
Genetic Analysis of Dengue Virus in Severe and Non-Severe Cases in Dhaka, Bangladesh, in 2018–2022

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Patients and Clinical Specimens
2.3. Detection of Dengue NS1 Antigen
2.4. RNA Extraction and Real-Time Reverse Transcriptase PCR
2.5. Serotype Specific Real-Time Reverse Transcriptase PCR
2.6. Sequence Determination of the DENV Envelope Region
2.7. Phylogenetic Analysis of Envelope Region
2.8. Mean Values of the Number of Nucleotide Differences among DENV3 E Regions
2.9. Statistical Analysis
3. Results
3.1. Dengue Demographic Data
3.2. Population Demography
3.3. Severity and Serotype
3.4. Clinical Features and Complications of Hospitalized Severe Dengue Cases
3.5. Laboratory Findings of Hospitalized Severe Dengue Cases
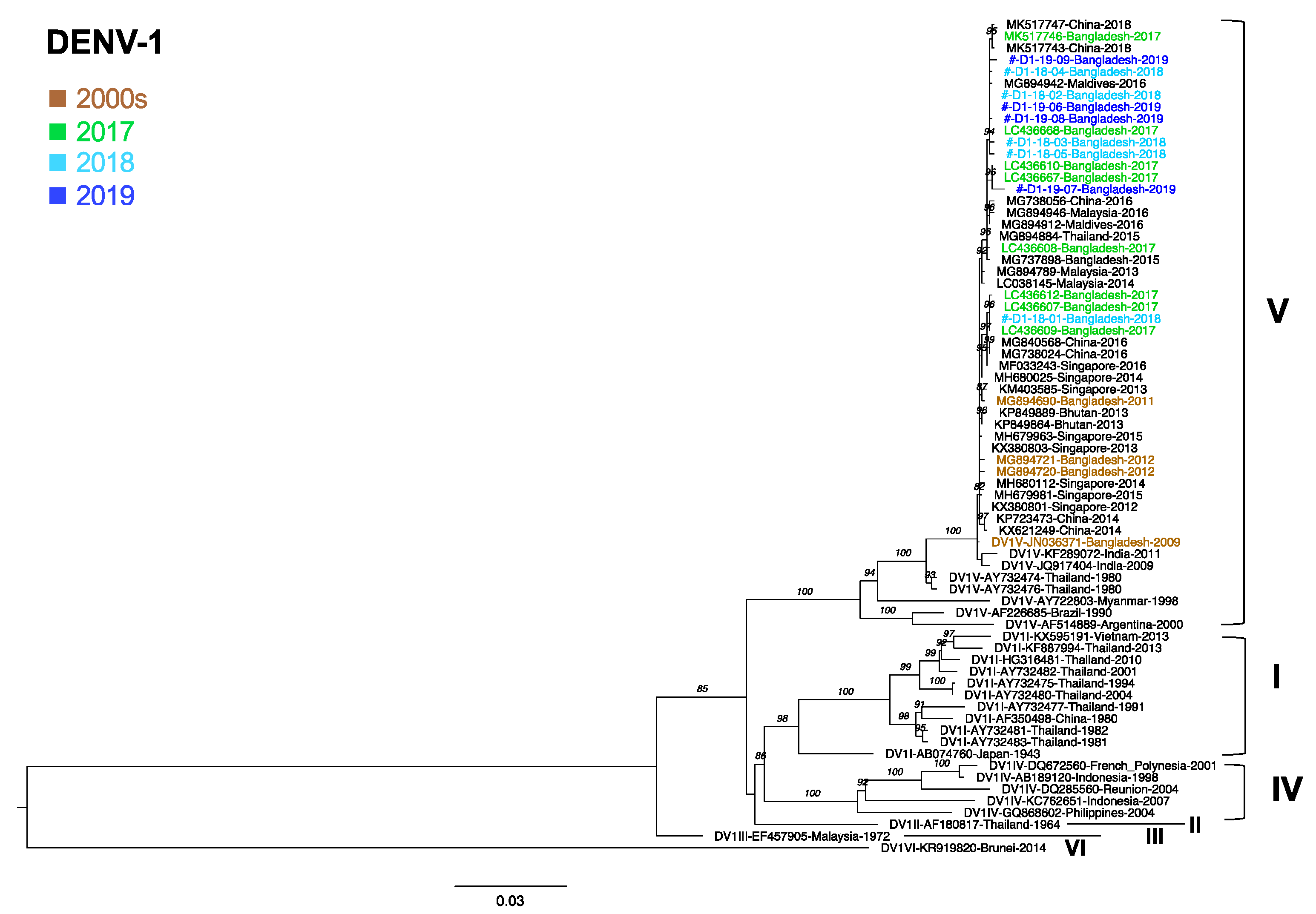
3.6. Phylogenetic Analysis of DENV1
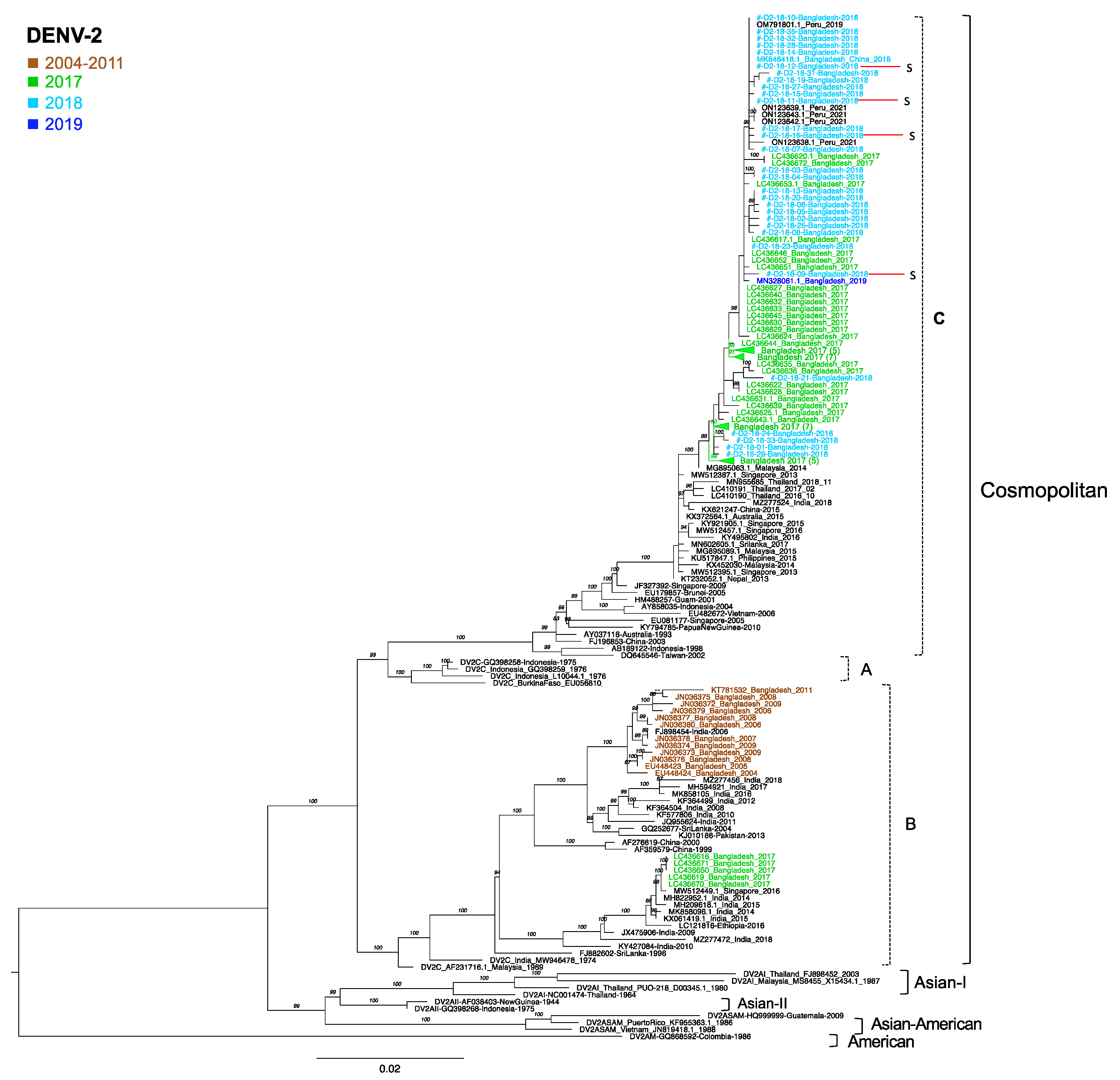
3.7. Phylogenetic Analysis of DENV2
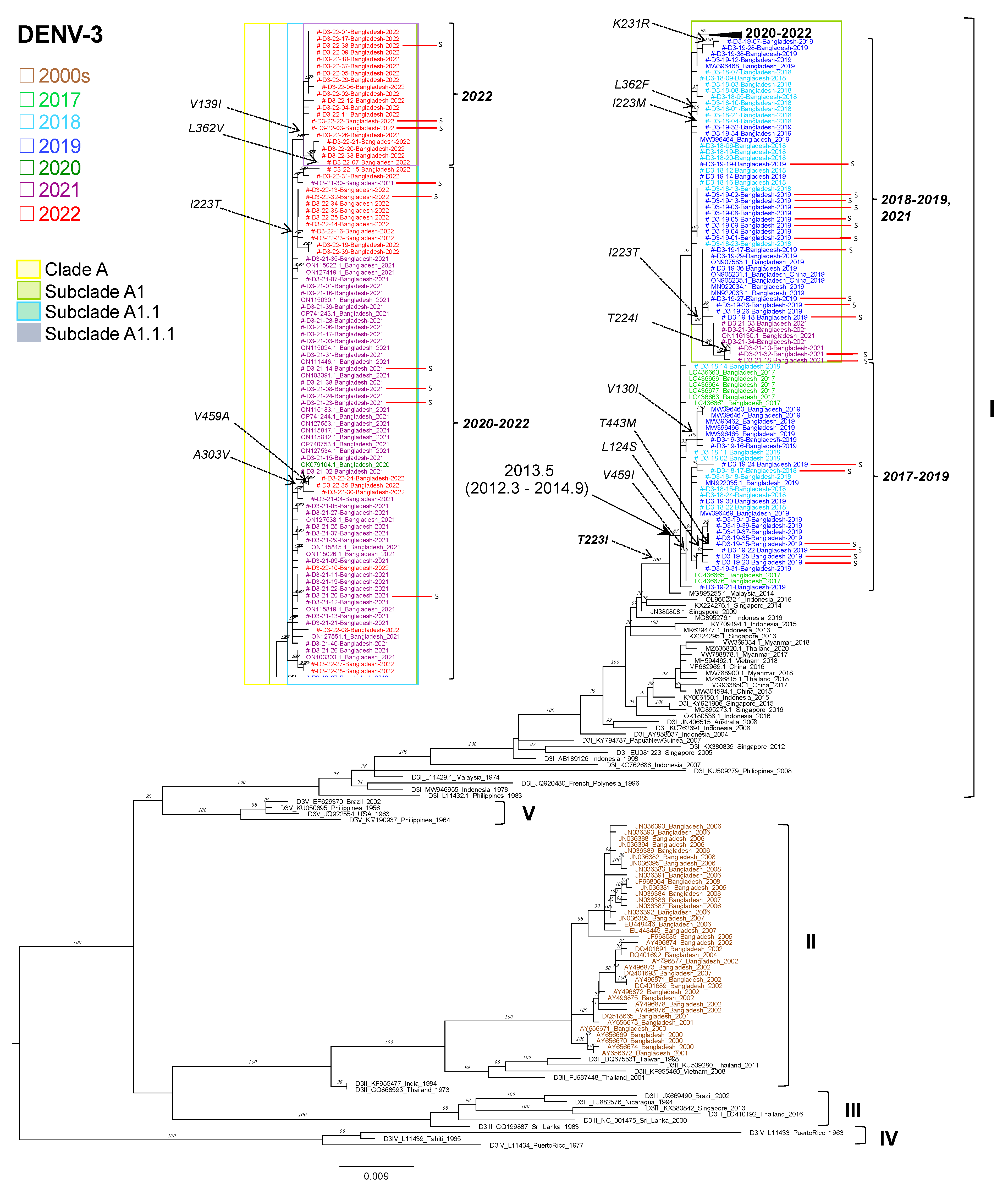
3.8. Phylogenetic Analysis of DENV3
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Regional Office for South-East; Dengue Bulletin; World Health Organization: Geneva, Switzerland, 2020; Volume 41. [Google Scholar]
- Lindenbach, B.D.; Rice, C.M. Flaviviridae: The viruses and their replication. In Fields Virology, 4th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2001; pp. 991–1041. [Google Scholar]
- Henchal, E.A.; Putnak, J.R. The dengue viruses. Clin. Microbiol. Rev. 1990, 3, 376–396. [Google Scholar] [CrossRef] [PubMed]
