
Allicin Inhibits Porcine Reproductive and Respiratory Syndrome Virus Infection In Vitro and Alleviates Inflammatory Responses
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses, Antibodies, and Compounds
2.2. Cytotoxicity Assay
2.3. Quantitative Real-Time PCR (qRT-PCR)
2.4. Western Blot Analysis
2.5. Immunofluorescence Assay (IFA)
2.6. Gaussia Luciferase Assay
2.7. In Vitro Viricidal Assay
2.8. Time Course Study of the Antiviral Effect of Allicin on PRRSV Infection
2.9. Viral Attachment, Entry, Replication, Assembly, and Release Assays
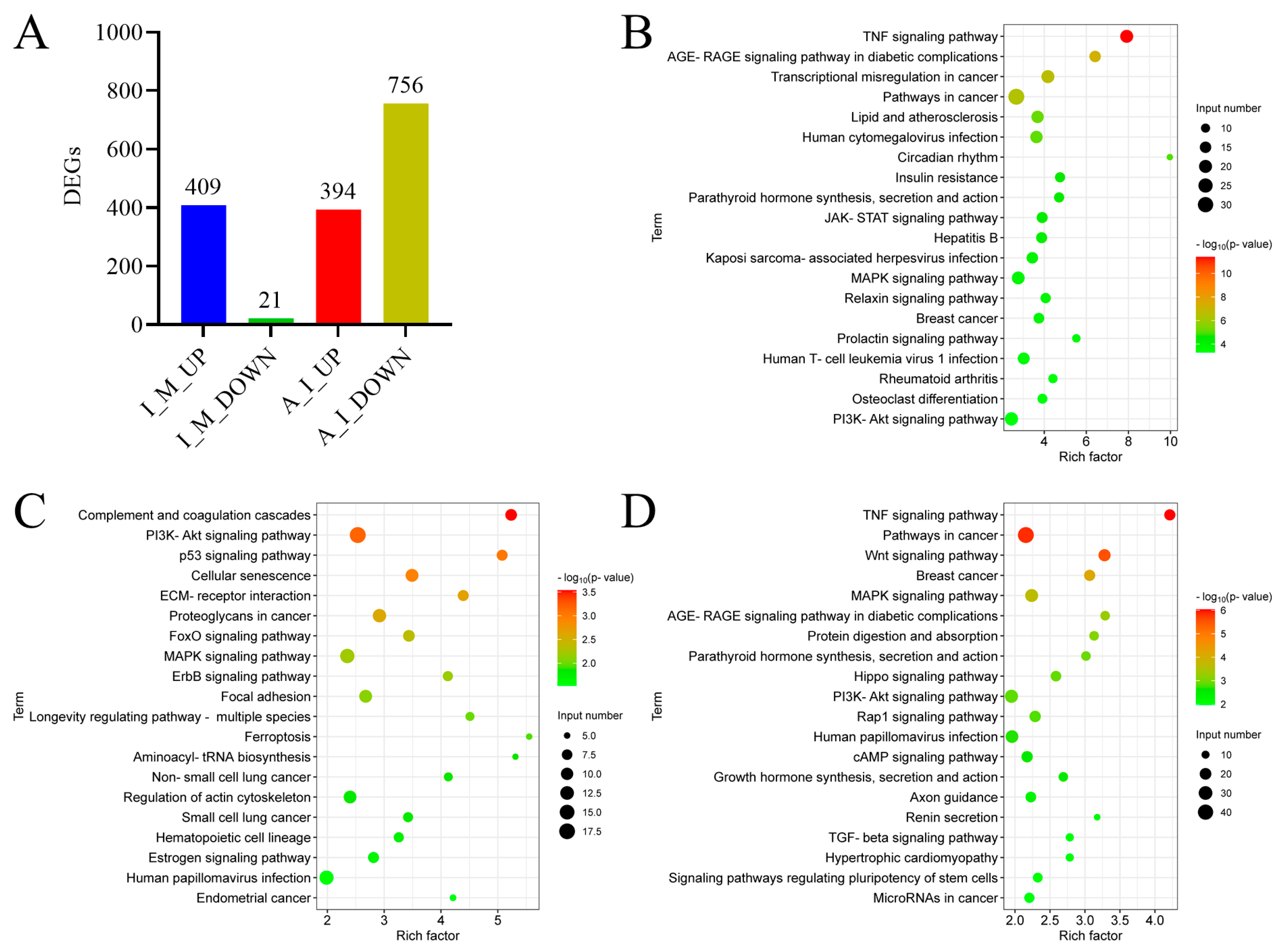
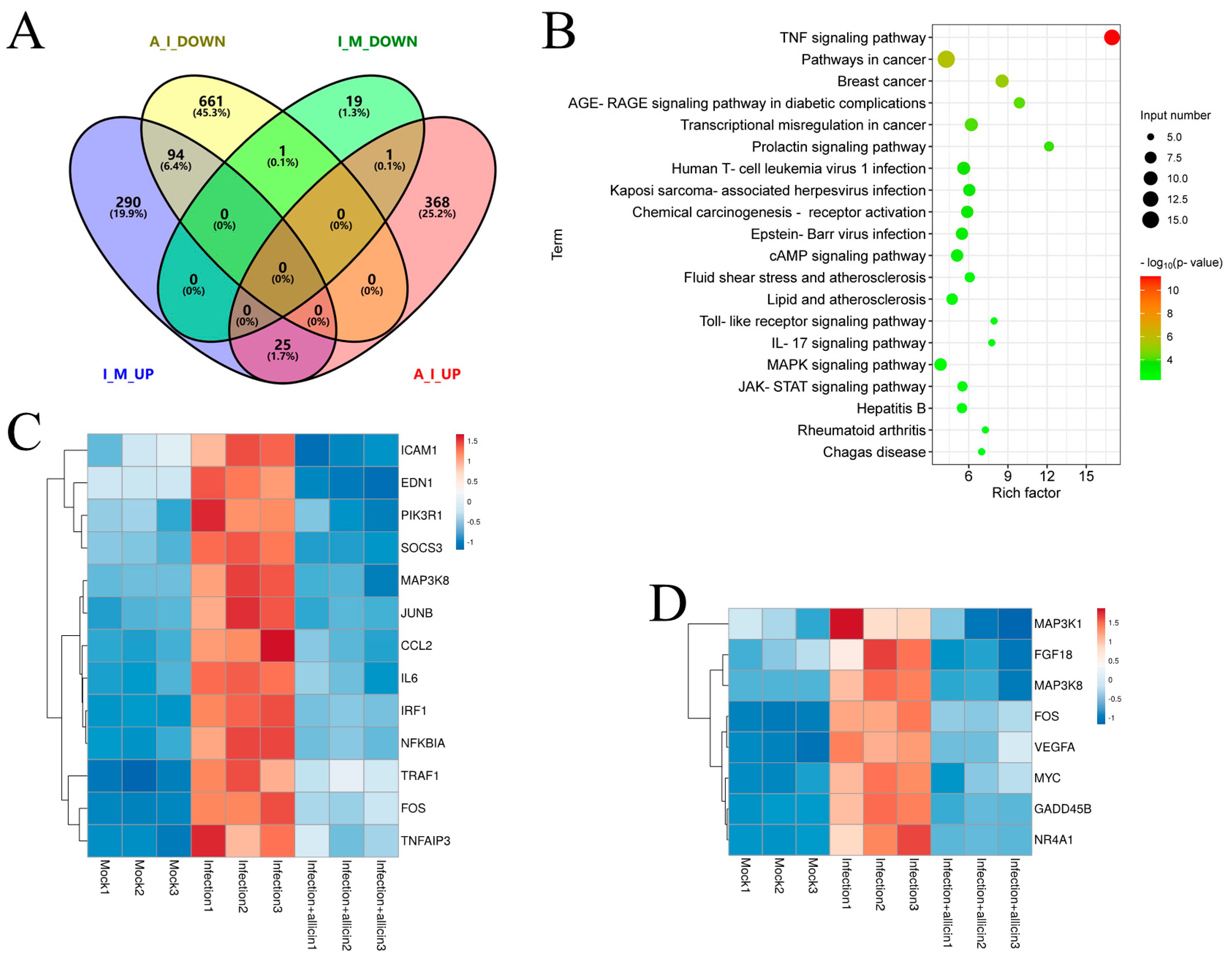
2.10. Transcriptome Analysis
2.11. Statistical Analysis
3. Results
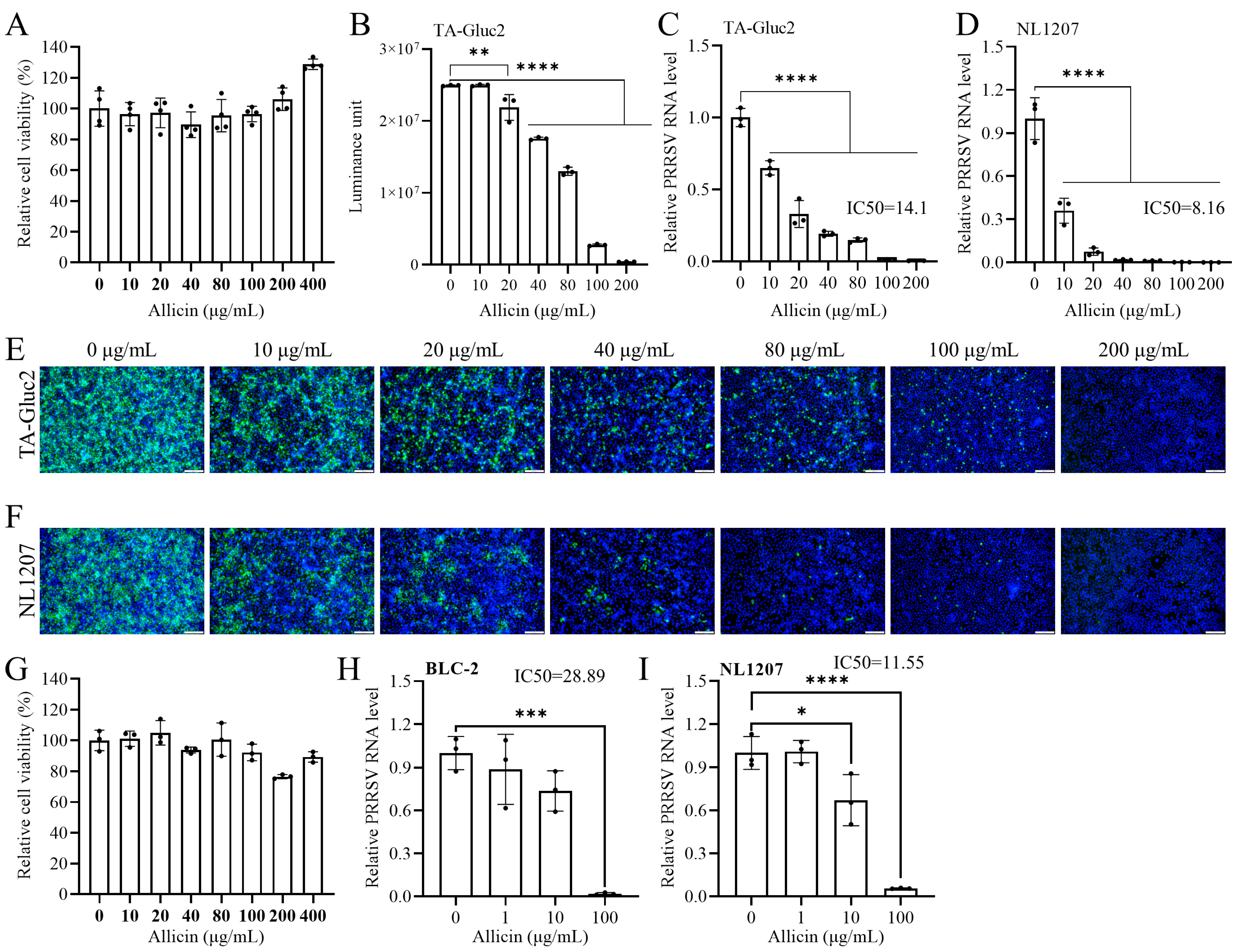
3.1. Allicin Inhibits the Infection of HP-PRRSV and NADC30-like PRRSV Strains in MARC-145 and PAM
3.2. Allicin Does Not Have a Significant Virucidal Effect on PRRSV
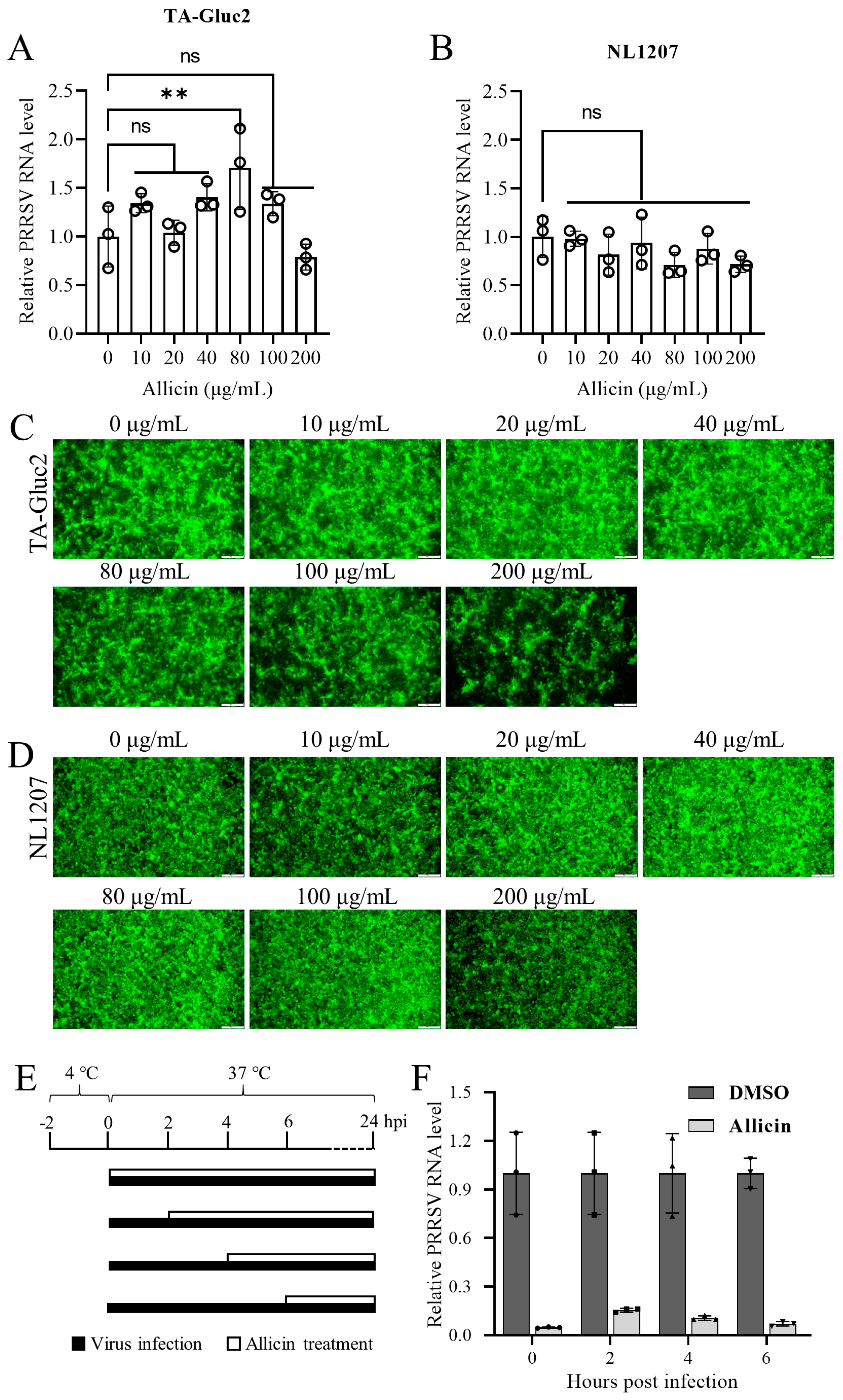
3.3. Allicin Exhibits Inhibitory Effects on Multiple Stages of the PRRSV Life Cycle
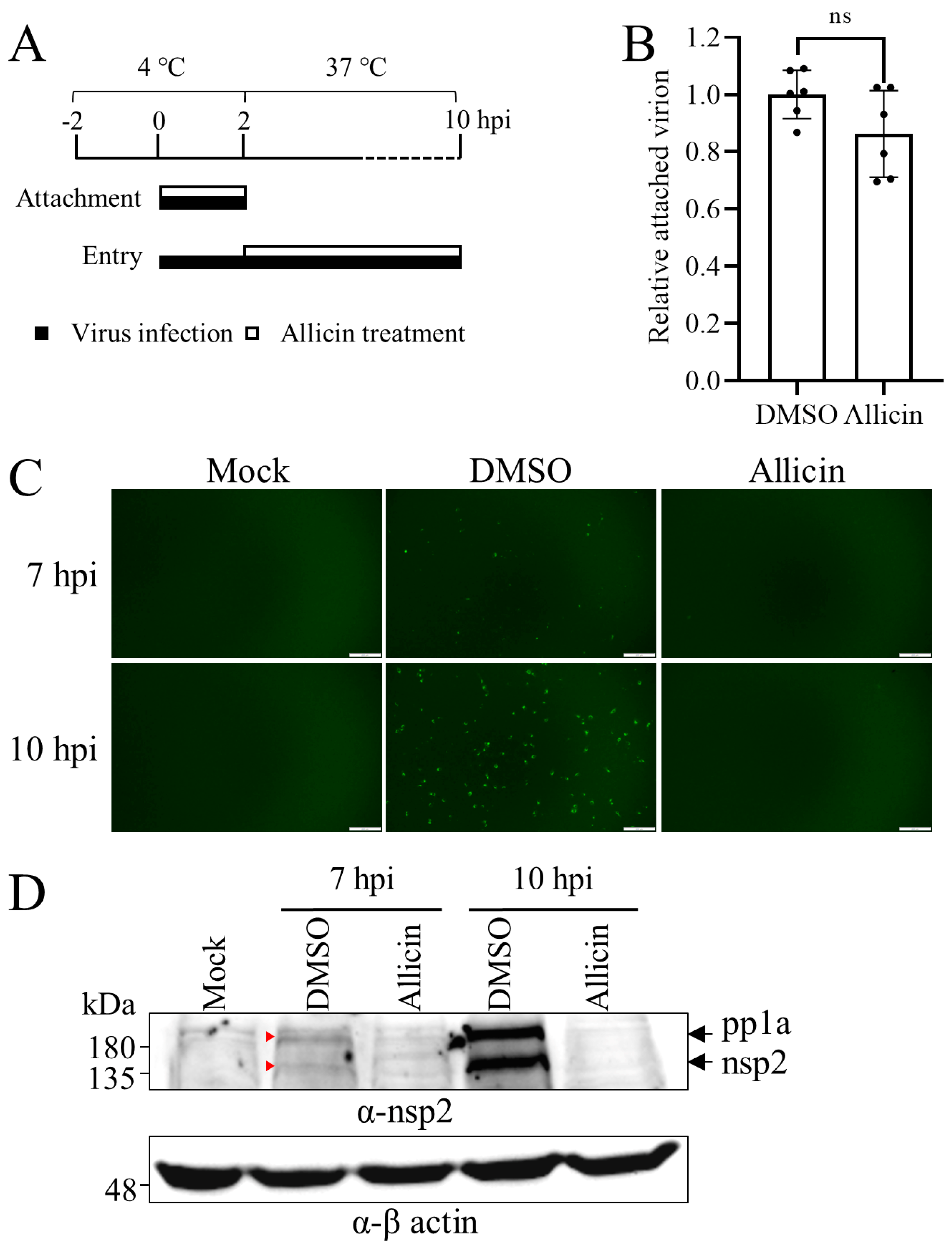
3.4. Allicin Alleviates PRRSV Entry, but Not Attachment
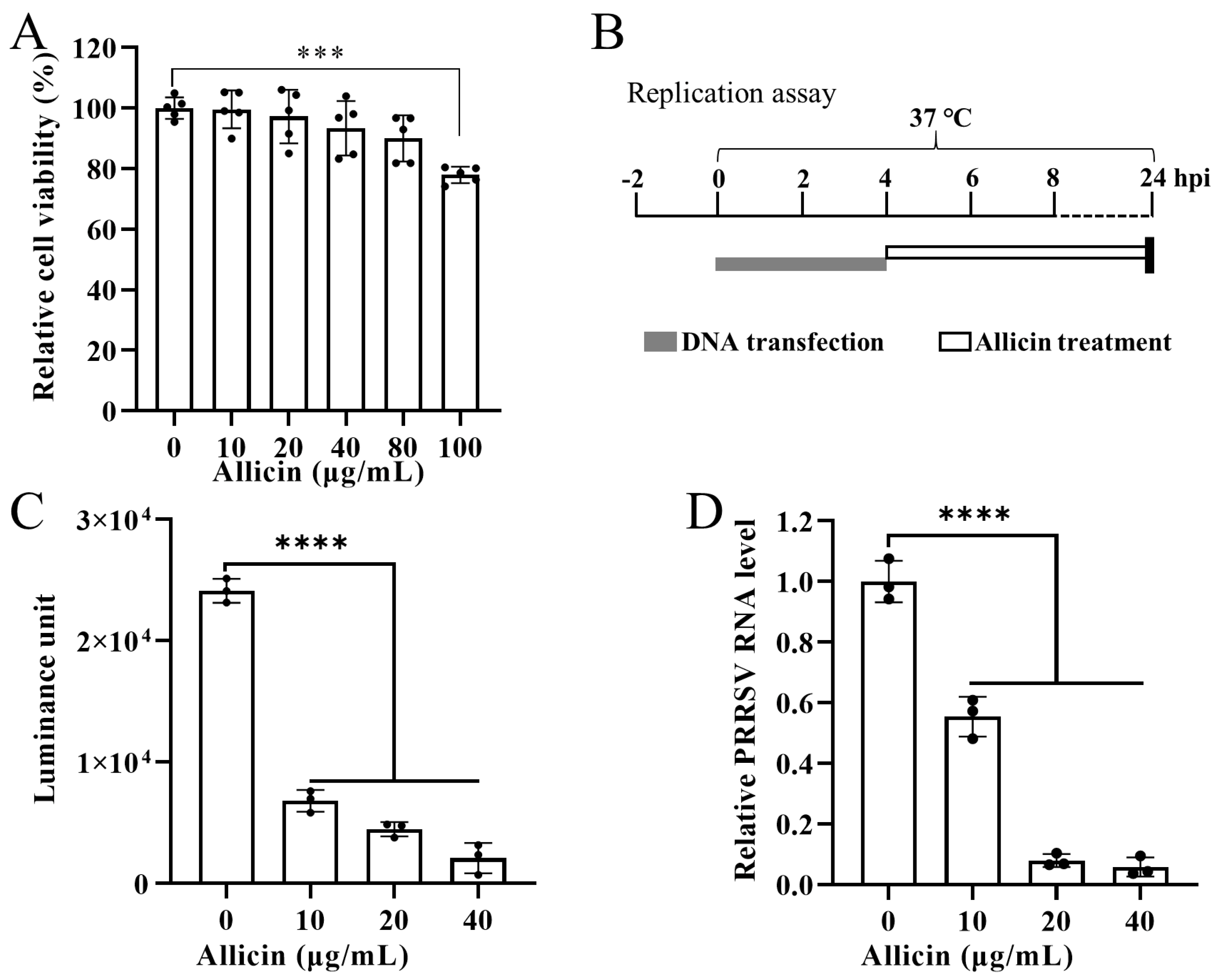
3.5. Allicin Suppresses PRRSV Replication in BHK-21 Cells
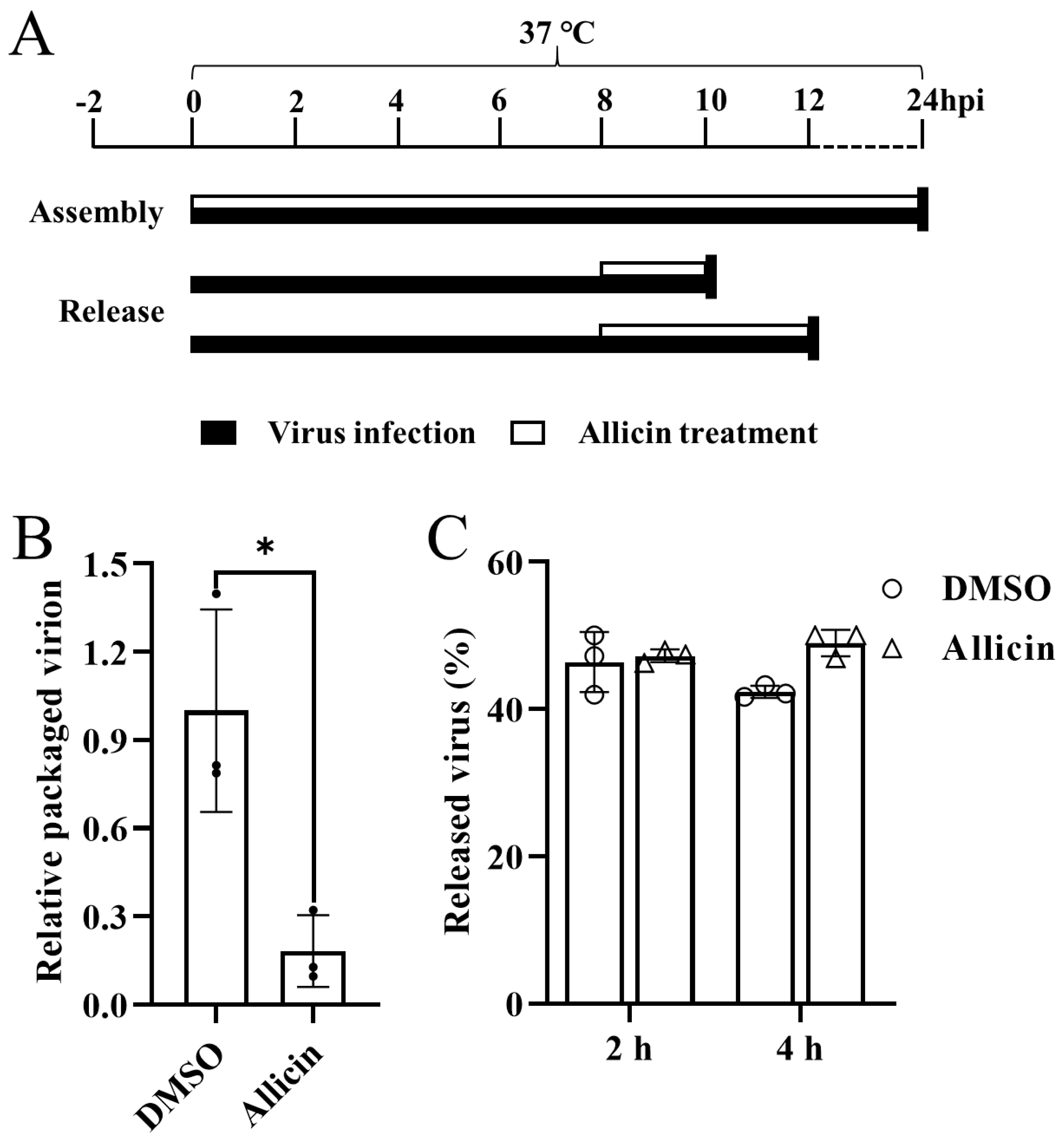
3.6. Allicin Blocks the Packaging of the PRRSV Genome but Does Not Inhibit the Release of PRRSV Progeny
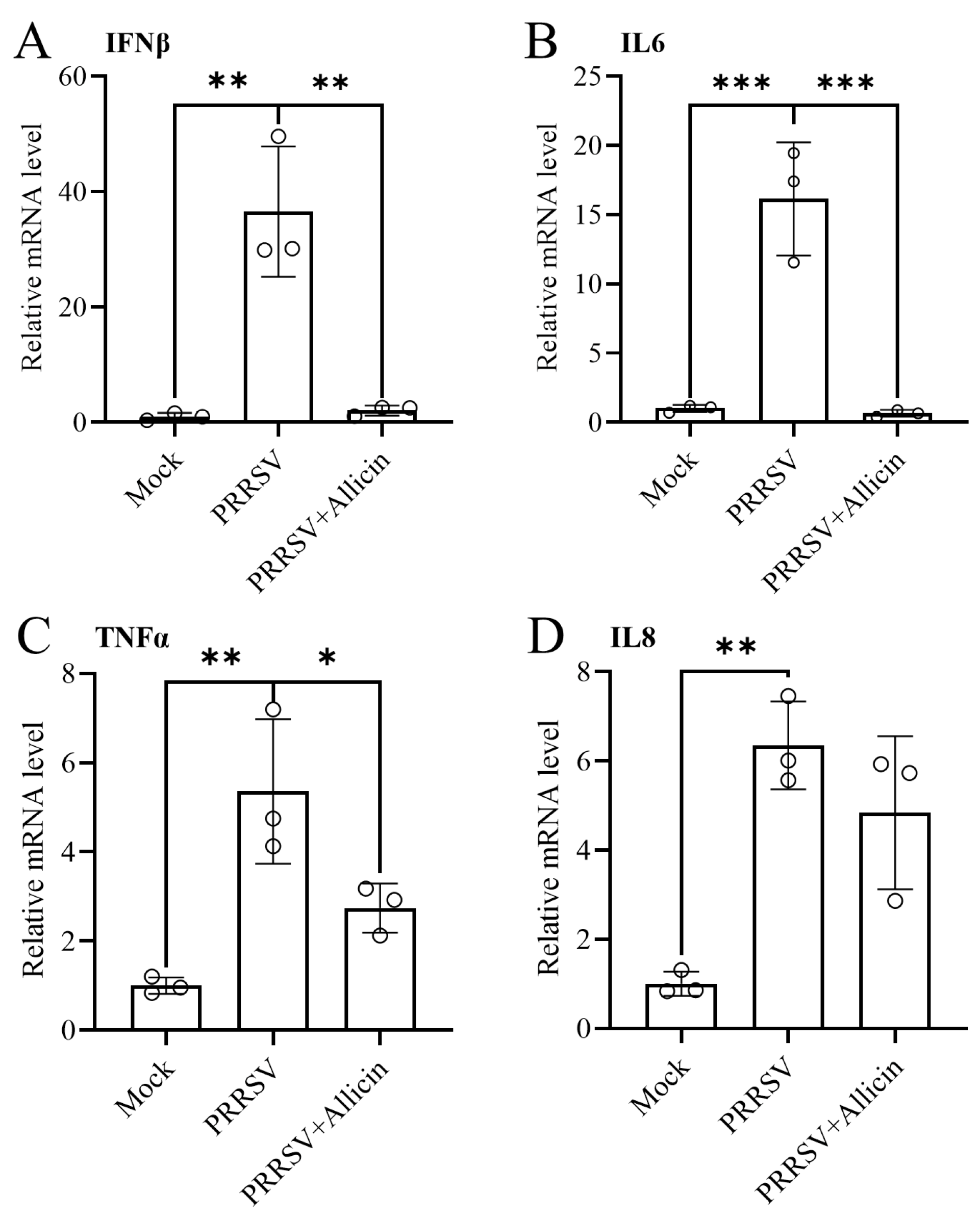
3.7. Allicin Ameliorates PRRSV-Induced Inflammatory Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thanawongnuwech, R.; Thacker, B.; Halbur, P.; Thacker, E.L. Increased production of proinflammatory cytokines following infection with porcine reproductive and respiratory syndrome virus and Mycoplasma hyopneumoniae. Clin. Diagn. Lab. Immunol. 