
Characterization of a Bacteriophage GEC_vB_Bfr_UZM3 Active against Bacteroides fragilis
 , , ,
, , ,  , , , ,
, , , ,  and
and 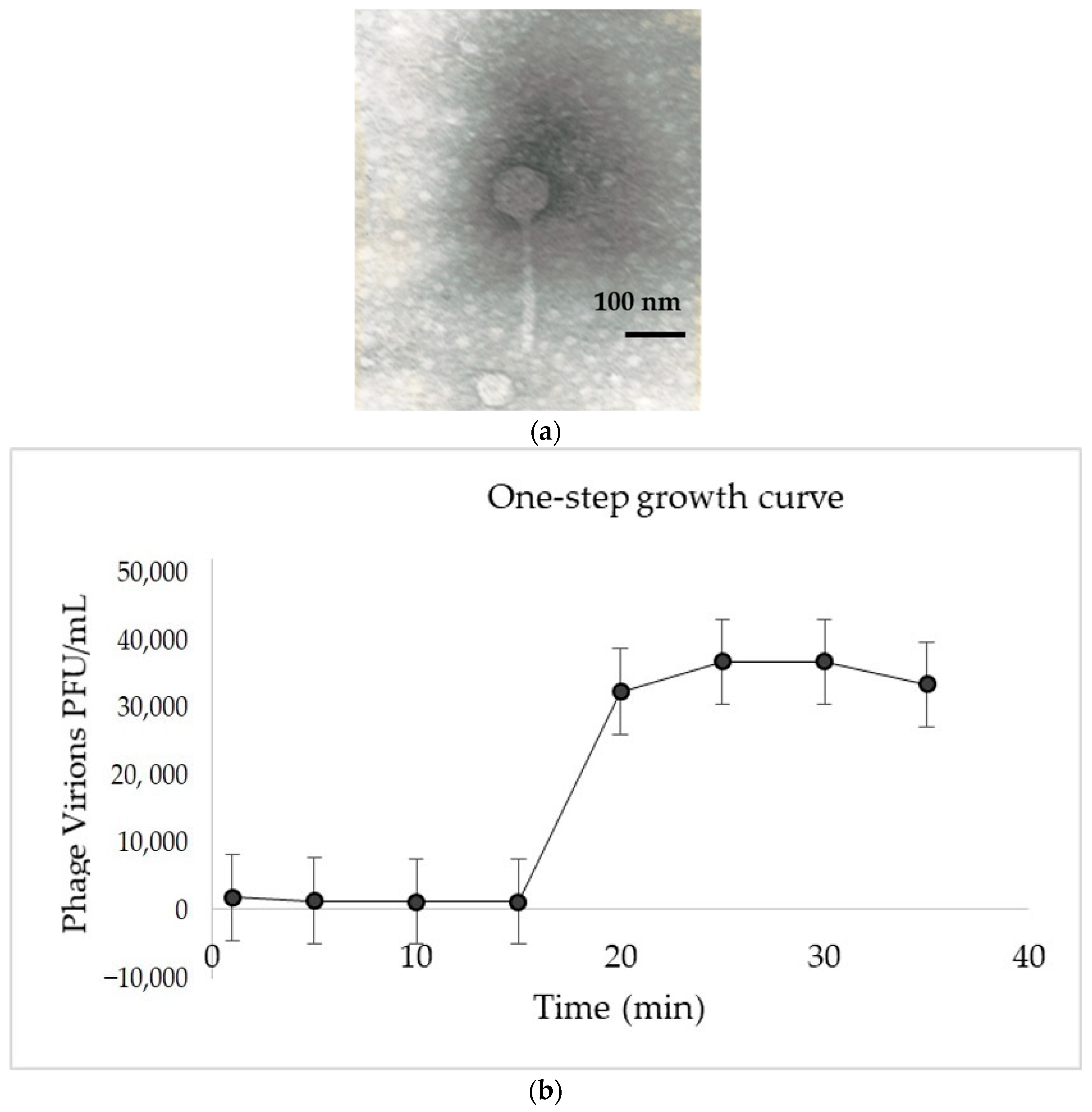{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Propagation of Bacteriophage
2.2. Morphological Characterization
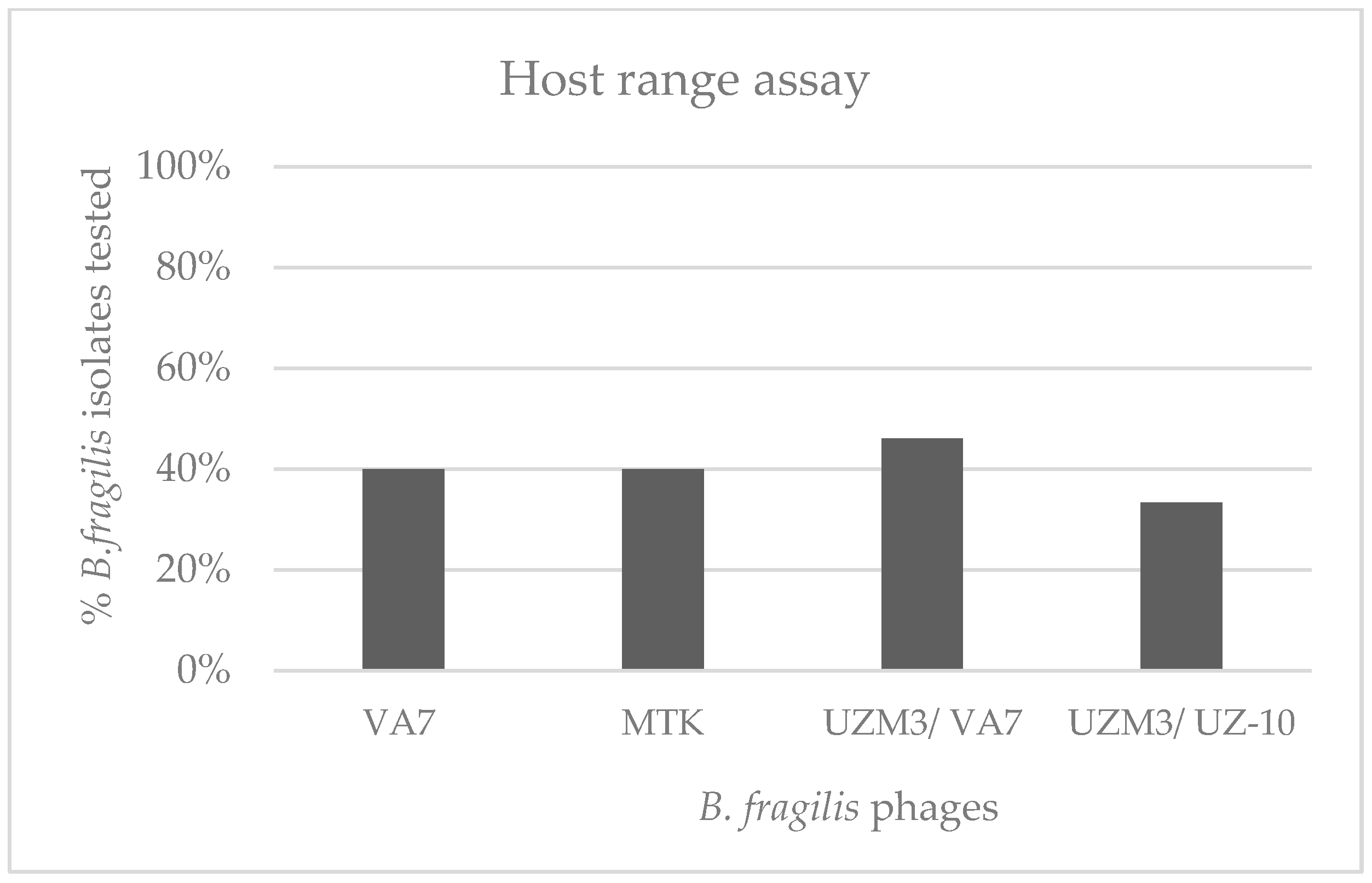
2.3. Host-Range Evaluation
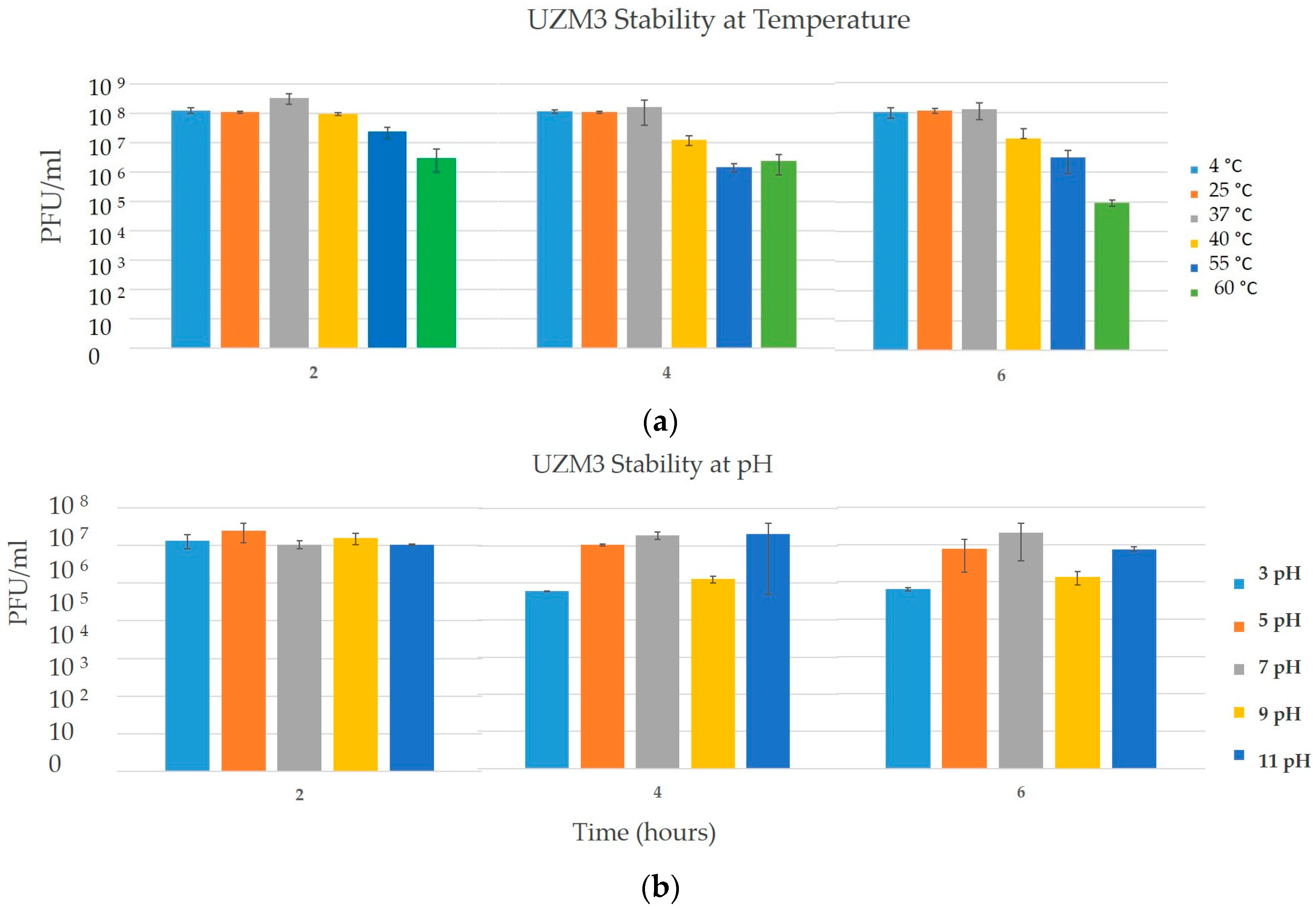
2.4. Temperature and pH Stability Assay
2.5. One-Step Growth Curve
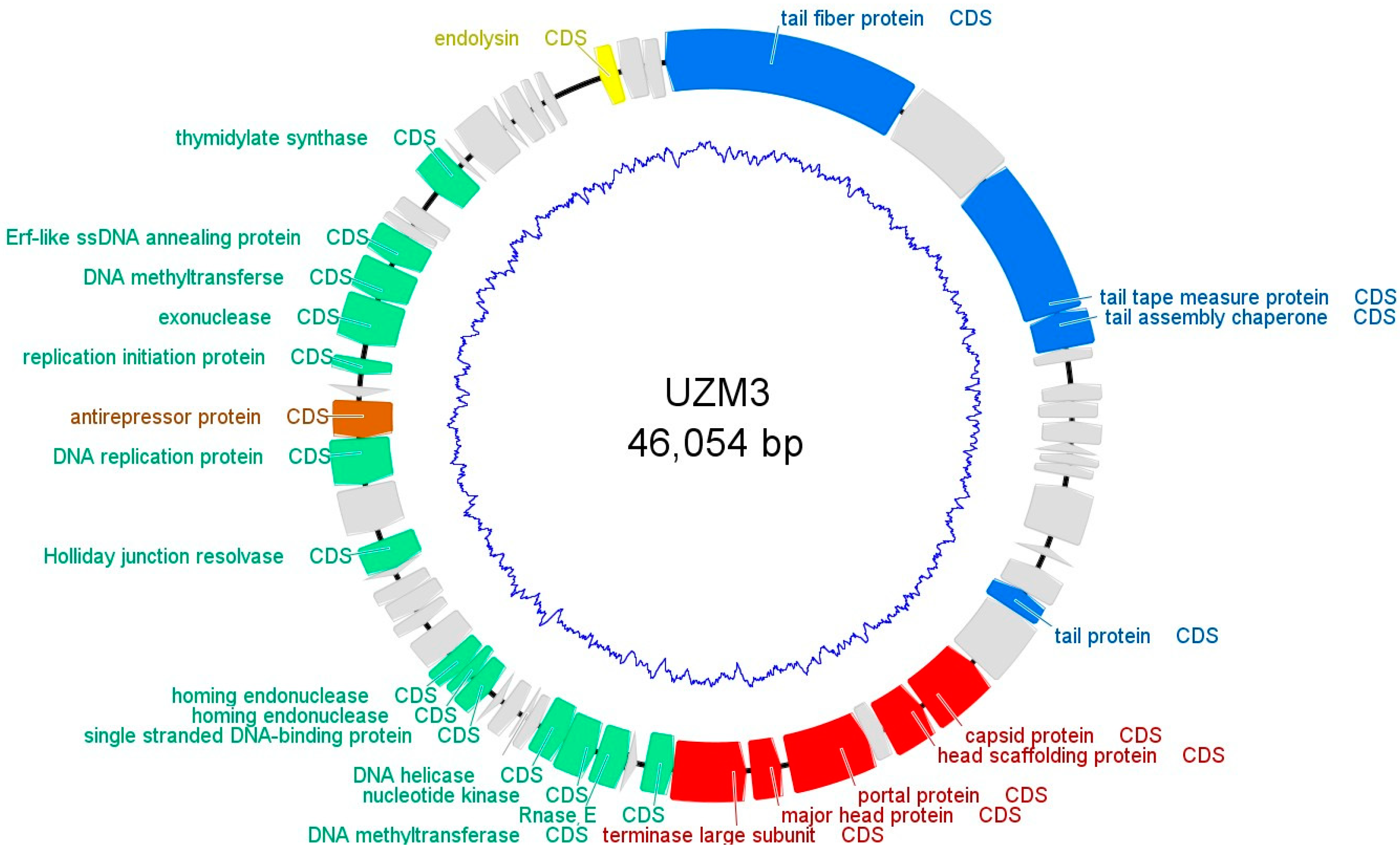
2.6. Whole Genome Sequencing and Analysis
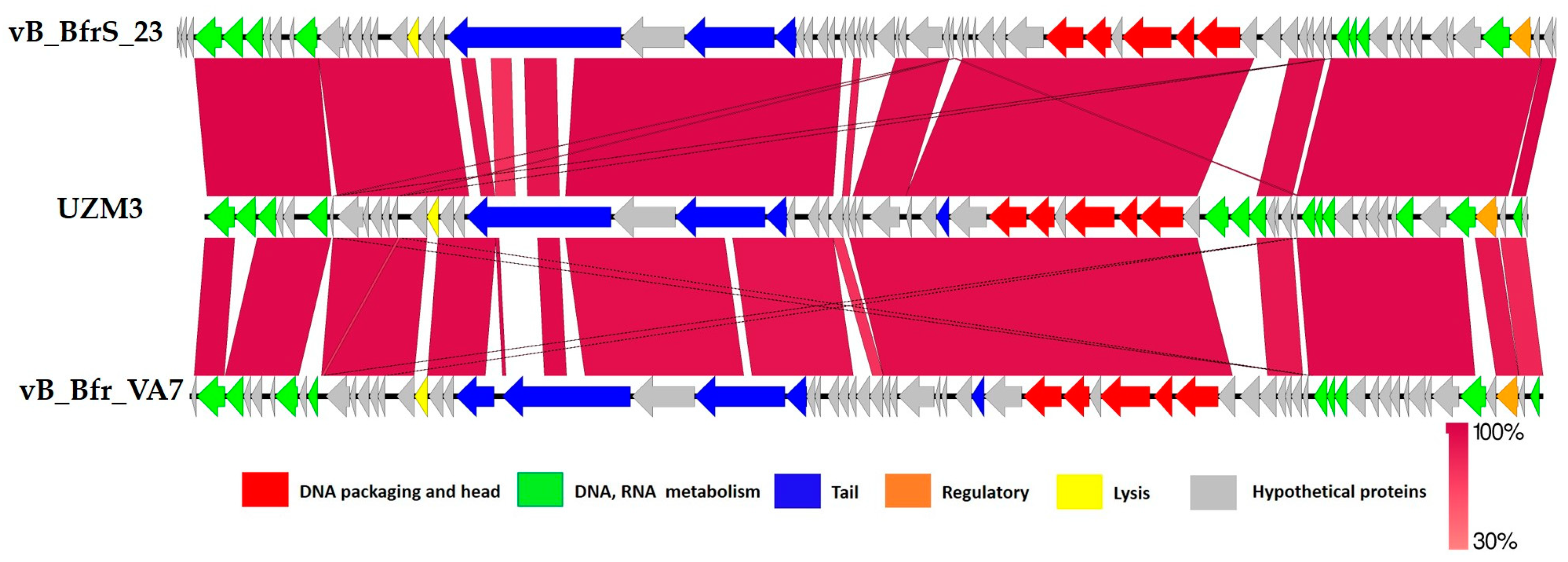
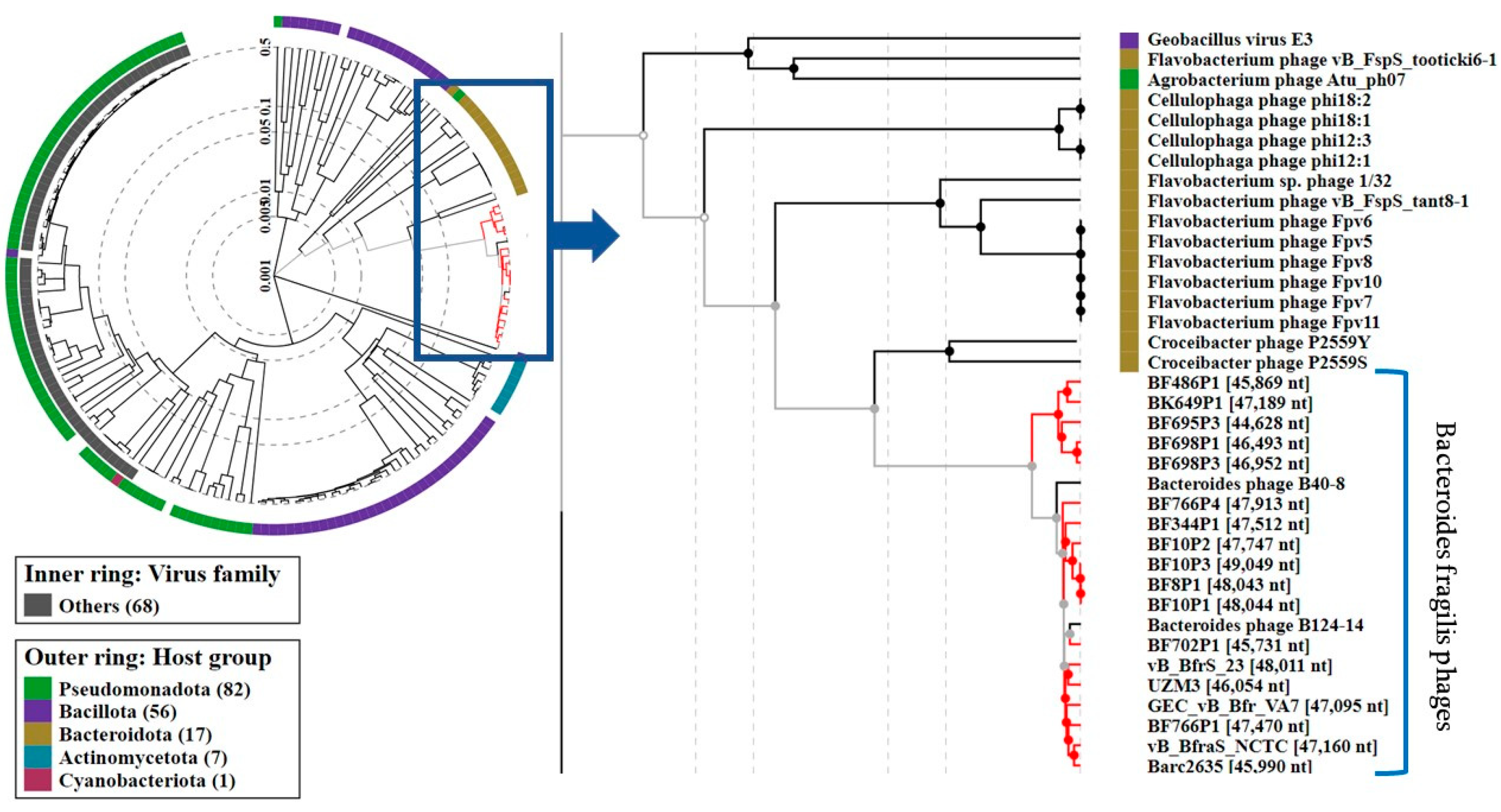
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Chanishvili, N. Phage therapy–history from Twort and d’Herelle through Soviet experience to current approaches. Adv. Virus Res. 2012, 83, 3–40. [Google Scholar] [CrossRef] [PubMed]
- Collaborators, A.R. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Pirnay, J.P.; Verbeken, G.; Ceyssens, P.J.; Huys, I.; De Vos, D.; Ameloot, C.; Fauconnier, A. The Magistral Phage. Viruses 2018, 10, 64. [Google Scholar] [CrossRef] [PubMed]
- Wexler, H.M. Bacteroides: The good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef]
- Yekani, M.; Rezaee, M.A.; Beheshtirouy, S.; Baghi, H.B.; Bazmani, A.; Farzinazar, A.; Memar, M.Y.; Sóki, J. Carbapenem resistance in Bacteroides fragilis: A review of molecular mechanisms. Anaerobe 2022, 76, 102606. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, J.E.; Fraser, S.; Citron, D.M.; Wexler, H.; Blakely, G.; Jobling, K.; Patrick, S. Multi-drug resistant Bacteroides fragilis recovered from blood and severe leg wounds caused by an improvised explosive device (IED) in Afghanistan. Anaerobe 2011, 17, 152–155. [Google Scholar] [CrossRef]
- Kalapila, A.; Pergam, S.; Pottinger, P.; Butler-Wu, S.; Whimbey, E.; Duchin, J. Multidrug-resistant bacteroides fragilis–Seattle, Washington, 2013. MMWR Morb. Mortal. Wkly. Rep. 2013, 62, 694–696. [Google Scholar]
- Merchan, C.; Parajuli, S.; Siegfried, J.; Scipione, M.R.; Dubrovskaya, Y.; Rahimian, J. Multidrug-Resistant Bacteroides fragilis Bacteremia in a US Resident: An Emerging Challenge. Case Rep. Infect. Dis. 2016, 2016, 3607125. [Google Scholar] [CrossRef] [PubMed]
- Hartmeyer, G.N.; Sóki, J.; Nagy, E.; Justesen, U.S. Multidrug-resistant Bacteroides fragilis group on the rise in Europe? J. Med. Microbiol. 2012, 61 Pt 12, 1784–1788. [Google Scholar] [CrossRef]
- Urbán, E.; Horváth, Z.; Sóki, J.; Lázár, G. First Hungarian case of an infection caused by multidrug-resistant Bacteroides fragilis strain. Anaerobe 2015, 31, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Wu, S.; Xu, T.; Zhao, X.; Huang, H.; Hu, F. Emergence of carbapenem resistance in Bacteroides fragilis in China. Int. J. Antimicrob. Agents 2019, 53, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, I.; Aoki, K.; Miura, Y.; Yamaguchi, T.; Matsumoto, T. Fatal sepsis caused by multidrug-resistant Bacteroides fragilis, harboring a cfiA gene and an upstream insertion sequence element, in Japan. Anaerobe 2017, 44, 36–39. [Google Scholar] [CrossRef]