- Rico-Hesse, R. Molecular evolution and distribution of dengue viruses type 1 and 2 in nature. Virology 1990, 174, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Twiddy, S.S.; Holmes, E.; Rambaut, A. Inferring the Rate and Time-Scale of Dengue Virus Evolution. Mol. Biol. Evol. 2003, 20, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Goncalvez, A.P.; Escalante, A.; Pujol, F.H.; Ludert, J.E.; Tovar, D.; Salas, R.A.; Liprandi, F. Diversity and Evolution of the Envelope Gene of Dengue Virus Type 1. Virology 2002, 303, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Pyke, A.T.; Moore, P.R.; Taylor, C.T.; Hall-Mendelin, S.; Cameron, J.N.; Hewitson, G.R.; Pukallus, D.S.; Huang, B.; Warrilow, D.; van den Hurk, A.F. Highly divergent dengue virus type 1 genotype sets a new distance record. Sci. Rep. 2016, 6, 22356. [Google Scholar] [CrossRef]
- Anez, G.; Morales-Betoulle, M.E.; Rios, M. Circulation of different lineages of dengue virus type 2 in Central America, their evolutionary time-scale and selection pressure analysis. PLoS ONE 2011, 6, e27459. [Google Scholar] [CrossRef]
- Khan, M.A.; Ellis, E.M.; Tissera, H.A.; Alvi, M.Y.; Rahman, F.F.; Masud, F.; Chow, A.; Howe, S.; Dhanasekaran, V.; Ellis, B.R.; et al. Emergence and Diversification of Dengue 2 Cosmopolitan Genotype in Pakistan, 2011. PLoS ONE 2013, 8, e56391. [Google Scholar] [CrossRef]
- Twiddy, S.S.; Farrar, J.J.; Vinh Chau, N.; Wills, B.; Gould, E.A.; Gritsun, T.; Lloyd, G.; Holmes, E.C. Phylogenetic relationships and differential selection pressures among genotypes of dengue-2 virus. Virology 2002, 298, 63–72. [Google Scholar] [CrossRef]
- Wittke, V.; Robb, T.; Thu, H.; Nisalak, A.; Nimmannitya, S.; Kalayanrooj, S.; Vaughn, D.; Endy, T.; Holmes, E.; Aaskov, J. Extinction and Rapid Emergence of Strains of Dengue 3 Virus during an Interepidemic Period. Virology 2002, 301, 148–156. [Google Scholar] [CrossRef]