2004, 11, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, S.; Li, C.; Wang, C.; Liu, Y.; Wang, G.; He, X.; Hu, L.; Liu, Y.; Cui, M.; et al. Secondary Haemophilus parasuis infection enhances highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) infection-mediated inflammatory responses. Vet. Microbiol. 2017, 204, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; ter Laak, E.A.; Bloemraad, M.; de Kluyver, E.P.; Kragten, C.; van Buiten, L.; den Besten, A.; Wagenaar, F.; et al. Mystery swine disease in The Netherlands: The isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Hanada, K.; Suzuki, Y.; Nakane, T.; Hirose, O.; Gojobori, T. The origin and evolution of porcine reproductive and respiratory syndrome viruses. Mol. Biol. Evol. 2005, 22, 1024–1031. [Google Scholar] [CrossRef]
- Murtaugh, M.P.; Stadejek, T.; Abrahante, J.E.; Lam, T.T.; Leung, F.C. The ever-expanding diversity of porcine reproductive and respiratory syndrome virus. Virus Res. 2010, 154, 18–30. [Google Scholar] [CrossRef]
- Cui, X.; Xia, D.; Huang, X.; Sun, Y.; Shi, M.; Zhang, J.; Li, G.; Yang, Y.; Wang, H.; Cai, X.; et al. Analysis of Recombinant Characteristics Based on 949 PRRSV-2 Genomic Sequences Obtained from 1991 to 2021 Shows That Viral Multiplication Ability Contributes to Dominant Recombination. Microbiol. Spectr. 2022, 10, e0293422. [Google Scholar] [CrossRef]
- Tian, K.; Yu, X.; Zhao, T.; Feng, Y.; Cao, Z.; Wang, C.; Hu, Y.; Chen, X.; Hu, D.; Tian, X.; et al. Emergence of fatal PRRSV variants: Unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark. PLoS ONE 2007, 2, e526. [Google Scholar] [CrossRef]
- Li, Y.; Ren, C.; Li, C.; Xiao, Y.; Zhou, Y. A Recombinant Porcine Reproductive and Respiratory Syndrome Virus Stably Expressing a Gaussia Luciferase for Antiviral Drug Screening Assay and Luciferase-Based Neutralization Assay. Front. Microbiol. 2022, 13, 907281. [Google Scholar] [CrossRef]
