
Inhibitor of CD147 Suppresses T Cell Activation and Recruitment in CVB3-Induced Acute Viral Myocarditis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Virus and CVB3 Infection
2.3. Treatment with AC-73
2.4. Viral Genome Load and CD147 mRNA Expression in the Heart
2.5. Histopathological Analysis
2.6. Fluorescence Staining
2.7. Immunohistochemistry
2.8. FACS Analysis for T Cell Subtype
2.9. Luminex Multiplex Immunoassay for Cytokine and Chemokine Profiling
2.10. Cytokine mRNA Expression
2.11. Statistical Analysis
3. Results
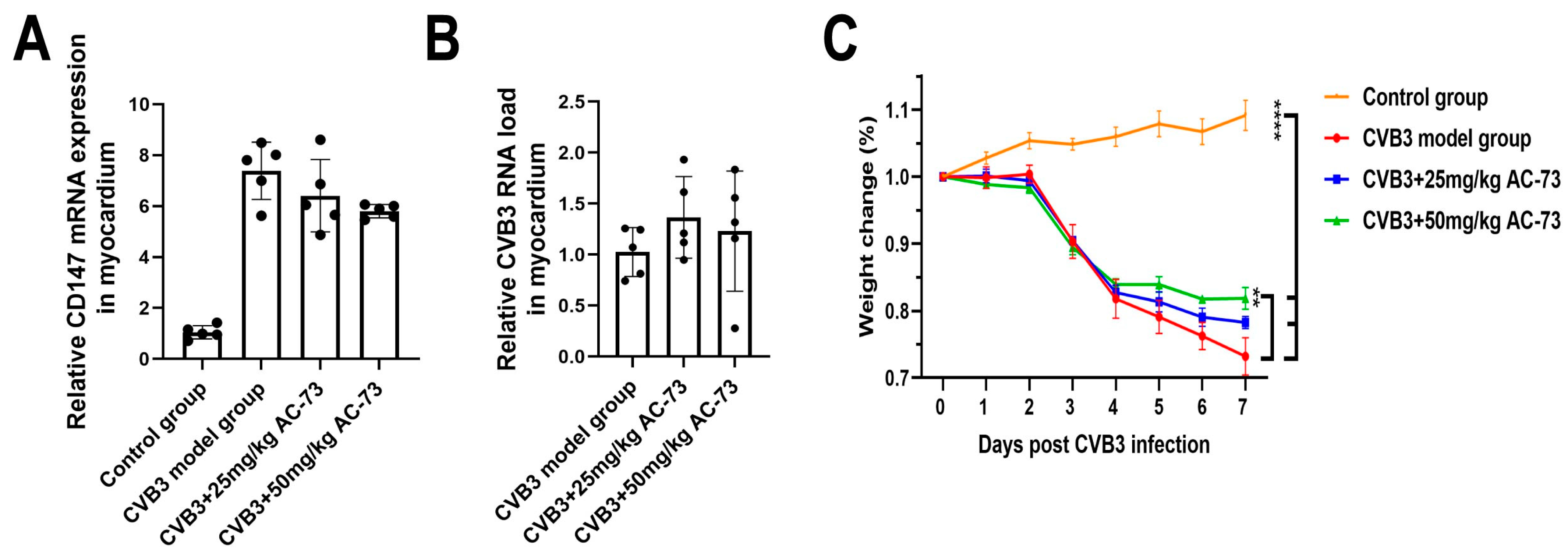
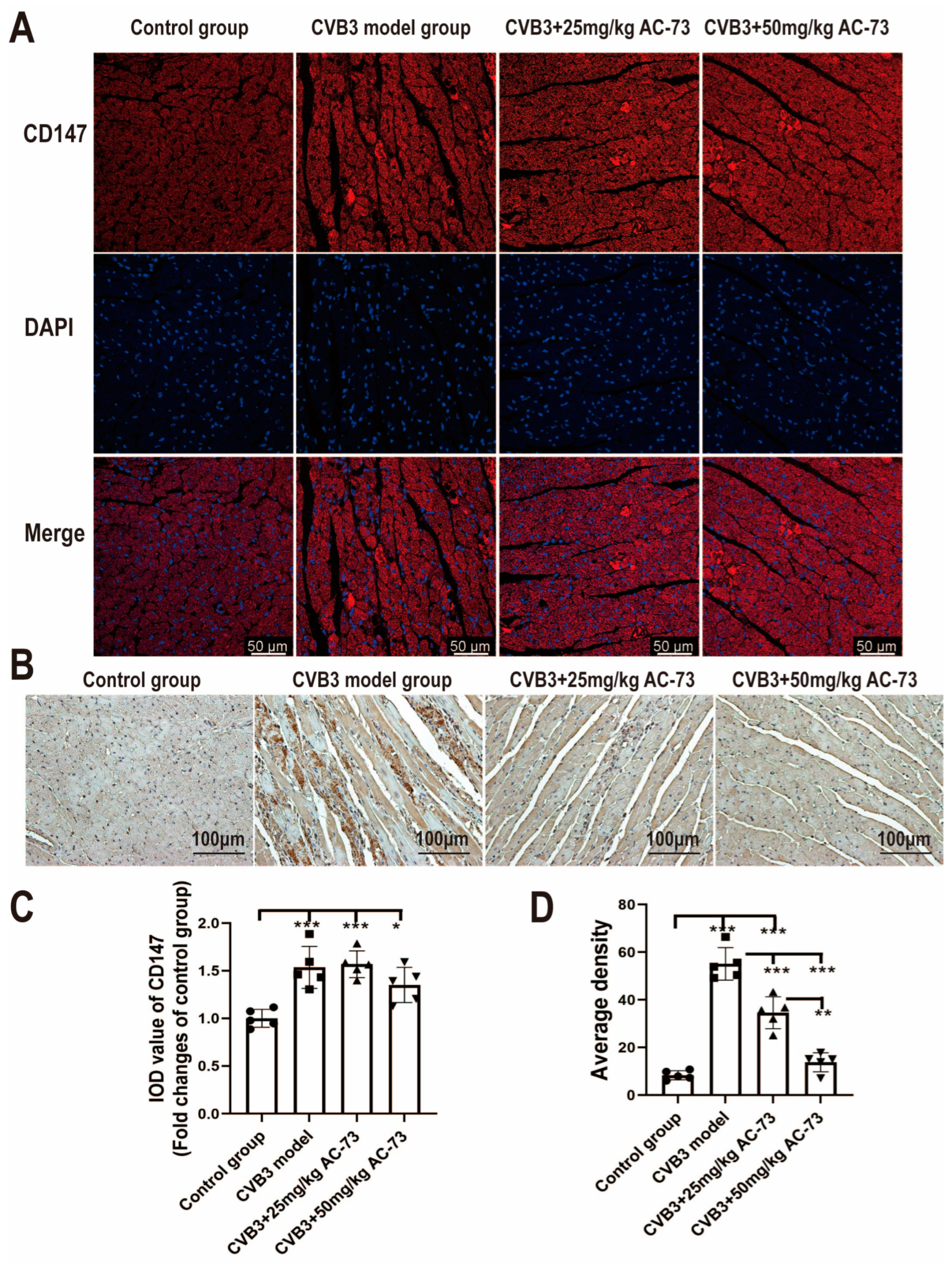
3.1. AC-73 Did Not Affect the Expression of CD147 Induced by CVB3 Infection
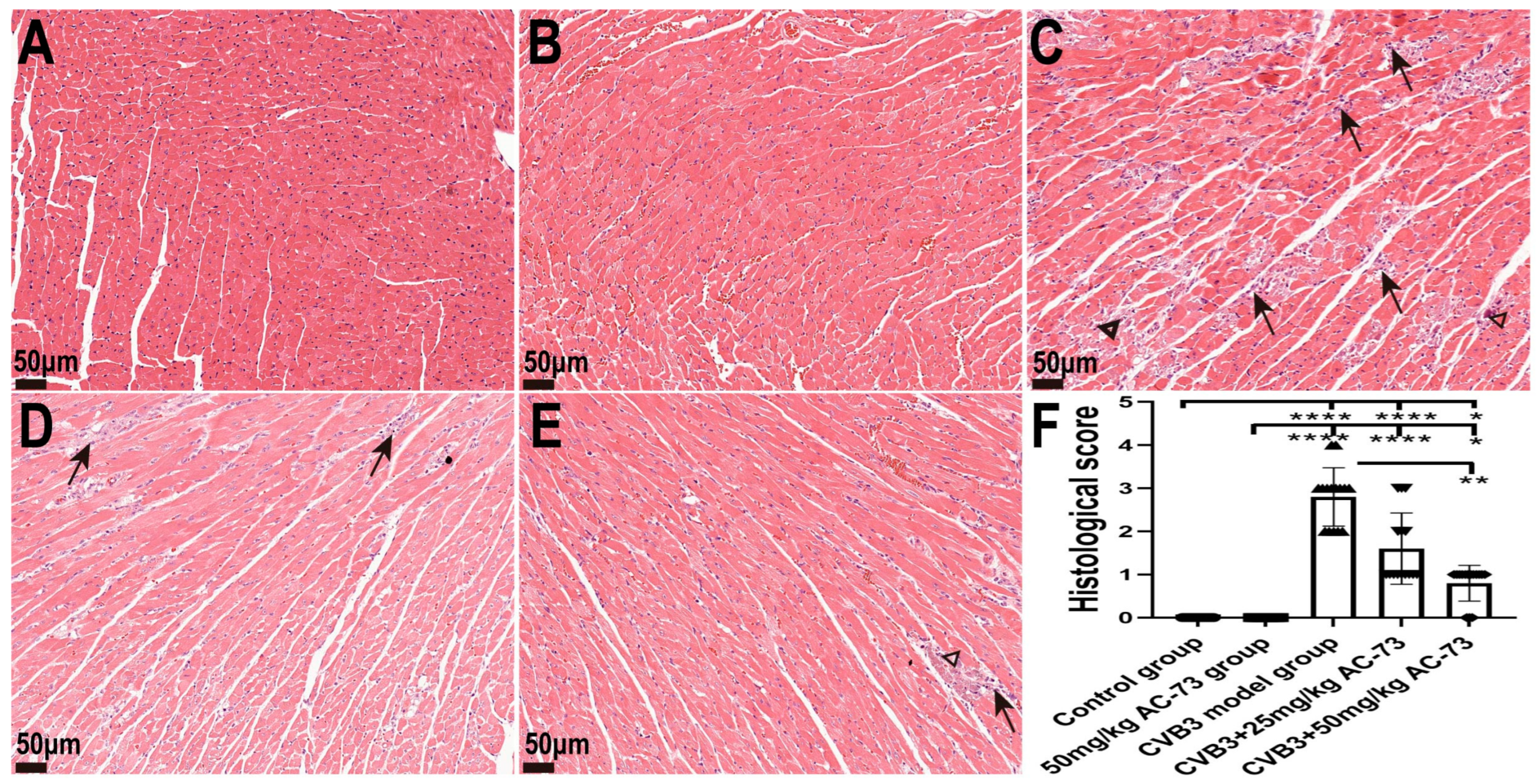
3.2. AC-73 Alleviated Pathological Injury in the Heart
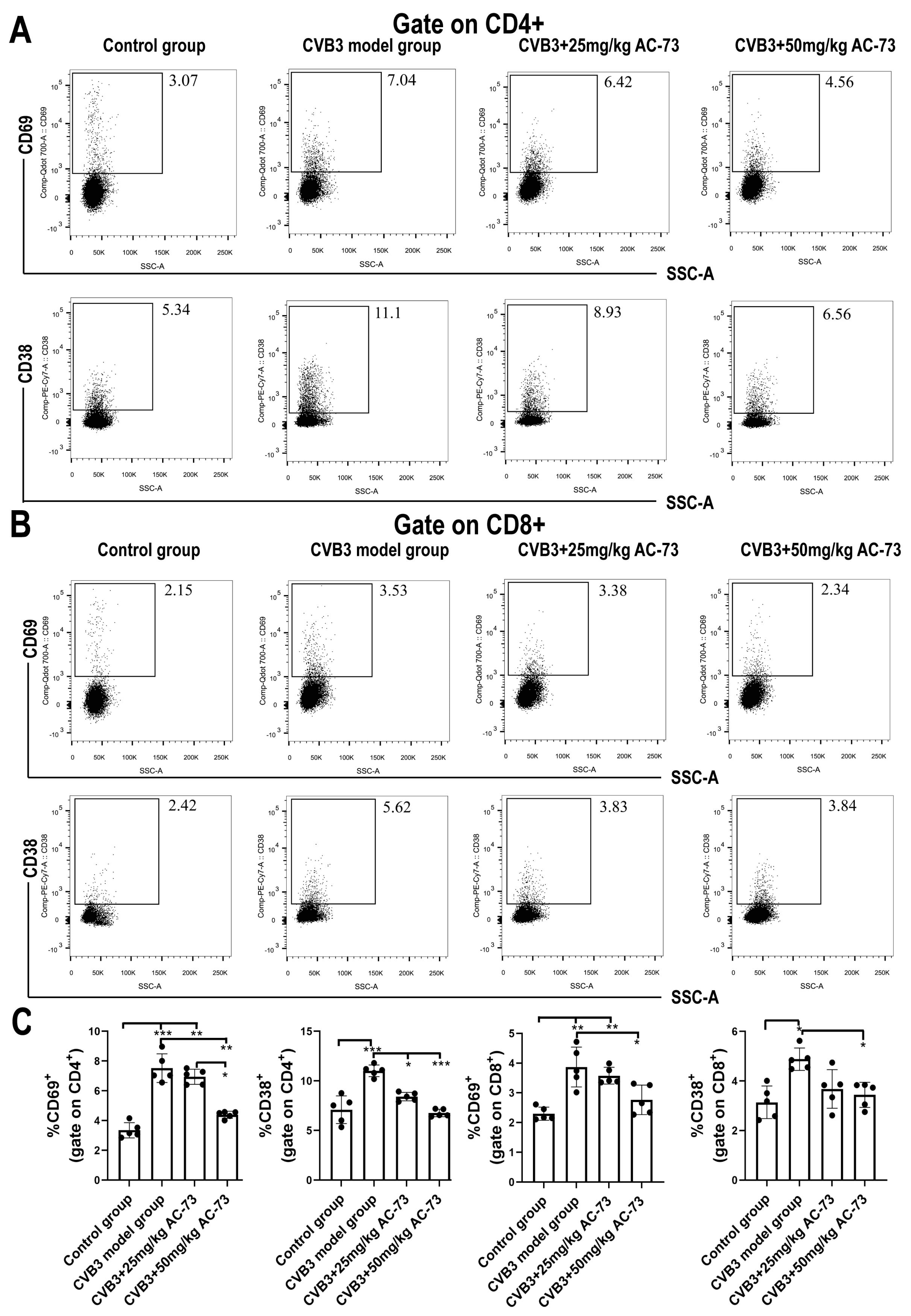
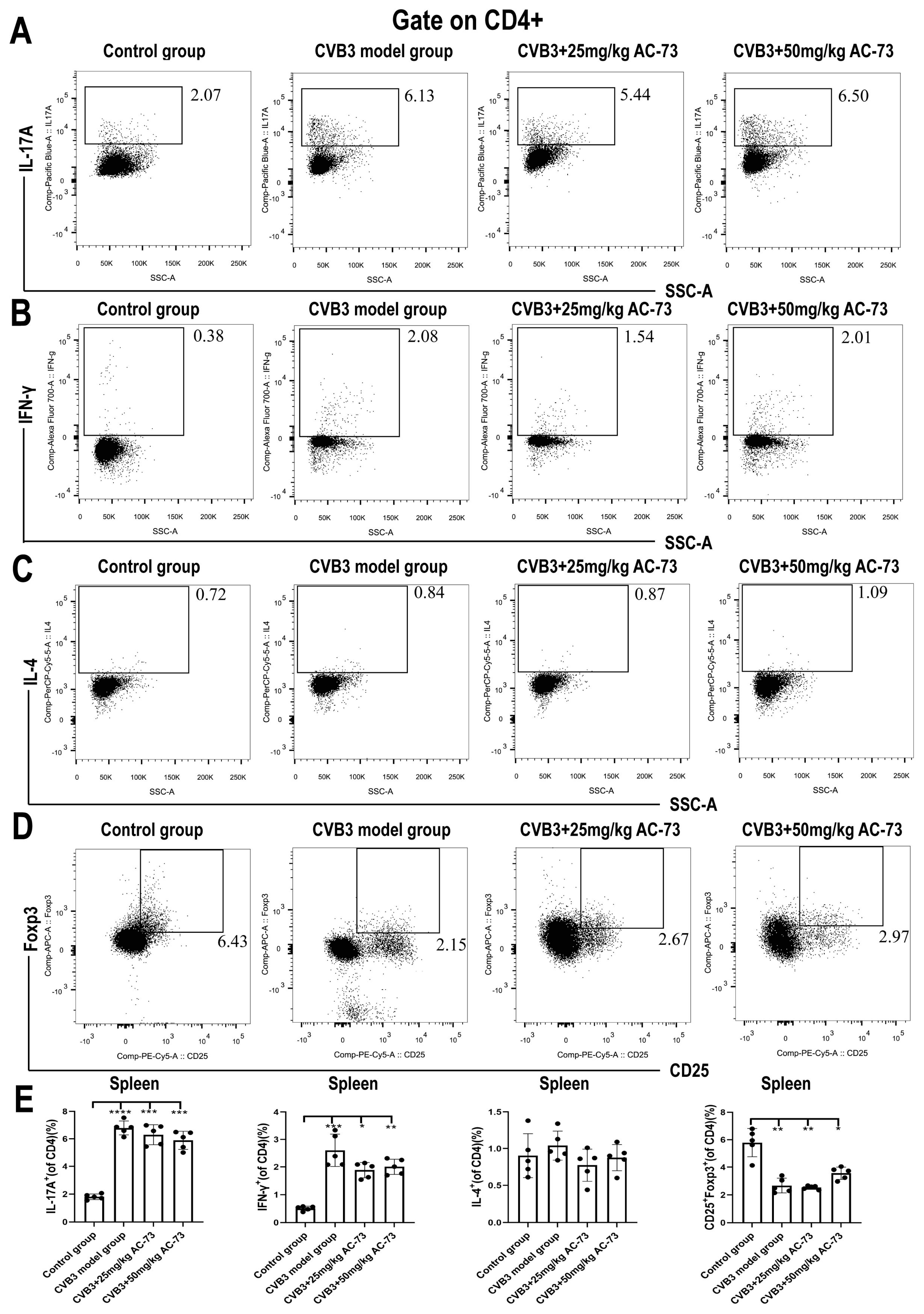
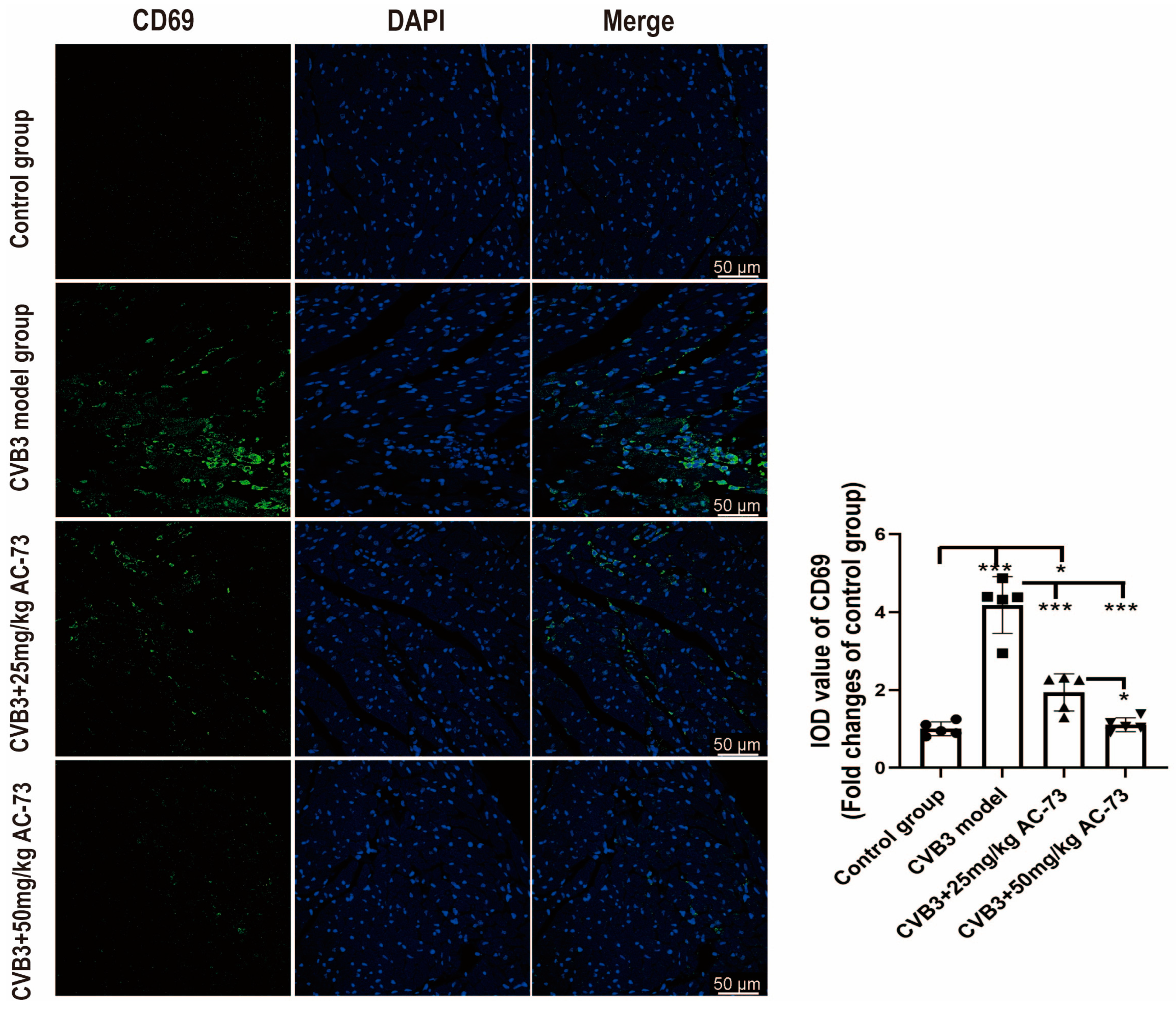
3.3. AC-73 Reduced the Proportion of Activating T Cells Induced by CVB3 Infection
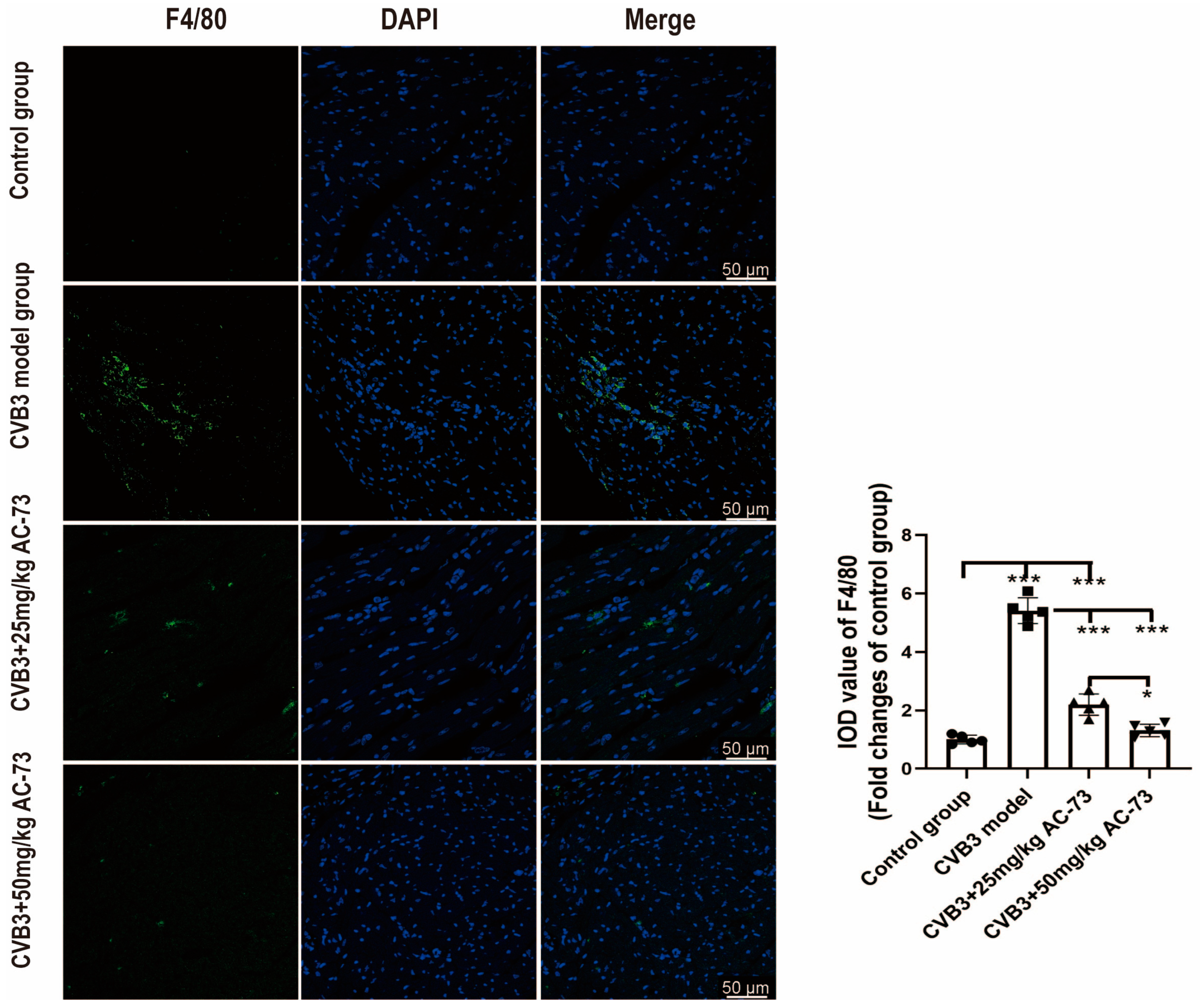
3.4. AC-73 Inhibited the Infiltration of Macrophages in CVB3-Infected Mice
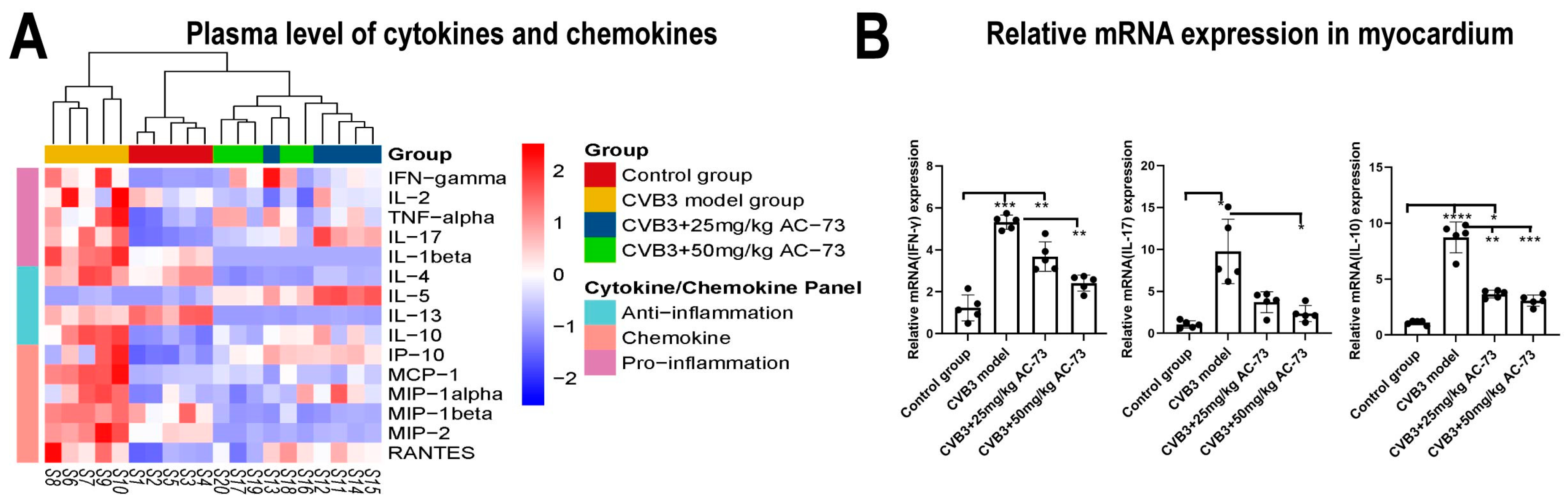
3.5. AC-73 Inhibited the Expression of Cytokines and Chemokines in CVB3-Infected Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caforio, A.L.; Pankuweit, S.; Arbustini, E.; Basso, C.; Gimeno-Blanes, J.; Felix, S.B.; Fu, M.; Heliö, T.; Heymans, S.; Jahns, R.; et al. Current state of knowledge on aetiology, diagnosis, management, and therapy of myocarditis: A position statement of the European Society of Cardiology Working Group on Myocardial and Pericardial Diseases. Eur. Heart J. 2013, 34, 2636–2648. [Google Scholar] [CrossRef] [PubMed]
- Cooper, L.T., Jr. Myocarditis. N. Engl. J. Med. 2009, 360, 1526–1538. [Google Scholar] [CrossRef] [PubMed]