- AbuBakar, S.; Wong, P.-F.; Chan, Y.F. Emergence of dengue virus type 4 genotype IIA in Malaysia. J. Gen. Virol. 2002, 83, 2437–2442. [Google Scholar] [CrossRef] [PubMed]
- Klungthong, C.; Zhang, C.; Mammen, M.P., Jr.; Ubol, S.; Holmes, E.C. The molecular epidemiology of dengue virus serotype 4 in Bangkok, Thailand. Virology 2004, 329, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S.; Gubler, D.J.; Trent, D.W. Molecular evolution and phylogeny of dengue-4 viruses. J. Gen. Virol. 1997, 78 Pt 9, 2279–2284. [Google Scholar] [CrossRef] [PubMed]
- Gentry, M.K.; McCown, J.M.; Dalrymple, J.M.; Brandt, W.E.; Henchal, E.A. Identification of Distinct Antigenic Determinants on Dengue-2 Virus Using Monoclonal Antibodies. Am. J. Trop. Med. Hyg. 1982, 31, 548–555. [Google Scholar] [CrossRef]
- Russell, P.K.; Nisalak, A. Dengue Virus Identification by the Plaque Reduction Neutralization Test. J. Immunol. 1967, 99, 291–296. [Google Scholar] [CrossRef]
- Bell, S.M.; Katzelnick, L.; Bedford, T. Dengue genetic divergence generates within-serotype antigenic variation, but serotypes dominate evolutionary dynamics. eLife 2019, 8, e42496. [Google Scholar] [CrossRef]
- Russell, P.K.; Buescher, E.L.; McCown, J.M.; Ordoñez, J. Recovery of Dengue Viruses from Patients during Epidemics in Puerto Rico and East Pakistan. Am. J. Trop. Med. Hyg. 1966, 15, 573–579. [Google Scholar] [CrossRef]
- Aziz, M.; Hasan, K.N.; Hasanat, M.A.; Siddiqui, M.A.; Salimullah, M.; Chowdhury, A.K.; Ahmed, M.; Alam, M.N.; Hassan, M.S. Predominance of the DEN-3 genotype during the recent dengue outbreak in Bangladesh. Southeast Asian J. Trop. Med. Public Health 2002, 33, 42–48. [Google Scholar]