- Xu, H.; Li, C.; Li, W.; Zhao, J.; Gong, B.; Sun, Q.; Tang, Y.D.; Xiang, L.; Leng, C.; Peng, J.; et al. Novel characteristics of Chinese NADC34-like PRRSV during 2020–2021. Transbound. Emerg. Dis. 2022, 69, e3215–e3224. [Google Scholar] [CrossRef]
- Chen, X.X.; Zhou, X.; Guo, T.; Qiao, S.; Guo, Z.; Li, R.; Jin, Q.; Hu, X.; Xing, G.; Deng, R.; et al. Efficacy of a live attenuated highly pathogenic PRRSV vaccine against a NADC30-like strain challenge: Implications for ADE of PRRSV. BMC Vet. Res. 2021, 17, 260. [Google Scholar] [CrossRef]
- Chai, W.; Liu, Z.; Sun, Z.; Su, L.; Zhang, C.; Huang, L. Efficacy of two porcine reproductive and respiratory syndrome (PRRS) modified-live virus (MLV) vaccines against heterologous NADC30-like PRRS virus challenge. Vet. Microbiol. 2020, 248, 108805. [Google Scholar] [CrossRef]
- Nakamoto, M.; Kunimura, K.; Suzuki, J.I.; Kodera, Y. Antimicrobial properties of hydrophobic compounds in garlic: Allicin, vinyldithiin, ajoene and diallyl polysulfides. Exp. Ther. Med. 2020, 19, 1550–1553. [Google Scholar] [CrossRef]
- Catanzaro, E.; Canistro, D.; Pellicioni, V.; Vivarelli, F.; Fimognari, C. Anticancer potential of allicin: A review. Pharmacol. Res. 2022, 177, 106118. [Google Scholar] [CrossRef]
- Li, C.; Lun, W.; Zhao, X.; Lei, S.; Guo, Y.; Ma, J.; Zhi, F. Allicin alleviates inflammation of trinitrobenzenesulfonic acid-induced rats and suppresses P38 and JNK pathways in Caco-2 cells. Mediat. Inflamm. 2015, 2015, 434692. [Google Scholar] [CrossRef]
- Rouf, R.; Uddin, S.J.; Sarker, D.K.; Islam, M.T.; Ali, E.S.; Shilpi, J.A.; Nahar, L.; Tiralongo, E.; Sarker, S.D. Antiviral potential of garlic (Allium sativum) and its organosulfur compounds: A systematic update of pre-clinical and clinical data. Trends Food Sci. Technol. 2020, 104, 219–234. [Google Scholar] [CrossRef]
- Borlinghaus, J.; Albrecht, F.; Gruhlke, M.C.; Nwachukwu, I.D.; Slusarenko, A.J. Allicin: Chemistry and biological properties. Molecules 2014, 19, 12591–12618. [Google Scholar] [CrossRef]