- Sagar, S.; Liu, P.P.; Cooper, L.T., Jr. Myocarditis. Lancet 2012, 379, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Brodehl, A.; Belke, D.D.; Garnett, L.; Martens, K.; Abdelfatah, N.; Rodriguez, M.; Diao, C.; Chen, Y.X.; Gordon, P.M.; Nygren, A.; et al. Transgenic mice overexpressing desmocollin-2 (DSC2) develop cardiomyopathy associated with myocardial inflammation and fibrotic remodeling. PLoS ONE 2017, 12, e0174019. [Google Scholar] [CrossRef]
- Trachtenberg, B.H.; Hare, J.M. Inflammatory Cardiomyopathic Syndromes. Circ. Res. 2017, 121, 803–818. [Google Scholar] [CrossRef]
- Esfandiarei, M.; McManus, B.M. Molecular biology and pathogenesis of viral myocarditis. Annu. Rev. Pathol. 2008, 3, 127–155. [Google Scholar] [CrossRef]
- Carai, P.; Florit Gonzalez, L.; Van Bruggen, S.; Spalart, V.; De Giorgio, D.; Geuens, N.; Martinod, K.; Jones, E.A.V.; Heymans, S. Neutrophil Inhibition Improves Acute Inflammation in a Murine Model of Viral Myocarditis. Cardiovasc. Res. 2022, 118, 3331–3345. [Google Scholar] [CrossRef]
- Pollack, A.; Kontorovich, A.R.; Fuster, V.; Dec, G.W. Viral myocarditis—Diagnosis, treatment options, and current controversies. Nat. Rev. Cardiol. 2015, 12, 670–680. [Google Scholar] [CrossRef]
- Zhao, L.; Fu, Z. Roles of Host Immunity in Viral Myocarditis and Dilated Cardiomyopathy. J. Immunol. Res. 2018, 2018, 5301548. [Google Scholar] [CrossRef]
- Chen, J.; Yang, F.; Shi, S.; Liu, X.; Qin, F.; Wei, X.; Huang, Y.; Liang, W.; Miao, L. The Severity of CVB3-Induced Myocarditis Can Be Improved by Blocking the Orchestration of NLRP3 and Th17 in Balb/c Mice. Mediat. Inflamm. 2021, 2021, 5551578. [Google Scholar] [CrossRef]
- Gopal, R.; Marinelli, M.A.; Alcorn, J.F. Immune Mechanisms in Cardiovascular Diseases Associated with Viral Infection. Front. Immunol. 2020, 11, 570681. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, M.; Niu, H.; Liu, J.; Ruilian, M.; Wang, Y.; Xiao, Y.; Xiao, Z.; Sun, J.; Dong, Y.; et al. Astragalus polysaccharide from Astragalus Melittin ameliorates inflammation via suppressing the activation of TLR-4/NF-κB p65 signal pathway and protects mice from CVB3-induced virus myocarditis. Int. J. Biol. Macromol. 2019, 126, 179–186. [Google Scholar] [CrossRef]