- Scotto d’Abusco, A.S.; Sanfilippo, L.; Menozzi, M.G.; Pantosti, A. Activity and role of BFT, an enterotoxin produced by Bacteroides fragilis. J. Nat. Toxins 2000, 9, 267–280. [Google Scholar]
- Xie, X.; Jiang, D.; Zhou, X.; Ye, X.; Yang, P.; He, Y. Recombinant. Open Life Sci. 2021, 16, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Sears, C.L. Enterotoxigenic Bacteroides fragilis: A rogue among symbiotes. Clin. Microbiol. Rev. 2009, 22, 349–369. [Google Scholar] [CrossRef] [PubMed]
- Wick, E.C.; Sears, C.L. Bacteroides spp. and diarrhea. Curr. Opin. Infect. Dis. 2010, 23, 470–474. [Google Scholar] [CrossRef]
- Cohen, S.H.; Shetab, R.; Tang-Feldman, Y.J.; Sarma, P.; Silva, J.; Prindiville, T.P. Prevalence of enterotoxigenic Bacteroides fragilis in hospital-acquired diarrhea. Diagn. Microbiol. Infect. Dis. 2006, 55, 251–254. [Google Scholar] [CrossRef]
- Queralt, N.; Jofre, J.; Araujo, R.; Muniesa, M. Homogeneity of the morphological groups of bacteriophages infecting Bacteroides fragilis strain HSP40 and strain RYC2056. Curr. Microbiol. 2003, 46, 163–168. [Google Scholar] [CrossRef]
- Tariq, M.A.; Newberry, F.; Haagmans, R.; Booth, C.; Wileman, T.; Hoyles, L.; Clokie, M.R.J.; Ebdon, J.; Carding, S.R. Genome Characterization of a Novel Wastewater. Front. Microbiol. 2020, 11, 583378. [Google Scholar] [CrossRef]