- Lawson, L.D.; Hunsaker, S.M. Allicin Bioavailability and Bioequivalence from Garlic Supplements and Garlic Foods. Nutrients 2018, 10, 812. [Google Scholar] [CrossRef]
- Weber, N.D.; Andersen, D.O.; North, J.A.; Murray, B.K.; Lawson, L.D.; Hughes, B.G. In vitro virucidal effects of Allium sativum (garlic) extract and compounds. Planta Med. 1992, 58, 417–423. [Google Scholar] [CrossRef]
- Mosbauer, K.; Fritsch, V.N.; Adrian, L.; Bernhardt, J.; Gruhlke, M.C.H.; Slusarenko, A.J.; Niemeyer, D.; Antelmann, H. The Effect of Allicin on the Proteome of SARS-CoV-2 Infected Calu-3 Cells. Front. Microbiol. 2021, 12, 746795. [Google Scholar] [CrossRef]
- Wang, L.; Jiao, H.; Zhao, J.; Wang, X.; Sun, S.; Lin, H. Allicin Alleviates Reticuloendotheliosis Virus-Induced Immunosuppression via ERK/Mitogen-Activated Protein Kinase Pathway in Specific Pathogen-Free Chickens. Front. Immunol. 2017, 8, 1856. [Google Scholar] [CrossRef]
- Che, H.Y.; Zhou, C.H.; Lyu, C.C.; Meng, Y.; He, Y.T.; Wang, H.Q.; Wu, H.Y.; Zhang, J.B.; Yuan, B. Allicin Alleviated LPS-Induced Mastitis via the TLR4/NF-kappaB Signaling Pathway in Bovine Mammary Epithelial Cells. Int. J. Mol. Sci. 2023, 24, 43805. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Che, T.M.; Song, M.; Lee, J.J.; Almeida, J.A.; Bravo, D.; Van Alstine, W.G.; Pettigrew, J.E. Dietary plant extracts improve immune responses and growth efficiency of pigs experimentally infected with porcine reproductive and respiratory syndrome virus. J. Anim. Sci. 2013, 91, 5668–5679. [Google Scholar] [CrossRef] [PubMed]