- Yao, H.L.; Song, J.; Sun, P.; Song, Q.Q.; Sheng, L.J.; Chi, M.M.; Han, J. Gene expression analysis during recovery process indicates the mechanism for innate immune injury and repair from Coxsackievirus B3-induced myocarditis. Virus Res. 2016, 213, 314–321. [Google Scholar] [CrossRef]
- Seizer, P.; Klingel, K.; Sauter, M.; Westermann, D.; Ochmann, C.; Schönberger, T.; Schleicher, R.; Stellos, K.; Schmidt, E.M.; Borst, O.; et al. Cyclophilin A affects inflammation, virus elimination and myocardial fibrosis in coxsackievirus B3-induced myocarditis. J. Mol. Cell. Cardiol. 2012, 53, 6–14. [Google Scholar] [CrossRef]
- Arora, K.; Gwinn, W.M.; Bower, M.A.; Watson, A.; Okwumabua, I.; MacDonald, H.R.; Bukrinsky, M.I.; Constant, S.L. Extracellular cyclophilins contribute to the regulation of inflammatory responses. J. Immunol. 2005, 175, 517–522. [Google Scholar] [CrossRef]
- Yurchenko, V.; Constant, S.; Bukrinsky, M. Dealing with the family: CD147 interactions with cyclophilins. Immunology 2006, 117, 301–309. [Google Scholar] [CrossRef]
- Agrawal, S.M.; Silva, C.; Wang, J.; Tong, J.P.; Yong, V.W. A novel anti-EMMPRIN function-blocking antibody reduces T cell proliferation and neurotoxicity: Relevance to multiple sclerosis. J. Neuroinflamm. 2012, 9, 64. [Google Scholar] [CrossRef]
- Gwinn, W.M.; Damsker, J.M.; Falahati, R.; Okwumabua, I.; Kelly-Welch, A.; Keegan, A.D.; Vanpouille, C.; Lee, J.J.; Dent, L.A.; Leitenberg, D.; et al. Novel approach to inhibit asthma-mediated lung inflammation using anti-CD147 intervention. J. Immunol. 2006, 177, 4870–4879. [Google Scholar] [CrossRef]
- Jia, J.; Wang, C.; Shi, Z.; Zhao, J.; Jia, Y.; Zhao-Hui, Z.; Li, X.; Chen, Z.; Zhu, P. Inhibitory effect of CD147/HAb18 monoclonal antibody on cartilage erosion and synovitis in the SCID mouse model for rheumatoid arthritis. Rheumatologys 2009, 48, 721–726. [Google Scholar] [CrossRef]