- Van Belleghem, J.D.; Clement, F.; Merabishvili, M.; Lavigne, R.; Vaneechoutte, M. Pro- and anti-inflammatory responses of peripheral blood mononuclear cells induced by Staphylococcus aureus and Pseudomonas aeruginosa phages. Sci. Rep. 2017, 7, 8004. [Google Scholar] [CrossRef] [PubMed]
- Friman, V.P.; Ghoul, M.; Molin, S.; Johansen, H.K.; Buckling, A. Pseudomonas aeruginosa adaptation to lungs of cystic fibrosis patients leads to lowered resistance to phage and protist enemies. PLoS ONE 2013, 8, e75380. [Google Scholar] [CrossRef] [PubMed]
- Onsea, J.; Soentjens, P.; Djebara, S.; Merabishvili, M.; Depypere, M.; Spriet, I.; De Munter, P.; Debaveye, Y.; Nijs, S.; Vanderschot, P.; et al. Bacteriophage Application for Difficult-to-treat Musculoskeletal Infections: Development of a Standardized Multidisciplinary Treatment Protocol. Viruses 2019, 11, 891. [Google Scholar] [CrossRef] [PubMed]
- Bakuradze, N.; Makalatia, K.; Merabishvili, M.; Togoshvili, L.; Chanishvili, N. Selection of the active phages against B. fragilis for further study of thrapeutic perpectives. Georgian Med. News 2018, 285, 111–116. [Google Scholar]
- Bakuradze, N.; Merabishvili, M.; Makalatia, K.; Kakabadze, E.; Grdzelishvili, N.; Wagemans, J.; Lood, C.; Chachua, I.; Vaneechoutte, M.; Lavigne, R.; et al. In Vitro Evaluation of the Therapeutic Potential of Phage VA7 against Enterotoxigenic B. fragilis. Viruses 2021, 13, 2044. [Google Scholar] [CrossRef]