- Tannous, B.A. Gaussia luciferase reporter assay for monitoring biological processes in culture and in vivo. Nat. Protoc. 2009, 4, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Zhang, M.; Du, T.; Long, F.; Yang, X.; Sun, Y.; Duan, M.; Zhang, G.; Liu, Y.; Zhou, E.M.; Chen, W.; et al. Platycodin D Suppresses Type 2 Porcine Reproductive and Respiratory Syndrome Virus in Primary and Established Cell Lines. Viruses 2018, 10, 657. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, Q.; Chen, Y.; Duan, M.; Tian, G.; Deng, X.; Sun, Y.; Zhou, T.; Zhang, G.; Chen, W.; et al. Inhibition of proanthocyanidin A2 on porcine reproductive and respiratory syndrome virus replication in vitro. PLoS ONE 2018, 13, e0193309. [Google Scholar] [CrossRef]
- Wang, G.; Song, T.; Yu, Y.; Liu, Y.; Shi, W.; Wang, S.; Rong, F.; Dong, J.; Liu, H.; Cai, X.; et al. Immune responses in piglets infected with highly pathogenic porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 2011, 142, 170–178. [Google Scholar] [CrossRef]
- Shin, J.H.; Ryu, J.H.; Kang, M.J.; Hwang, C.R.; Han, J.; Kang, D. Short-term heating reduces the anti-inflammatory effects of fresh raw garlic extracts on the LPS-induced production of NO and pro-inflammatory cytokines by downregulating allicin activity in RAW 264.7 macrophages. Food Chem. Toxicol. 2013, 58, 545–551. [Google Scholar] [CrossRef]
- Gu, X.; Wu, H.; Fu, P. Allicin attenuates inflammation and suppresses HLA-B27 protein expression in ankylosing spondylitis mice. Biomed. Res. Int. 2013, 2013, 171573. [Google Scholar] [CrossRef]
- Ayaz, E.; Alpsoy, H.C. Garlic (Allium sativum) and traditional medicine. Turk. Parazitolojii Derg. 2007, 31, 145–149. [Google Scholar]
- Siegers, C.P.; Steffen, B.; Röbke, A.; Pentz, R. The effects of garlic preparations against human tumor cell proliferation. Phytomedicine Int. J. Phytother. Phytopharm. 1999, 6, 7–11. [Google Scholar] [CrossRef]
- Rahman, K.; Lowe, G.M. Garlic and cardiovascular disease: A critical review. J. Nutr. 2006, 136, 736s–740s. [Google Scholar] [CrossRef]