- Qing, J.; Wang, Y.; Sun, Y.; Huang, J.; Yan, W.; Wang, J.; Su, D.; Ni, C.; Li, J.; Rao, Z.; et al. Cyclophilin A associates with enterovirus-71 virus capsid and plays an essential role in viral infection as an uncoating regulator. PLoS Pathog. 2014, 10, e1004422. [Google Scholar] [CrossRef]
- Pushkarsky, T.; Zybarth, G.; Dubrovsky, L.; Yurchenko, V.; Tang, H.; Guo, H.; Toole, B.; Sherry, B.; Bukrinsky, M. CD147 facilitates HIV-1 infection by interacting with virus-associated cyclophilin A. Proc. Natl. Acad. Sci. USA 2001, 98, 6360–6365. [Google Scholar] [CrossRef] [PubMed]
- Heldin, C.H. Dimerization of cell surface receptors in signal transduction. Cell 1995, 80, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Koch, C.; Staffler, G.; Hüttinger, R.; Hilgert, I.; Prager, E.; Cerný, J.; Steinlein, P.; Majdic, O.; Horejsí, V.; Stockinger, H. T cell activation-associated epitopes of CD147 in regulation of the T cell response, and their definition by antibody affinity and antigen density. Int. Immunol. 1999, 11, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.Q.; Lu, M.Z.; Song, J.; Chi, M.M.; Sheng, L.J.; Yu, J.; Luo, X.N.; Zhang, L.; Yao, H.L.; Han, J. Coxsackievirus B3 2A protease promotes encephalomyocarditis virus replication. Virus Res. 2015, 208, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.G.; Wang, L.; Cui, H.Y.; Peng, J.L.; Wang, S.J.; Geng, J.J.; Liu, J.D.; Feng, F.; Song, F.; Li, L.; et al. A novel small-molecule compound targeting CD147 inhibits the motility and invasion of hepatocellular carcinoma cells. Oncotarget 2016, 7, 9429–9447. [Google Scholar] [CrossRef]
- Yen, M.H.; Tsao, K.C.; Huang, Y.C.; Huang, C.G.; Huang, Y.L.; Lin, R.; Chang, M.L.; Huang, C.C.; Yan, D.C.; Lin, T.Y. Viral load in blood is correlated with disease severity of neonatal coxsackievirus B3 infection: Early diagnosis and predicting disease severity is possible in severe neonatal enterovirus infection. Clin. Infect. Dis. 2007, 44, e78–e81. [Google Scholar] [CrossRef]
- Qiao, J.; Li, W.; Bao, J.; Peng, Q.; Wen, D.; Wang, J.; Sun, B. The expression of SARS-CoV-2 receptor ACE2 and CD147, and protease TMPRSS2 in human and mouse brain cells and mouse brain tissues. Biochem. Biophys. Res. Commun. 2020, 533, 867–871. [Google Scholar] [CrossRef]