- Muraduzzaman, A.K.M.; Alam, A.N.; Sultana, S.; Siddiqua, M.; Khan, M.H.; Akram, A.; Haque, F.; Flora, M.S.; Shirin, T. Circulating dengue virus serotypes in Bangladesh from 2013 to 2016. Virus Dis. 2018, 29, 303–307. [Google Scholar] [CrossRef]
- Racherla, R.G.; Pamireddy, M.L.; Mohan, A.; Mudhigeti, N.; Mahalakshmi, P.A.; Nallapireddy, U.; Kalawat, U. Co-Circulation of Four Dengue Serotypes at South Eastern Andhra Pradesh, India: A Prospective Study. Indian J. Med. Microbiol. 2018, 36, 236–240. [Google Scholar] [CrossRef]
- Nunes, P.C.G.; Sampaio, S.A.F.; da Costa, N.R.; de Mendonça, M.C.L.; Lima, M.D.R.Q.; Araujo, S.E.M.; dos Santos, F.B.; Simões, J.B.S.; Gonçalves, B.D.S.; Nogueira, R.M.R.; et al. Dengue severity associated with age and a new lineage of dengue virus-type 2 during an outbreak in Rio De Janeiro, Brazil. J. Med. Virol. 2016, 88, 1130–1136. [Google Scholar] [CrossRef] [PubMed]
- Hang, V.T.T.; Holmes, E.; Veasna, D.; Quy, N.T.; Hien, T.T.; Quail, M.; Churcher, C.; Parkhill, J.; Cardosa, J.; Farrar, J.; et al. Emergence of the Asian 1 Genotype of Dengue Virus Serotype 2 in Viet Nam: In Vivo Fitness Advantage and Lineage Replacement in South-East Asia. PLoS Negl. Trop. Dis. 2010, 4, e757. [Google Scholar] [CrossRef] [PubMed]
- Rahim, R.; Hasan, A.; Hasan, N.; Nakayama, E.E.; Shioda, T.; Rahman, M. Diversity of Dengue Virus Serotypes in Dhaka City: From 2017 to 2021. Bangladesh J. Med. Microbiol. 2021, 15, 23–29. [Google Scholar] [CrossRef]
- Chen, R.; Vasilakis, N. Dengue—Quo tu et quo vadis? Viruses 2011, 3, 1562–1608. [Google Scholar] [CrossRef]
- Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 1 September 2022).