- El-Saber Batiha, G.; Magdy Beshbishy, A.; Wasef, G.L.; Elewa, Y.H.A.; Al-Sagan, A.A.-S.; Abd El-Hack, M.E.; Taha, A.E.; Abd-Elhakim, Y.M.; Prasad Devkota, H. Chemical Constituents and Pharmacological Activities of Garlic (Allium sativum L.): A Review. Nutrients 2020, 12, 872. [Google Scholar] [CrossRef]
- Block, E. The chemistry of garlic and onions. Sci. Am. 1985, 252, 114–119. [Google Scholar] [CrossRef]
- Melendez-Villanueva, M.A.; Moran-Santibanez, K.; Martinez-Sanmiguel, J.J.; Rangel-Lopez, R.; Garza-Navarro, M.A.; Rodriguez-Padilla, C.; Zarate-Trivino, D.G.; Trejo-Avila, L.M. Virucidal Activity of Gold Nanoparticles Synthesized by Green Chemistry Using Garlic Extract. Viruses 2019, 11, 1111. [Google Scholar] [CrossRef]
- Wei, X.; She, G.; Wu, T.; Xue, C.; Cao, Y. PEDV enters cells through clathrin-, caveolae-, and lipid raft-mediated endocytosis and traffics via the endo-/lysosome pathway. Vet. Res. 2020, 51, 10. [Google Scholar] [CrossRef]
- Benarroch, Y.; Juttukonda, L.; Sabharwal, V.; Boateng, J.; Khan, A.R.; Yarrington, C.; Wachman, E.M.; Taglauer, E. Differential Expression of Rab5 and Rab7 Small GTPase Proteins in Placental Tissues from Pregnancies Affected by Maternal Coronavirus Disease 2019. Clin. Ther. 2021, 43, 308–318. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Liu, Y.Y.; Xiao, F.C.; Liu, C.C.; Liang, X.D.; Chen, J.; Zhou, J.; Baloch, A.S.; Kan, L.; Zhou, B.; et al. Rab5, Rab7, and Rab11 Are Required for Caveola-Dependent Endocytosis of Classical Swine Fever Virus in Porcine Alveolar Macrophages. J. Virol. 2018, 92, e00797-18. [Google Scholar] [CrossRef]
- Shi, B.J.; Liu, C.C.; Zhou, J.; Wang, S.Q.; Gao, Z.C.; Zhang, X.M.; Zhou, B.; Chen, P.Y. Entry of Classical Swine Fever Virus into PK-15 Cells via a pH-, Dynamin-, and Cholesterol-Dependent, Clathrin-Mediated Endocytic Pathway That Requires Rab5 and Rab7. J. Virol. 2016, 90, 9194–9208. [Google Scholar] [CrossRef]