- Szalay, G.; Sauter, M.; Hald, J.; Weinzierl, A.; Kandolf, R.; Klingel, K. Sustained nitric oxide synthesis contributes to immunopathology in ongoing myocarditis attributable to interleukin-10 disorders. Am. J. Pathol. 2006, 169, 2085–2093. [Google Scholar] [CrossRef]
- Pappritz, K.; Savvatis, K.; Miteva, K.; Kerim, B.; Dong, F.; Fechner, H.; Müller, I.; Brandt, C.; Lopez, B.; González, A.; et al. Immunomodulation by adoptive regulatory T-cell transfer improves Coxsackievirus B3-induced myocarditis. FASEB J. 2018, 32, 6066–6078. [Google Scholar] [CrossRef]
- Sesti-Costa, R.; Silva, G.K.; Proenca-Modena, J.L.; Carlos, D.; Silva, M.L.; Alves-Filho, J.C.; Arruda, E.; Liew, F.Y.; Silva, J.S. The IL-33/ST2 pathway controls coxsackievirus B5-induced experimental pancreatitis. J. Immunol. 2013, 191, 283–292. [Google Scholar] [CrossRef]
- Liu, K.; Zhao, E.; Ilyas, G.; Lalazar, G.; Lin, Y.; Haseeb, M.; Tanaka, K.E.; Czaja, M.J. Impaired macrophage autophagy increases the immune response in obese mice by promoting proinflammatory macrophage polarization. Autophagy 2015, 11, 271–284. [Google Scholar] [CrossRef]
- Pan, H.; Zhang, Y.; Luo, Z.; Li, P.; Liu, L.; Wang, C.; Wang, H.; Li, H.; Ma, Y. Autophagy mediates avian influenza H5N1 pseudotyped particle-induced lung inflammation through NF-κB and p38 MAPK signaling pathways. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, L183–L195. [Google Scholar] [CrossRef]
- Wainwright, D.A.; Sengupta, S.; Han, Y.; Ulasov, I.V.; Lesniak, M.S. The presence of IL-17A and T helper 17 cells in experimental mouse brain tumors and human glioma. PLoS ONE 2010, 5, e15390. [Google Scholar] [CrossRef]
- Keyes, L.R.; Bego, M.G.; Soland, M.; St Jeor, S. Cyclophilin A is required for efficient human cytomegalovirus DNA replication and reactivation. J. Gen. Virol. 2012, 93, 722–732. [Google Scholar] [CrossRef]
