaillant, V.; Lhomme, S.; Le Guyader, F.S.; Rose, N.; Le Saux, J.C.; King, L.A.; et al. High Proportion of Asymptomatic Infections in an Outbreak of Hepatitis E Associated with a Spit-Roasted Piglet, France, 2013. Clin. Infect. Dis. 2016, 62, 351–357. [Google Scholar] [CrossRef]
- Tei, S.; Kitajima, N.; Takahashi, K.; Mishiro, S. Zoonotic transmission of hepatitis E virus from deer to human beings. Lancet 2003, 362, 371–373. [Google Scholar] [CrossRef]
- Takuissu, G.R.; Kenmoe, S.; Ndip, L.; Ebogo-Belobo, J.T.; Kengne-Nde, C.; Mbaga, D.S.; Bowo-Ngandji, A.; Oyono, M.G.; Kenfack-Momo, R.; Tchatchouang, S.; et al. Hepatitis E Virus in Water Environments: A Systematic Review and Meta-analysis. Food Environ. Virol. 2022, 14, 223–235. [Google Scholar] [CrossRef]
- Fenaux, H.; Chassaing, M.; Berger, S.; Gantzer, C.; Bertrand, I.; Schvoerer, E. Transmission of hepatitis E virus by water: An issue still pending in industrialized countries. Water Res. 2019, 151, 144–157. [Google Scholar] [CrossRef]
- Mansuy, J.M.; Gallian, P.; Dimeglio, C.; Saune, K.; Arnaud, C.; Pelletier, B.; Morel, P.; Legrand, D.; Tiberghien, P.; Izopet, J. A nationwide survey of hepatitis E viral infection in French blood donors. Hepatology 2016, 63, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Izopet, J.; Pavio, N.; Aggarwal, R.; Labrique, A.; Wedemeyer, H.; Dalton, H.R. Hepatitis E virus infection. Nat. Rev. Dis. Prim. 2017, 3, 17086. [Google Scholar] [CrossRef] [PubMed]
- Lhomme, S.; Legrand-Abravanel, F.; Kamar, N.; Izopet, J. Screening, diagnosis and risks associated with Hepatitis E virus infection. Expert. Rev. Anti-Infect. Ther. 2019, 17, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Mansuy, J.M.; Bendall, R.; Legrand-Abravanel, F.; Saune, K.; Miedouge, M.; Ellis, V.; Rech, H.; Destruel, F.; Kamar, N.; Dalton, H.R.; et al. Hepatitis E virus antibodies in blood donors, France. Emerg. Infect. Dis. 2011, 17, 2309–2312. [Google Scholar] [CrossRef]
- Abravanel, F.; Sandres-Saune, K.; Lhomme, S.; Dubois, M.; Mansuy, J.M.; Izopet, J. Genotype 3 diversity and quantification of hepatitis E virus RNA. J. Clin. Microbiol. 2012, 50, 897–902. [Google Scholar] [CrossRef]
- Legrand-Abravanel, F.; Mansuy, J.M.; Dubois, M.; Kamar, N.; Peron, J.M.; Rostaing, L.; Izopet, J. Hepatitis E virus genotype 3 diversity, France. Emerg. Infect. Dis. 2009, 15, 110–114. [Google Scholar] [CrossRef]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G.; et al. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef]
- Nicot, F.; Jeanne, N.; Roulet, A.; Lefebvre, C.; Carcenac, R.; Manno, M.; Dubois, M.; Kamar, N.; Lhomme, S.; Abravanel, F.; et al. Diversity of hepatitis E virus genotype 3. Rev. Med. Virol. 2018, 28, e1987. [Google Scholar] [CrossRef]