- Zhao, R.; Shi, Q.; Han, Z.; Fan, Z.; Ai, H.; Chen, L.; Li, L.; Liu, T.; Sun, J.; Liu, S. Newcastle Disease Virus Entry into Chicken Macrophages via a pH-Dependent, Dynamin and Caveola-Mediated Endocytic Pathway That Requires Rab5. J. Virol. 2021, 95, e0228820. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.Y.; Yang, Y.L.; Tang, Y.X.; Qin, P.; Wang, G.; Xie, J.Y.; Chen, S.X.; Ding, C.; Huang, Y.W.; Zhu, S.J. Bile acids promote the caveolae-associated entry of swine acute diarrhea syndrome coronavirus in porcine intestinal enteroids. PLoS Pathog. 2022, 18, e1010620. [Google Scholar] [CrossRef] [PubMed]
- Hu, J. The inhibitory effect of allicin on the expression of Rab5 and Rab7. College of Veterinary Medicine, Yangzhou University, Yangzhou, China. 2023; manuscript in preparation. [Google Scholar]
- Lu, Y.; He, Z.; Shen, X.; Xu, X.; Fan, J.; Wu, S.; Zhang, D. Cholesterol-lowering effect of allicin on hypercholesterolemic ICR mice. Oxid. Med. Cell Longev. 2012, 2012, 489690. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.H.; Lee, C. Cellular cholesterol is required for porcine nidovirus infection. Arch. Virol. 2017, 162, 3753–3767. [Google Scholar] [CrossRef]
- Gruhlke, M.C.H.; Antelmann, H.; Bernhardt, J.; Kloubert, V.; Rink, L.; Slusarenko, A.J. The human allicin-proteome: S-thioallylation of proteins by the garlic defence substance allicin and its biological effects. Free Radic. Biol. Med. 2019, 131, 144–153. [Google Scholar] [CrossRef]
- Bastikar, V.A.; Bastikar, A.V.; Chhajed, S.S. Understanding the Role of Natural Medicinal Compounds Such as Curcumin and Allicin against SARS-CoV-2 Proteins as Potential Treatment against COVID-19: An in silico Approach. J. Proteom. Bioinform. 2020, 13, 1–14. [Google Scholar]
- Shekh, S.; Reddy, K.K.A.; Gowd, K.H. In silico allicin induced S -thioallylation of SARS-CoV-2 main protease. J. Sulfur Chem. 2020, 42, 109–120. [Google Scholar] [CrossRef]