- Issa, R.; Chanishvili, N.; Caplin, J.; Kakabadze, E.; Bakuradze, N.; Makalatia, K.; Cooper, I. Antibiofilm potential of purified environmental bacteriophage preparations against early stage Pseudomonas aeruginosa biofilms. J. Appl. Microbiol. 2019, 126, 1657–1667. [Google Scholar] [CrossRef]
- Fanaei Pirlar, R.; Wagemans, J.; Ponce Benavente, L.; Lavigne, R.; Trampuz, A.; Gonzalez Moreno, M. Novel Bacteriophage Specific against. Viruses 2022, 14, 1340. [Google Scholar] [CrossRef]
- Kropinski, A.M. Practical Advice on the One-Step Growth Curve. Methods Mol. Biol. 2018, 1681, 41–47. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Carver, T.; Harris, S.R.; Berriman, M.; Parkhill, J.; McQuillan, J.A. Artemis: An integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics 2012, 28, 464–469. [Google Scholar] [CrossRef]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef]
- Terzian, P.; Olo Ndela, E.; Galiez, C.; Lossouarn, J.; Pérez Bucio, R.E.; Mom, R.; Toussaint, A.; Petit, M.A.; Enault, F. PHROG: Families of prokaryotic virus proteins clustered using remote homology. NAR Genom. Bioinform. 2021, 3, lqab067. [Google Scholar] [CrossRef] [PubMed]
- Gabler, F.; Nam, S.Z.; Till, S.; Mirdita, M.; Steinegger, M.; Söding, J.; Lupas, A.N.; Alva, V. Protein Sequence Analysis Using the MPI Bioinformatics Toolkit. Curr. Protoc. Bioinform. 2020, 72, e108. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Rohwer, F.; Edwards, R. The Phage Proteomic Tree: A genome-based taxonomy for phage. J. Bacteriol. 2002, 184, 4529–4535. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Moraru, C.; Varsani, A.; Kropinski, A.M. VIRIDIC-A Novel Tool to Calculate the Intergenomic Similarities of Prokaryote-Infecting Viruses. Viruses 2020, 12, 1268. [Google Scholar] [CrossRef]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA Genes in Genomic Sequences. Methods Mol. Biol. 2019, 1962, 1–14. [Google Scholar] [CrossRef]
- Adriaenssens, E.; Brister, J.R. How to Name and Classify Your Phage: An Informal Guide. Viruses 2017, 9, 70. [Google Scholar] [CrossRef]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and Use of Personalized Bacteriophage-Based Therapeutic Cocktails To Treat a Patient with a Disseminated Resistant Acinetobacter baumannii Infection. Antimicrob. Agents Chemother. 2017, 61, e00954-17. [Google Scholar] [CrossRef]
- Sulakvelidze, A. Phage therapy: An attractive option for dealing with antibiotic-resistant bacterial infections. Drug Discov. Today 2005, 10, 807–809. [Google Scholar] [CrossRef]
- Jasemi, S.; Emaneini, M.; Ahmadinejad, Z.; Fazeli, M.S.; Sechi, L.A.; Sadeghpour Heravi, F.; Feizabadi, M.M. Antibiotic resistance pattern of Bacteroides fragilis isolated from clinical and colorectal specimens. Ann. Clin. Microbiol. Antimicrob. 2021, 20, 27. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, H.; Edberg, S.; Pierzo, V.; Delattre, J.M. Bacteriophages as indicators of enteric viruses and public health risk in groundwaters. J. Appl. Microbiol. 2000, 88, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Jofre, J.; Blanch, A.R.; Lucena, F.; Muniesa, M. Bacteriophages infecting Bacteroides as a marker for microbial source tracking. Water Res. 2014, 55, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.; Sillankorva, S.; Merabishvili, M.; Kluskens, L.D.; Azeredo, J. Unexploited opportunities for phage therapy. Front. Pharm. 2015, 6, 180. [Google Scholar] [CrossRef] [PubMed]
- Międzybrodzki, R.; Kłak, M.; Jończyk-Matysiak, E.; Bubak, B.; Wójcik, A.; Kaszowska, M.; Weber-Dąbrowska, B.; Łobocka, M.; Górski, A. Means to Facilitate the Overcoming of Gastric Juice Barrier by a Therapeutic Staphylococcal Bacteriophage A5/80. Front. Microbiol. 2017, 8, 467. [Google Scholar] [CrossRef]
- Abdulamir, A.S.; Jassim, S.A.; Abu Bakar, F. Novel approach of using a cocktail of designed bacteriophages against gut pathogenic E. coli for bacterial load biocontrol. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 39. [Google Scholar] [CrossRef]
- Jończyk-Matysiak, E.; Łodej, N.; Kula, D.; Owczarek, B.; Orwat, F.; Międzybrodzki, R.; Neuberg, J.; Bagińska, N.; Weber-Dąbrowska, B.; Górski, A. Factors determining phage stability/activity: Challenges in practical phage application. Expert Rev. Anti Infect. Ther. 2019, 17, 583–606. [Google Scholar] [CrossRef]