- Martin-Latil, S.; Hennechart-Collette, C.; Guillier, L.; Perelle, S. Duplex RT-qPCR for the detection of hepatitis E virus in water, using a process control. Int. J. Food Microbiol. 2012, 157, 167–173. [Google Scholar] [CrossRef]
- Hazam, R.K.; Singla, R.; Kishore, J.; Singh, S.; Gupta, R.K.; Kar, P. Surveillance of hepatitis E virus in sewage and drinking water in a resettlement colony of Delhi: What has been the experience? Arch. Virol. 2010, 155, 1227–1233. [Google Scholar] [CrossRef]
- Naik, S.R.; Aggarwal, R.; Salunke, P.N.; Mehrotra, N.N. A large waterborne viral hepatitis E epidemic in Kanpur, India. Bull. World Health Organ. 1992, 70, 597–604. [Google Scholar] [PubMed]
- Crossan, C.; Baker, P.J.; Craft, J.; Takeuchi, Y.; Dalton, H.R.; Scobie, L. Hepatitis E virus genotype 3 in shellfish, United Kingdom. Emerg. Infect. Dis. 2012, 18, 2085–2087. [Google Scholar] [CrossRef] [PubMed]
- Said, B.; Ijaz, S.; Kafatos, G.; Booth, L.; Thomas, H.L.; Walsh, A.; Ramsay, M.; Morgan, D.; Hepatitis, E.I.I.T. Hepatitis E outbreak on cruise ship. Emerg. Infect. Dis. 2009, 15, 1738–1744. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.M.; Costafreda, M.I.; Bosch, A. Risk assessment in shellfish-borne outbreaks of hepatitis A. Appl. Environ. Microbiol. 2009, 75, 7350–7355. [Google Scholar] [CrossRef] [PubMed]
- Potasman, I.; Paz, A.; Odeh, M. Infectious outbreaks associated with bivalve shellfish consumption: A worldwide perspective. Clin. Infect. Dis. 2002, 35, 921–928. [Google Scholar] [CrossRef]
- Le Guyader, F.; Haugarreau, L.; Miossec, L.; Dubois, E.; Pommepuy, M. Three-year study to assess human enteric viruses in shellfish. Appl. Environ. Microbiol. 2000, 66, 3241–3248. [Google Scholar] [CrossRef]
- Rutjes, S.A.; Lodder, W.J.; Lodder-Verschoor, F.; van den Berg, H.H.; Vennema, H.; Duizer, E.; Koopmans, M.; de Roda Husman, A.M. Sources of hepatitis E virus genotype 3 in The Netherlands. Emerg. Infect. Dis. 2009, 15, 381–387. [Google Scholar] [CrossRef]
- Steyer, A.; Naglic, T.; Mocilnik, T.; Poljsak-Prijatelj, M.; Poljak, M. Hepatitis E virus in domestic pigs and surface waters in Slovenia: Prevalence and molecular characterization of a novel genotype 3 lineage. Infect. Genet. Evol. 2011, 11, 1732–1737. [Google Scholar] [CrossRef]
- La Rosa, G.; Pourshaban, M.; Iaconelli, M.; Vennarucci, V.S.; Muscillo, M. Molecular detection of hepatitis E virus in sewage samples. Appl. Environ. Microbiol. 2010, 76, 5870–5873. [Google Scholar] [CrossRef]
- Miura, T.; Lhomme, S.; Le Saux, J.C.; Le Mehaute, P.; Guillois, Y.; Couturier, E.; Izopet, J.; Abranavel, F.; Le Guyader, F.S. Detection of Hepatitis E Virus in Sewage After an Outbreak on a French Island. Food Environ. Virol. 2016, 8, 194–199. [Google Scholar] [CrossRef]