- Hu, J. The inhibitory effect of allicin on protease activity of PRRSV nsp4. College of Veterinary Medicine, Yangzhou University, Yangzhou, China. 2023; manuscript in preparation. [Google Scholar]
- Ganjhu, R.K.; Mudgal, P.P.; Maity, H.; Dowarha, D.; Devadiga, S.; Nag, S.; Arunkumar, G. Herbal plants and plant preparations as remedial approach for viral diseases. Virusdisease 2015, 26, 225–236. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Probe |
|---|---|---|
| PRRSV-ORF6 | CGGCAARTGATAACCACGC | GTCGTCCGGCGTCCCGG |
| TGCCACCCAACACGAGG | ||
| PRRSV-nsp9 | CGGCGGCTTAGTTGTTACTG | |
| TCAACCTCACTGGCCACTTT | ||
| IFN-β | CCTGAAGGCCAAGGAGTACA | CCACTCTGACTATGGTCCAGGCACA |
| AGCAATTGTCCAGTCCCAGA | ||
| IL-6 | CAGCCCTGAGAAAGGAGACA | TGCCAGTGCCTCTTTGCTGCT |
| CCAGGCAAGTCTCCTCATTG | ||
| TNFα | TCCCCAGGAAGACAGCG | |
| GCAGCAGACAGAAGAGCGT | ||
| IL8 | ACTCCAAACCTATCCACCCC | |
| CCACAACCCTAGACACCCAT | ||
| β-actin | CCCTGGAGAAGAGCTACGAG | CAGGAAGGAAGGTTGGAAGAG |
| CGGTTCCGCTGCCCTGAGGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Li, C.; Zhou, Y.; Ding, J.; Li, X.; Li, Y. Allicin Inhibits Porcine Reproductive and Respiratory Syndrome Virus Infection In Vitro and Alleviates Inflammatory Responses. Viruses 2023, 15, 1050. https://doi.org/10.3390/v15051050
Hu J, Li C, Zhou Y, Ding J, Li X, Li Y. Allicin Inhibits Porcine Reproductive and Respiratory Syndrome Virus Infection In Vitro and Alleviates Inflammatory Responses. Viruses. 2023; 15(5):1050. https://doi.org/10.3390/v15051050
Chicago/Turabian StyleHu, Jingbo, Chenxi Li, Yanyang Zhou, Jingjing Ding, Xiangdong Li, and Yanhua Li. 2023. "Allicin Inhibits Porcine Reproductive and Respiratory Syndrome Virus Infection In Vitro and Alleviates Inflammatory Responses" Viruses 15, no. 5: 1050. https://doi.org/10.3390/v15051050