- Available online: http://ncbi.nlm.nih.gov/genbank (accessed on 1 September 2022).
- Trifinopoulos, J.; Nguyen, L.-T.; Von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- National Institutes of Health. Available online: https://www.nih.gov/nih-style-guide/age (accessed on 1 November 2022).
- Poltep, K.; Phadungsombat, J.; Nakayama, E.E.; Kosoltanapiwat, N.; Hanboonkunupakarn, B.; Wiriyarat, W.; Shioda, T.; Leaungwutiwong, P. Genetic Diversity of Dengue Virus in Clinical Specimens from Bangkok, Thailand, during 2018–2020: Co-Circulation of All Four Serotypes with Multiple Genotypes and/or Clades. Trop. Med. Infect. Dis. 2021, 6, 162. [Google Scholar] [CrossRef]
- Phadungsombat, J.; Lin, M.Y.-C.; Srimark, N.; Yamanaka, A.; Nakayama, E.E.; Moolasart, V.; Suttha, P.; Shioda, T.; Uttayamakul, S. Emergence of genotype Cosmopolitan of dengue virus type 2 and genotype III of dengue virus type 3 in Thailand. PLoS ONE 2018, 13, e0207220. [Google Scholar] [CrossRef]
- Suzuki, K.; Nakayama, E.E.; Saito, A.; Egawa, A.; Sato, T.; Phadungsombat, J.; Rahim, R.; Abu Hasan, A.; Iwamoto, H.; Rahman, M.; et al. Evaluation of novel rapid detection kits for dengue virus NS1 antigen in Dhaka, Bangladesh, in 2017. Virol. J. 2019, 16, 102. [Google Scholar] [CrossRef]
- Moore, P.R.; van den Hurk, A.F.; Mackenzie, J.S.; Pyke, A.T. Dengue viruses in Papua New Guinea: Evidence of endemicity and phylogenetic variation, including the evolution of new genetic lineages. Emerg. Microbes Infect. 2017, 6, e114. [Google Scholar] [CrossRef] [PubMed]
- Shu, P.Y.; Su, C.L.; Liao, T.L.; Yang, C.F.; Chang, S.F.; Lin, C.C.; Chang, M.C.; Hu, H.C.; Huang, J.H. Molecular characterization of dengue viruses imported into Taiwan during 2003-2007: Geographic distribution and genotype shift. Am. J. Trop. Med. Hyg. 2009, 80, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- OhAinle, M.; Balmaseda, A.; Macalalad, A.R.; Tellez, Y.; Zody, M.C.; Saborío, S.; Nuñez, A.; Lennon, N.J.; Birren, B.W.; Gordon, A.; et al. Dynamics of Dengue Disease Severity Determined by the Interplay between Viral Genetics and Serotype-Specific Immunity. Sci. Transl. Med. 2011, 3, 114ra128. [Google Scholar] [CrossRef] [PubMed]
- Yung, C.-F.; Lee, K.-S.; Thein, T.-L.; Tan, L.-K.; Gan, V.; Wong, J.; Lye, D.; Ng, L.-C.; Leo, Y.-S. Dengue Serotype-Specific Differences in Clinical Manifestation, Laboratory Parameters and Risk of Severe Disease in Adults, Singapore. Am. J. Trop. Med. Hyg. 2015, 92, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Higgs, S. Impact of climate change and other factors on emerging arbovirus diseases. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 109–121. [Google Scholar] [CrossRef]
- Swamy, A.M.; Mahesh, P.Y.; Tumkur, S.R. Liver function in dengue and its correlation with disease severity: An retrospective cross-sectional observational study in a tertiary care center in Coastal India. Pan Afr. Med. J. 2021, 40, 261. [Google Scholar] [CrossRef]
- Shirin, T.; Muraduzzaman, A.; Alam, A.; Sultana, S.; Siddiqua, M.; Khan, M.; Akram, A.; Sharif, A.; Hossain, S.; Flora, M. Largest dengue outbreak of the decade with high fatality may be due to reemergence of DEN-3 serotype in Dhaka, Bangladesh, necessitating immediate public health attention. New Microbes New Infect. 2019, 29, 100511. [Google Scholar] [CrossRef]
- Riad, M.H.; Cohnstaedt, L.W.; Scoglio, C.M. Risk Assessment of Dengue Transmission in Bangladesh Using a Spatiotemporal Network Model and Climate Data. Am. J. Trop. Med. Hyg. 2021, 104, 1444–1455. [Google Scholar] [CrossRef]
- Titir, S.R.; Paul, S.K.; Ahmed, S.; Haque, N.; Nasreen, S.A.; Hossain, K.S.; Ahmad, F.U.; Nila, S.S.; Khanam, J.; Nowsher, N.; et al. Nationwide Distribution of Dengue Virus Type 3 (DENV3) Genotype I and Emergence of DENV3 Genotype III during the 2019 Outbreak in Bangladesh. Trop. Med. Infect. Dis. 2021, 6, 58. [Google Scholar]
- Dengue Serotype-3 Dominates in Dhaka; The Business Standard Report; Business Standard Ltd.: Dhaka, Bangladesh, 2021.