- Tsarev, S.A.; Tsareva, T.S.; Emerson, S.U.; Yarbough, P.O.; Legters, L.J.; Moskal, T.; Purcell, R.H. Infectivity titration of a prototype strain of hepatitis E virus in cynomolgus monkeys. J. Med. Virol. 1994, 43, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Kjellberg, I.; Sikora, P.; Rydberg, H.; Lindh, M.; Bergstedt, O.; Norder, H. Hepatitis E virus genotype 3 strains and a plethora of other viruses detected in raw and still in tap water. Water Res. 2020, 168, 115141. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, S.J. Detection of infectious enteroviruses and adenoviruses in tap water in urban areas in Korea. Water Res. 2002, 36, 248–256. [Google Scholar] [CrossRef]
- Salvador, D.; Neto, C.; Benoliel, M.J.; Caeiro, M.F. Assessment of the Presence of Hepatitis E virus in Surface Water and Drinking Water in Portugal. Microorganisms 2020, 8, 761. [Google Scholar] [CrossRef]
- Kumar, T.; Shrivastava, A.; Bhatia, D.; Mitra, Y.; Kumar, A.; Hussain, S.; Chauhan, L.S.; Laserson, K.F.; Narain, J.P.; Kumar, R.; et al. Jaundice outbreak likely caused by HEV in Amritsar, Punjab, India, 2013. BMC Public Health 2019, 19, 464. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, A.S.; Sharma, M.; Thorat, N.C.; Jadhav, S.; Koshatwar, K.A. An outbreak of hepatitis E in Yavatmal, India, 2019. J. Med. Virol. 2021, 93, 3761–3768. [Google Scholar] [CrossRef]
- Lenglet, A.; Ehlkes, L.; Taylor, D.; Fesselet, J.F.; Nassariman, J.N.; Ahamat, A.; Chen, A.; Noh, I.; Moustapha, A.; Spina, A. Does community-wide water chlorination reduce hepatitis E virus infections during an outbreak? A geospatial analysis of data from an outbreak in Am Timan, Chad (2016–2017). J. Water Health 2020, 18, 556–565. [Google Scholar] [CrossRef]
- Berto, A.; Van der Poel, W.H.; Hakze-van der Honing, R.; Martelli, F.; La Ragione, R.M.; Inglese, N.; Collins, J.; Grierson, S.; Johne, R.; Reetz, J.; et al. Replication of hepatitis E virus in three-dimensional cell culture. J. Virol. Methods 2013, 187, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Fenaux, H.; Chassaing, M.; Berger, S.; Jeulin, H.; Gentilhomme, A.; Bensenane, M.; Bronowicki, J.P.; Gantzer, C.; Bertrand, I.; Schvoerer, E. Molecular features of Hepatitis E Virus circulation in environmental and human samples. J. Clin. Virol. 2018, 103, 63–70. [Google Scholar] [CrossRef]
- Kaba, M.; Davoust, B.; Marie, J.L.; Colson, P. Detection of hepatitis E virus in wild boar (Sus scrofa) livers. Vet. J. 2010, 186, 259–261. [Google Scholar] [CrossRef]
- Gentry-Shields, J.; Myers, K.; Pisanic, N.; Heaney, C.; Stewart, J. Hepatitis E virus and coliphages in waters proximal to swine concentrated animal feeding operations. Sci. Total Environ. 2015, 505, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, M.P.; Scott, R.M.; Joshi, D.M.; Mammen, M.P., Jr.; Thapa, G.B.; Thapa, N.; Myint, K.S.; Fourneau, M.; Kuschner, R.A.; Shrestha, S.K.; et al. Safety and efficacy of a recombinant hepatitis E vaccine. N. Engl. J. Med. 2007, 356, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shih, J.W.; Xia, N.S. Long-term efficacy of a hepatitis E vaccine. N. Engl. J. Med. 2015, 372, 2265–2266. [Google Scholar] [CrossRef] [PubMed]