- Zhong, Z.; Ning, J.; Boggs, E.A.; Jang, S.; Wallace, C.; Telmer, C.; Bruchez, M.P.; Ahn, J.; Engelman, A.N.; Zhang, P.; et al. Cytoplasmic CPSF6 Regulates HIV-1 Capsid Trafficking and Infection in a Cyclophilin A-Dependent Manner. mBio 2021, 12, e03142-20. [Google Scholar] [CrossRef]
- Hahn, J.N.; Kaushik, D.K.; Yong, V.W. The role of EMMPRIN in T cell biology and immunological diseases. J. Leukoc. Biol. 2015, 98, 33–48. [Google Scholar] [CrossRef]
- Tracy, S.; Liu, H.L.; Chapman, N.M. Coxsackievirus B3: Primary structure of the 5′ non-coding and capsid protein-coding regions of the genome. Virus Res. 1985, 3, 263–270. [Google Scholar] [CrossRef]
- Chau, D.H.; Yuan, J.; Zhang, H.; Cheung, P.; Lim, T.; Liu, Z.; Sall, A.; Yang, D. Coxsackievirus B3 proteases 2A and 3C induce apoptotic cell death through mitochondrial injury and cleavage of eIF4GI but not DAP5/p97/NAT1. Apoptosis Int. J. Program. Cell Death 2007, 12, 513–524. [Google Scholar] [CrossRef]
- Lee, K.Y. A common immunopathogenesis mechanism for infectious diseases: The protein-homeostasis-system hypothesis. Infect. Chemother. 2015, 47, 12–26. [Google Scholar] [CrossRef]
- Lee, K.Y. Common immunopathogenesis of central nervous system diseases: The protein-homeostasis-system hypothesis. Cell Biosci. 2022, 12, 184. [Google Scholar] [CrossRef]
- Wang, X.; Li, M.; Xie, Y.; Yu, Y.; Liu, G.; Yu, Y.; Yang, X.; Zou, Y.; Ge, J.; Chen, R. The frequency of invariant natural killer T cells correlates with the severity of myocarditis. Viral Immunol. 2014, 27, 88–95. [Google Scholar] [CrossRef] [PubMed]
- De-Pu, Z.; Li-Sha, G.; Guang-Yi, C.; Xiaohong, G.; Chao, X.; Cheng, Z.; Wen-Wu, Z.; Jia, L.; Jia-Feng, L.; Maoping, C.; et al. The cholinergic anti-inflammatory pathway ameliorates acute viral myocarditis in mice by regulating CD4(+) T cell differentiation. Virulence 2018, 9, 1364–1376. [Google Scholar] [CrossRef] [PubMed]