- Kalita, J.M.; Aggarwal, A.; Yedale, K.; Gadepalli, R.; Nag, V.L. A 5-year study of dengue seropositivity among suspected cases attending a teaching hospital of North-Western region of India. J. Med. Virol. 2021, 93, 3338–3343. [Google Scholar] [CrossRef]
- Suzuki, K.; Phadungsombat, J.; Nakayama, E.E.; Saito, A.; Egawa, A.; Sato, T.; Rahim, R.; Hasan, A.; Lin, M.Y.-C.; Takasaki, T.; et al. Genotype replacement of dengue virus type 3 and clade replacement of dengue virus type 2 genotype Cosmopolitan in Dhaka, Bangladesh in 2017. Infect. Genet. Evol. 2019, 75, 103977. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.F.; Chang, S.F.; Hsu, T.C.; Su, C.L.; Wang, T.C.; Lin, S.H.; Yang, S.L.; Lin, C.C.; Shu, P.Y. Molecular characterization and phylogenetic analysis of dengue viruses imported into Taiwan during 2011–2016. PLoS Negl. Trop. Dis. 2018, 12, e0006773. [Google Scholar] [CrossRef] [PubMed]
- Zangmo, S.; Darnal, J.B.; Tsheten; Gyeltshen, S.; Thapa, B.T.; Rodpradit, P.; Chinnawirotpisan, P.; Manasatienkij, W.; Macareo, L.R.; Fernandez, S.; et al. Molecular epidemiology of dengue fever outbreaks in Bhutan, 2016–2017. PLoS Negl. Trop. Dis. 2020, 14, e0008165. [Google Scholar] [CrossRef] [PubMed]
- Pandey, B.D.; Nabeshima, T.; Pandey, K.; Rajendra, S.P.; Shah, Y.; Adhikari, B.R.; Gupta, G.; Gautam, I.; Tun, M.M.N.; Uchida, R.; et al. First Isolation of Dengue Virus from the 2010 Epidemic in Nepal. Trop. Med. Health 2013, 41, 103–111. [Google Scholar] [CrossRef]
- De Bruycker-Nogueira, F.; Souza, T.M.A.; Chouin-Carneiro, T.; Faria, N.R.D.C.; Santos, J.B.; Torres, M.C.; Ramalho, I.L.C.; de Aguiar, S.F.; Nogueira, R.M.R.; de Filippis, A.M.B.; et al. DENV-1 Genotype V in Brazil: Spatiotemporal dispersion pattern reveals continuous co-circulation of distinct lineages until 2016. Sci. Rep. 2018, 8, 17160. [Google Scholar] [CrossRef]
- Dieng, I.; Cunha, M.d.P.; Diagne, M.M.; Sembène, P.M.; Zanotto, P.M.d.A.; Faye, O.; Faye, O.; Sall, A.A. Origin and Spread of the Dengue Virus Type 1, Genotype V in Senegal, 2015–2019. Viruses 2021, 13, 57. [Google Scholar] [CrossRef]
- Alagarasu, K.; Patil, J.; Kakade, M.; More, A.; Yogesh, B.; Newase, P.; Jadhav, S.; Parashar, D.; Kaur, H.; Gupta, N.; et al. Serotype and genotype diversity of dengue viruses circulating in India: A multi-centre retrospective study involving the Virus Research Diagnostic Laboratory Network in 2018. Int. J. Infect. Dis. 2021, 111, 242–252. [Google Scholar] [CrossRef]
- Cecilia, D.; Patil, J.; Kakade, M.; Walimbe, A.; Alagarasu, K.; Anukumar, B.; Abraham, A. Emergence of the Asian genotype of DENV-1 in South India. Virology 2017, 510, 40–45. [Google Scholar] [CrossRef]
- Soe, A.M.; Ngwe Tun, M.M.; Nabeshima, T.; Myat, T.W.; Htun, M.M.; Lin, H.; Hom, N.S.; Inoue, S.; Nwe, K.M.; Aye, L.P.P.; et al. Emergence of a Novel Dengue Virus 3 (DENV-3) Genotype-I Coincident with Increased DENV-3 Cases in Yangon, Myanmar between 2017 and 2019. Viruses 2021, 13, 1152. [Google Scholar] [CrossRef]
- Cao, J.; Deng, H.; Ye, L.; Ma, X.; Chen, S.; Sun, X.; Wu, X.; Yan, T.; Zhang, L.; Liu, L.; et al. Epidemiological and clinical characteristics of Dengue virus outbreaks in two regions of China, 2014–2015. PLoS ONE 2019, 14, e0213353. [Google Scholar] [CrossRef]