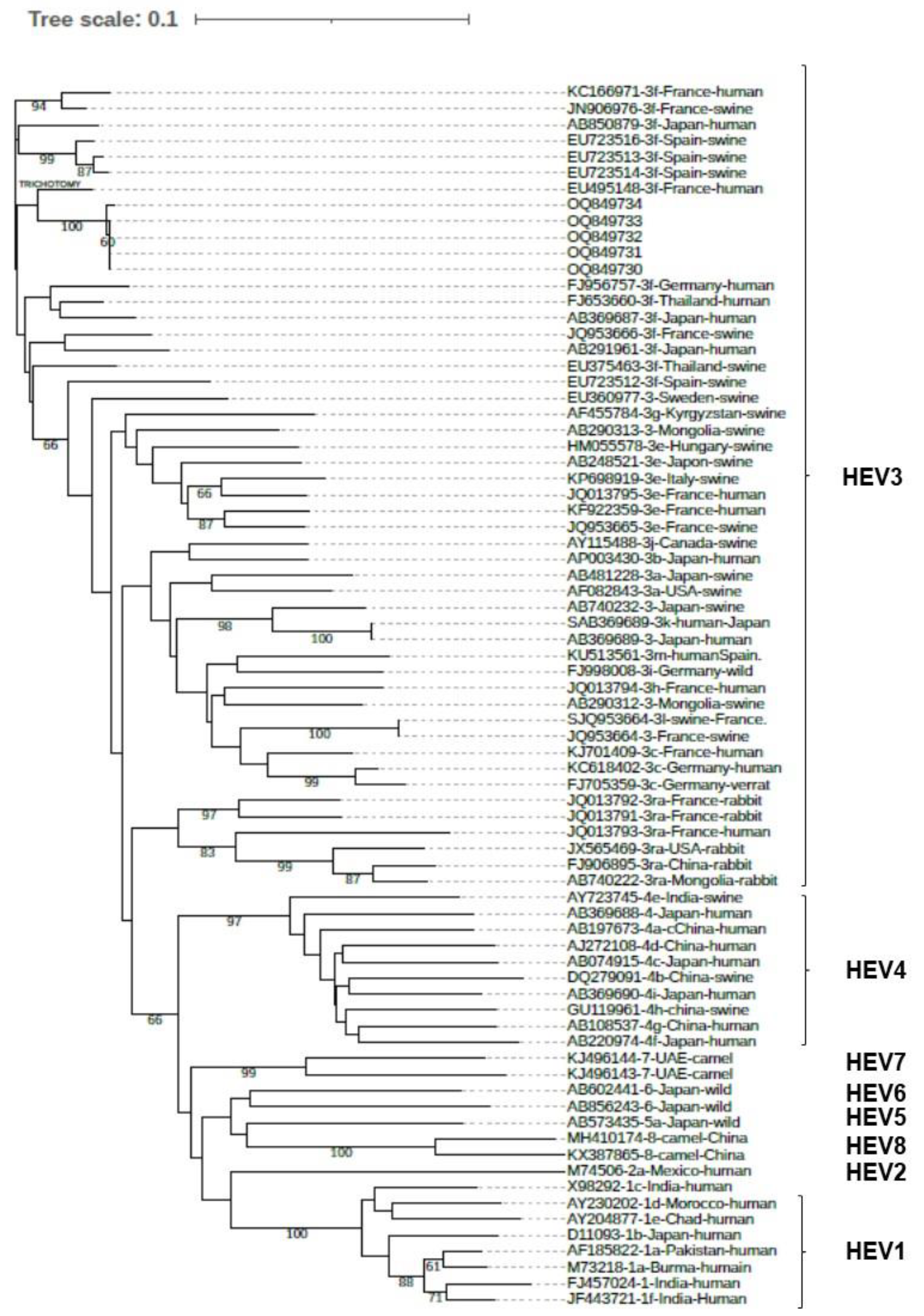{kind=link}
{kind=link}
| Case | Age (Years) | Sex | House Number Where the Patients Live | Sampling Date (Year-Week) | Comorbidities | Clinical Signs | Disease Outcome | Estimated Exposure Period (Week) |
|---|---|---|---|---|---|---|---|---|
| Case 1 | 78 | Male | 1 | 2015-10 | Diabetes, heart failure, chronic alcoholism | Jaundice, vomiting, diarrhea, alteration of the general state | Death | 2–8 |
| Case 2 | 46 | Female | 2 | 2015-13 | - | Asthenia | Self-limiting | 5–11 |
| Case 3 | 63 | Female | 3 | 2015-14 | - | Asthenia | Self-limiting | 7–8 |
| Case 4 | 60 | Male | 3 | 2015-13 | - | None | - | 5–11 |
| Case 5 | 27 | Male | 3 | 2015-13 | - | Asthenia | Self-limiting | 5–11 |
| Case 6 | 58 | Male | 4 | 2015-14 | - | None | - | 6–12 |
| Case 7 | 62 | Male | 4 | 2015-14 | - | None | - | 6–12 |
| Risk Factor | Number of Patients |
|---|---|
| Travel abroad | 0/6 |
| Contact with: | |
| Domestic animals | 4/6 |
| Farm animals | 3/6 |
| Boar or deer | 0/6 |
| Consumption of pork products: | |
| Cured ham | 6/6 |
| Bacon | 6/6 |
| Dry-cured sausage | 6/6 |
| Garlic sausage | 5/6 |
| Sausage | 5/6 |
| Pork roast | 5/6 |
| Boiled ham | 4/6 |
| Liver pâté | 4/6 |
| Black pudding | 1/6 |
| Raw pork liver products | 0/6 |
| Consumption of shellfish | 2/6 |
| Consumption of water from public network | 6/6 |
| Case | Detection of Anti-HEV IgM | Detection of HEV RNA | HEV RNA Concentration (IU/mL) | Genotype/Subgenotype |
|---|---|---|---|---|
| 1 | not tested | positive | 96,750 | 3f |
| 2 | positive | positive | <75 | 3f |
| 3 | positive | positive | 325 | - |
| 4 | positive | negative | - | - |
| 5 | positive | positive | <75 | 3f |
| 6 | positive | positive | <75 | 3f |
| 7 | positive | positive | <75 | 3f |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lhomme, S.; Magne, S.; Perelle, S.; Vaissière, E.; Abravanel, F.; Trelon, L.; Hennechart-Collette, C.; Fraisse, A.; Martin-Latil, S.; Izopet, J.; et al. Clustered Cases of Waterborne Hepatitis E Virus Infection, France. Viruses 2023, 15, 1149. https://doi.org/10.3390/v15051149
Lhomme S, Magne S, Perelle S, Vaissière E, Abravanel F, Trelon L, Hennechart-Collette C, Fraisse A, Martin-Latil S, Izopet J, et al. Clustered Cases of Waterborne Hepatitis E Virus Infection, France. Viruses. 2023; 15(5):1149. https://doi.org/10.3390/v15051149
Chicago/Turabian StyleLhomme, Sébastien, Sébastien Magne, Sylvie Perelle, Emmanuelle Vaissière, Florence Abravanel, Laetitia Trelon, Catherine Hennechart-Collette, Audrey Fraisse, Sandra Martin-Latil, Jacques Izopet, and et al. 2023. "Clustered Cases of Waterborne Hepatitis E Virus Infection, France" Viruses 15, no. 5: 1149. https://doi.org/10.3390/v15051149