- Fairweather, D.; Stafford, K.A.; Sung, Y.K. Update on coxsackievirus B3 myocarditis. Curr. Opin. Rheumatol. 2012, 24, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Seizer, P.; Geisler, T.; Bigalke, B.; Schneider, M.; Klingel, K.; Kandolf, R.; Stellos, K.; Schreieck, J.; Gawaz, M.; May, A.E. EMMPRIN and its ligand cyclophilin A as novel diagnostic markers in inflammatory cardiomyopathy. Int. J. Cardiol. 2013, 163, 299–304. [Google Scholar] [CrossRef]
- Seizer, P.; Gawaz, M.; May, A.E. Platelet-monocyte interactions—A dangerous liaison linking thrombosis, inflammation and atherosclerosis. Curr. Med. Chem. 2008, 15, 1976–1980. [Google Scholar] [CrossRef]
- Chiampanichayakul, S.; Peng-in, P.; Khunkaewla, P.; Stockinger, H.; Kasinrerk, W. CD147 contains different bioactive epitopes involving the regulation of cell adhesion and lymphocyte activation. Immunobiology 2006, 211, 167–178. [Google Scholar] [CrossRef]
- Maeda, K.; Kosugi, T.; Sato, W.; Kojima, H.; Sato, Y.; Kamimura, D.; Kato, N.; Tsuboi, N.; Yuzawa, Y.; Matsuo, S.; et al. CD147/basigin limits lupus nephritis and Th17 cell differentiation in mice by inhibiting the interleukin-6/STAT-3 pathway. Arthritis Rheumatol. 2015, 67, 2185–2195. [Google Scholar] [CrossRef]
- Okubo, A.; Uchida, Y.; Higashi, Y.; Sato, T.; Ogawa, Y.; Ryuge, A.; Kadomatsu, K.; Kanekura, T. CD147 Is Essential for the Development of Psoriasis via the Induction of Th17 Cell Differentiation. Int. J. Mol. Sci. 2021, 23, 177. [Google Scholar] [CrossRef]
- Bukrinsky, M.I. Cyclophilins: Unexpected messengers in intercellular communications. Trends Immunol. 2002, 23, 323–325. [Google Scholar] [CrossRef]
- Jin, R.; Liu, S.; Wang, M.; Zhong, W.; Li, G. Inhibition of CD147 Attenuates Stroke-Associated Pneumonia through Modulating Lung Immune Response in Mice. Front. Neurol. 2019, 10, 853. [Google Scholar] [CrossRef]
- Guo, N.; Ye, S.; Zhang, K.; Yu, X.; Cui, H.; Yang, X.; Lin, P.; Lv, M.; Miao, J.; Zhang, Y.; et al. A critical epitope in CD147 facilitates memory CD4(+) T-cell hyper-activation in rheumatoid arthritis. Cell. Mol. Immunol. 2019, 16, 568–579. [Google Scholar] [CrossRef]