- Ujwal, S.; Sabeena, S.; Bhaskar, R.; D’Souza, G.; Santhosha, D.; Auti, A.; Kumar, R.; Ramachandran, S.; Hindol, M.; Aithal, A.; et al. Circulation of Asian-I and Cosmopolitan genotypes of Dengue-2 virus in northeast India, 2016–2017. J. Vector Borne Dis. 2019, 56, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Haryanto, S.; Hayati, R.F.; Yohan, B.; Sijabat, L.; Sihite, I.F.; Fahri, S.; Meutiawati, F.; Halim, J.A.N.; Halim, S.N.; Soebandrio, A.; et al. The molecular and clinical features of dengue during outbreak in Jambi, Indonesia in 2015. Pathog. Glob. Health 2016, 110, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Roche, C.; Cassar, O.; Laille, M.; Murgue, B. Dengue-3 virus genomic differences that correlate with in vitro phenotype on a human cell line but not with disease severity. Microbes Infect. 2007, 9, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Suppiah, J.; Ching, S.M.; Amin-Nordin, S.; Mat-Nor, L.A.; Ahmad-Najimudin, N.A.; Low, G.K.; Abdul-Wahid, M.Z.; Thayan, R.; Chee, H.Y. Clinical manifestations of dengue in relation to dengue serotype and genotype in Malaysia: A retrospective observational study. PLoS Negl. Trop. Dis. 2018, 12, e0006817. [Google Scholar] [CrossRef]
- Tan, K.-K.; Zulkifle, N.-I.; Abd-Jamil, J.; Sulaiman, S.; Yaacob, C.N.; Azizan, N.S.; Seri, N.A.A.C.M.; Samsudin, N.I.; Mahfodz, N.H.; AbuBakar, S. Disruption of predicted dengue virus type 3 major outbreak cycle coincided with switching of the dominant circulating virus genotype. Infect. Genet. Evol. 2017, 54, 271–275. [Google Scholar] [CrossRef]
- Tripathi, S.K.; Gupta, P.; Khare, V.; Chatterjee, A.; Kumar, R.; Khan, M.Y.; Dhole, T.N. Emergence of new lineage of Dengue virus 3 (genotype III) in Lucknow, India. Iran. J. Microbiol. 2013, 5, 68–75. [Google Scholar]
- Samanta, J.; Sharma, V. Dengue and its effects on liver. World J. Clin. Cases 2015, 3, 125–131. [Google Scholar] [CrossRef]
- Dhanwada, S.; Reddy, S.S. A study of various hepatic manifestations in dengue fever and their correlation with severity of dengue fever. Int. J. Contemp. Pediatr. 2020, 7, 527–531. [Google Scholar] [CrossRef]
- Tsang, T.K.; Ghebremariam, S.L.; Gresh, L.; Gordon, A.; Halloran, M.E.; Katzelnick, L.C.; Rojas, D.P.; Kuan, G.; Balmaseda, A.; Sugimoto, J.; et al. Effects of infection history on dengue virus infection and pathogenicity. Nat. Commun. 2019, 10, 1246. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Pathogenesis of Severe Dengue. Available online: https://www.cdc.gov-page48046 (accessed on 7 May 2023).




{kind=link}
{kind=link}
{kind=link}
| 2018 | 2019 | 2020 | 2021 | 2022 | ||||
| Total number of tests (n = 3759) | 1016 | 566 | 130 | 1171 | 876 | |||
| Positive NS1 antigen/positive RT-PCR (n = 839) | 174 | 132 | 30 | 315 | 188 | |||
| Positive serotyping (n = 495) | 127 | 86 | 1 | 178 | 103 | |||
| Serotype | DENV1 | DENV2 | DENV3 | DENV1 | DENV3 | DENV3 | DENV3 | DENV3 |
| Positive cases | 33 (26) | 52 (40.9) | 42 (33.1) | 7 (8.1) | 79 (91.9) | 1 (100) | 178 (100) | 103 (100) |
| % male | 8 (6.3) | 12 (9.4) | 13 (10.2) | 2 (40) | 31 (55.36) | 19 (65.52) | 11 (55) | |
| Mean age | 20 | 31 | 23 | 13 | 17 | 30 | 30 | |
| Children and adolescent | 16 (12.6) | 19 (15) | 16 (12.6) | 4 (4.7) | 52 (60.5) | 0 | 53 (29.8) | 40 (38.8) |