- Egido, J.E.; Costa, A.R.; Aparicio-Maldonado, C.; Haas, P.J.; Brouns, S.J.J. Mechanisms and clinical importance of bacteriophage resistance. FEMS Microbiol. Rev. 2022, 46, fuab048. [Google Scholar] [CrossRef]
- Kossykh, V.G.; Schlagman, S.L.; Hattman, S. Phage T4 DNA [N6-adenine] methyltransferase. Overexpression, purification, and characterization. J. Biol. Chem. 1995, 270, 14389–14393. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakuradze, N.; Merabishvili, M.; Kusradze, I.; Ceyssens, P.-J.; Onsea, J.; Metsemakers, W.-J.; Grdzelishvili, N.; Natroshvili, G.; Tatrishvili, T.; Lazvliashvili, D.; et al. Characterization of a Bacteriophage GEC_vB_Bfr_UZM3 Active against Bacteroides fragilis. Viruses 2023, 15, 1042. https://doi.org/10.3390/v15051042
Bakuradze N, Merabishvili M, Kusradze I, Ceyssens P-J, Onsea J, Metsemakers W-J, Grdzelishvili N, Natroshvili G, Tatrishvili T, Lazvliashvili D, et al. Characterization of a Bacteriophage GEC_vB_Bfr_UZM3 Active against Bacteroides fragilis. Viruses. 2023; 15(5):1042. https://doi.org/10.3390/v15051042
Chicago/Turabian StyleBakuradze, Nata, Maia Merabishvili, Ia Kusradze, Pieter-Jan Ceyssens, Jolien Onsea, Willem-Jan Metsemakers, Nino Grdzelishvili, Guliko Natroshvili, Tamar Tatrishvili, Davit Lazvliashvili, and et al. 2023. "Characterization of a Bacteriophage GEC_vB_Bfr_UZM3 Active against Bacteroides fragilis" Viruses 15, no. 5: 1042. https://doi.org/10.3390/v15051042