- Yue-Chun, L.; Gu, X.H.; Li-Sha, G.; Zhou, D.P.; Xing, C.; Guo, X.L.; Pan, L.L.; Song, S.Y.; Yu, L.L.; Chen, G.Y.; et al. Vagus nerve plays a pivotal role in CD4+ T cell differentiation during CVB3-induced murine acute myocarditis. Virulence 2021, 12, 360–376. [Google Scholar] [CrossRef]
- Jin, H.; Guo, X. Valproic acid ameliorates coxsackievirus-B3-induced viral myocarditis by modulating Th17/Treg imbalance. Virol. J. 2016, 13, 168. [Google Scholar] [CrossRef]
- Yuan, J.; Yu, M.; Lin, Q.W.; Cao, A.L.; Yu, X.; Dong, J.H.; Wang, J.P.; Zhang, J.H.; Wang, M.; Guo, H.P.; et al. Th17 cells contribute to viral replication in coxsackievirus B3-induced acute viral myocarditis. J. Immunol. 2010, 185, 4004–4010. [Google Scholar] [CrossRef]
- Butera, A.; Quaranta, M.T.; Crippa, L.; Spinello, I.; Saulle, E.; Di Carlo, N.; Campanile, D.; Boirivant, M.; Labbaye, C. CD147 Targeting by AC-73 Induces Autophagy and Reduces Intestinal Fibrosis Associated with TNBS Chronic Colitis. J. Crohn’s Colitis 2022, 16, 1751–1761. [Google Scholar] [CrossRef]
- Pushkarsky, T.; Yurchenko, V.; Laborico, A.; Bukrinsky, M. CD147 stimulates HIV-1 infection in a signal-independent fashion. Biochem. Biophys. Res. Commun. 2007, 363, 495–499. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Zong, K.; Song, J.; Song, Q.; Xia, D.; Liu, M.; Du, H.; Xia, Z.; Yao, H.; Han, J. Inhibitor of CD147 Suppresses T Cell Activation and Recruitment in CVB3-Induced Acute Viral Myocarditis. Viruses 2023, 15, 1137. https://doi.org/10.3390/v15051137
Wang R, Zong K, Song J, Song Q, Xia D, Liu M, Du H, Xia Z, Yao H, Han J. Inhibitor of CD147 Suppresses T Cell Activation and Recruitment in CVB3-Induced Acute Viral Myocarditis. Viruses. 2023; 15(5):1137. https://doi.org/10.3390/v15051137
Chicago/Turabian StyleWang, Ruifang, Kexin Zong, Juan Song, Qinqin Song, Dong Xia, Mi Liu, Haijun Du, Zhiqiang Xia, Hailan Yao, and Jun Han. 2023. "Inhibitor of CD147 Suppresses T Cell Activation and Recruitment in CVB3-Induced Acute Viral Myocarditis" Viruses 15, no. 5: 1137. https://doi.org/10.3390/v15051137