| Adult | 15 (11.8) | 32 (25.2) | 25 (19.7) | 3 (3.5) | 27 (31.4) | 1 (100) | 117 (65.7) | 59 (57.3) |
| Elderly | 2 (1.6) | 1 (0.8) | 1 (0.8) | 0 | 0 | 0 | 8 (4.5) | 4 (3.9) |
| Hospitalization | ||||||||
| Inpatients n (%) | 13 (10.2) | 29 (22.8) | 22 (17.3) | 5 (5.8) | 64 (74.4) | 0 | 34 (19.1) | 21 (20.4) |
| Admission day | 3 (1–5) | 4 (1–6) | 4 (1–7) | 3 (1–4) | 3 (1–9) | 3 (1–4) | 3 (1–4) | |
| Days febrile | 4 (2–6) | 5 (2–8) | 5 (2–8) | 4 (2–6) | 5 (2–8) | 5 (2–8) | 5 (2–8) | |
| Days in hospital | 5 (1–12) | 5 (1–8) | 5 (1–8) | 3 (1–5) | 5 (2–14) | 5 (1–11) | 5 (1–10) | |
| Classical (%) | 12 (92.3) | 23 (79.3) | 16 (72.7) | 5 (100) | 29 (45.3) | 22 (64.7) | 14 (66.7) | |
| Severe (%) | 1 (7.7) | 6 (20.7) | 6 (27.3) | 0 | 35 (54.7) | 12 (35.3) | 7 (33.3) | |
| Sequence Analysis (n = 179) | 5 | 30 | 24 | 4 | 37 | 40 | 39 | |
| Complications | Number of Patients, n = 67 (%) * |
| Bleeding | 10 (14.9) |
| Pleural effusion | 2 (29.9) |
| Breathlessness | 12 (17.9) |
| Ascites | 9 (13.4) |
| Oliguria | 4 (6) |
| Hepatomegaly | 4 (6) |
| Splenomegaly | 2 (3) |
| Seizures | 1 (1.5) |
| Multiple organ failure | 3 (4.5) |
| Mechanical ventilation | 3 (4.5) |
| Sepsis | 3 (4.5) |
| Symptoms | |
| Anorexia | 12 (17.9) |
| Nausea | 36 (53.7) |
| Vomiting | 36 (53.7) |
| Abdominal pain | 37 (55.2) |
| Body ache | 23 (34.3) |
| Headache | 16 (23.9) |
| Cough | 7 (10.4) |
| Diarrhea | 6 (9) |
| Rash | 9 (13.4) |
| Weakness | 8 (11.9) |
| Lab Parameters | No. of Patients, n = 67 (%) | Median Value | p Value * |
| Platelet count (/cumm) | |||
| ≤50,000 | 40 (59.7) | 27,000 | <0.001 |
| >50,000 | 27 (40.3) | 80,000 | |
| Hematocrit value (%) | |||
| Normal hematocrit (Male) | 22 (32.8) | 42 | <0.001 |
| Raised hematocrit (Male) | 15 (22.4) | 46.6 | |
| Normal hematocrit (Female) | 22 (32.8) | 35.45 | <0.001 |
| Raised hematocrit (Female) | 8 (11.9) | 43.25 | |
| Hemoglobin (g/dL) | |||
| Normal (Male) | 27 (40.3) | 15.5 | <0.001 |
| Below normal (Male) | 10 (14.9) | 12.26 | |
| Normal (Female) | 10 (14.9) | 14.75 | <0.001 |
| Below normal (Female) | 20 (29.9) | 11.4 | |
| Albumin (g/dL) | |||
| Hypoalbuminemia | 56 (83.6) | 2.9 | <0.001 |
| Normal | 11 (16.4) | 4 | |
| Leukocyte count (/cumm) | |||
| ≤4000 | 16 (23.9) | 2935 | <0.001 |
| >4000 | 51 (76.1) | 6840 | |
| Liver enzyme (IU/L) | |||
| Raised ALT, AST | 58 (86.6) | 109.5, 123.5 | <0.001 ** |
| Normal ALT, AST | 9 (13.4) | 34, 30 | <0.001 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahim, R.; Hasan, A.; Phadungsombat, J.; Hasan, N.; Ara, N.; Biswas, S.M.; Nakayama, E.E.; Rahman, M.; Shioda, T. Genetic Analysis of Dengue Virus in Severe and Non-Severe Cases in Dhaka, Bangladesh, in 2018–2022. Viruses 2023, 15, 1144. https://doi.org/10.3390/v15051144
Rahim R, Hasan A, Phadungsombat J, Hasan N, Ara N, Biswas SM, Nakayama EE, Rahman M, Shioda T. Genetic Analysis of Dengue Virus in Severe and Non-Severe Cases in Dhaka, Bangladesh, in 2018–2022. Viruses. 2023; 15(5):1144. https://doi.org/10.3390/v15051144
Chicago/Turabian StyleRahim, Rummana, Abu Hasan, Juthamas Phadungsombat, Nazmul Hasan, Nikhat Ara, Suma Mita Biswas, Emi E. Nakayama, Mizanur Rahman, and Tatsuo Shioda. 2023. "Genetic Analysis of Dengue Virus in Severe and Non-Severe Cases in Dhaka, Bangladesh, in 2018–2022" Viruses 15, no. 5: 1144. https://doi.org/10.3390/v15051144