
The Establishment and Application of Indirect 3AB-ELISA for the Detection of Antibodies against Senecavirus A
Abstract
:1. Introduction
2. Materials and Methods
2.1. Serum Samples
2.2. Expression of Recombinant Protein
2.3. Purification of Recombinant Protein
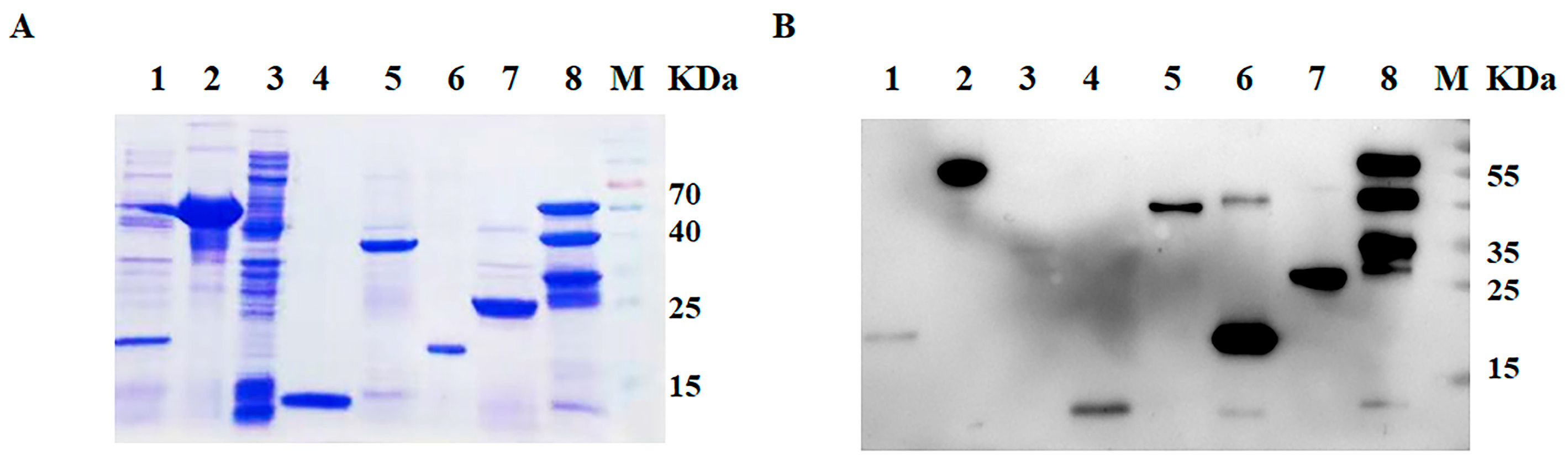
2.4. SDS-PAGE and Western Blotting
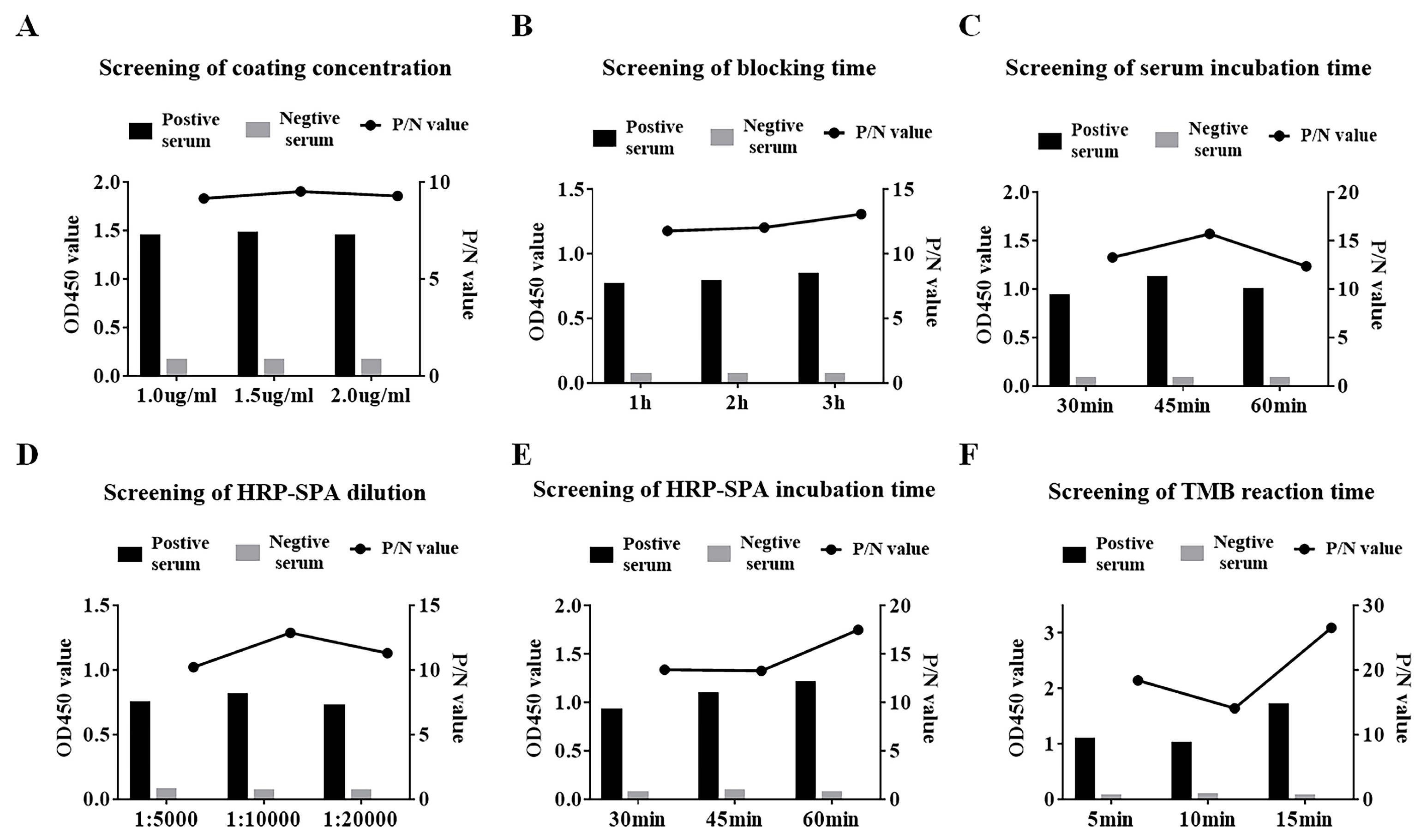
2.5. ELISA Procedure
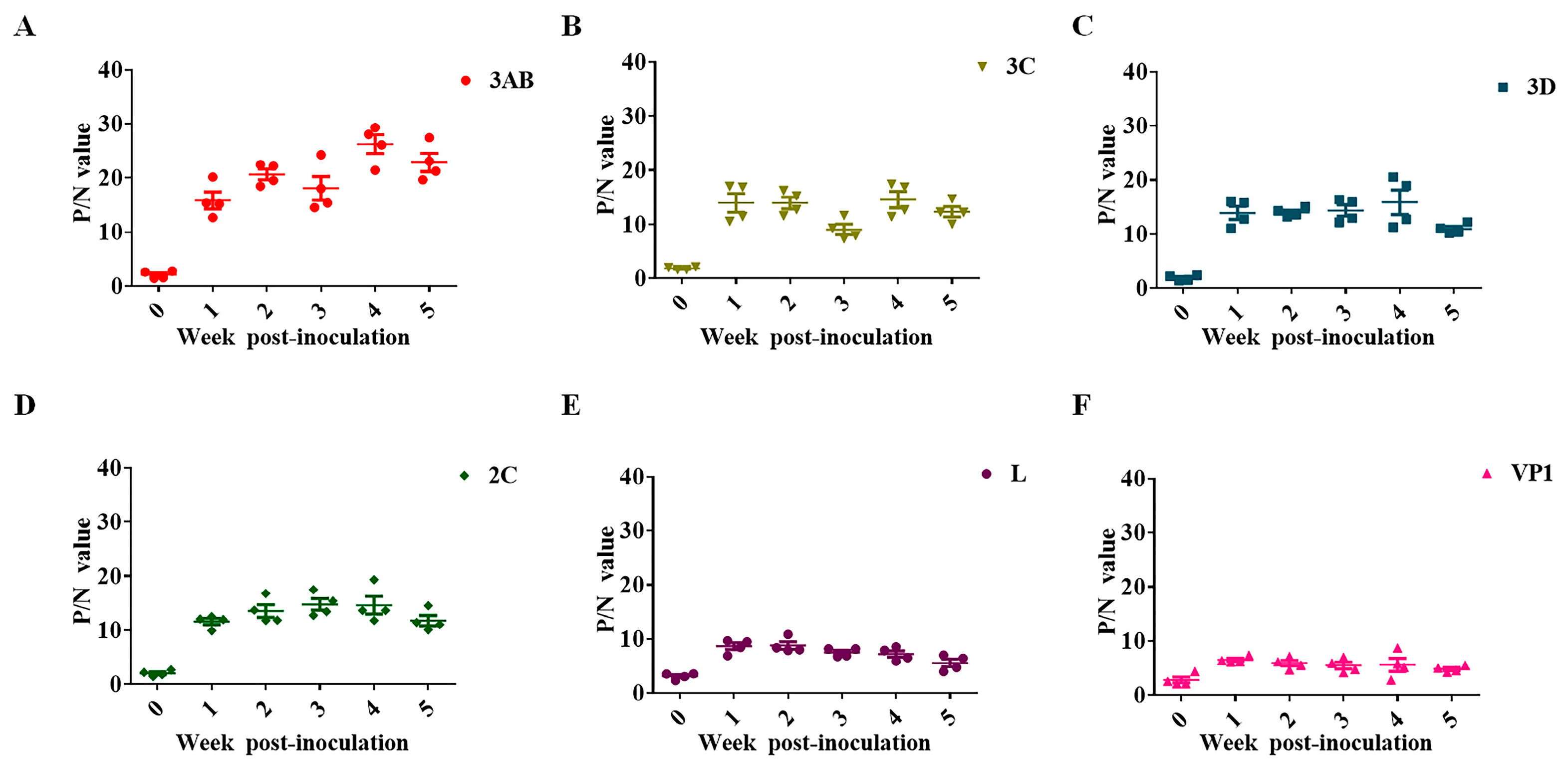
2.6. Antibody Kinetics Analysis
2.7. Virus Neutralizing Test (VNT)
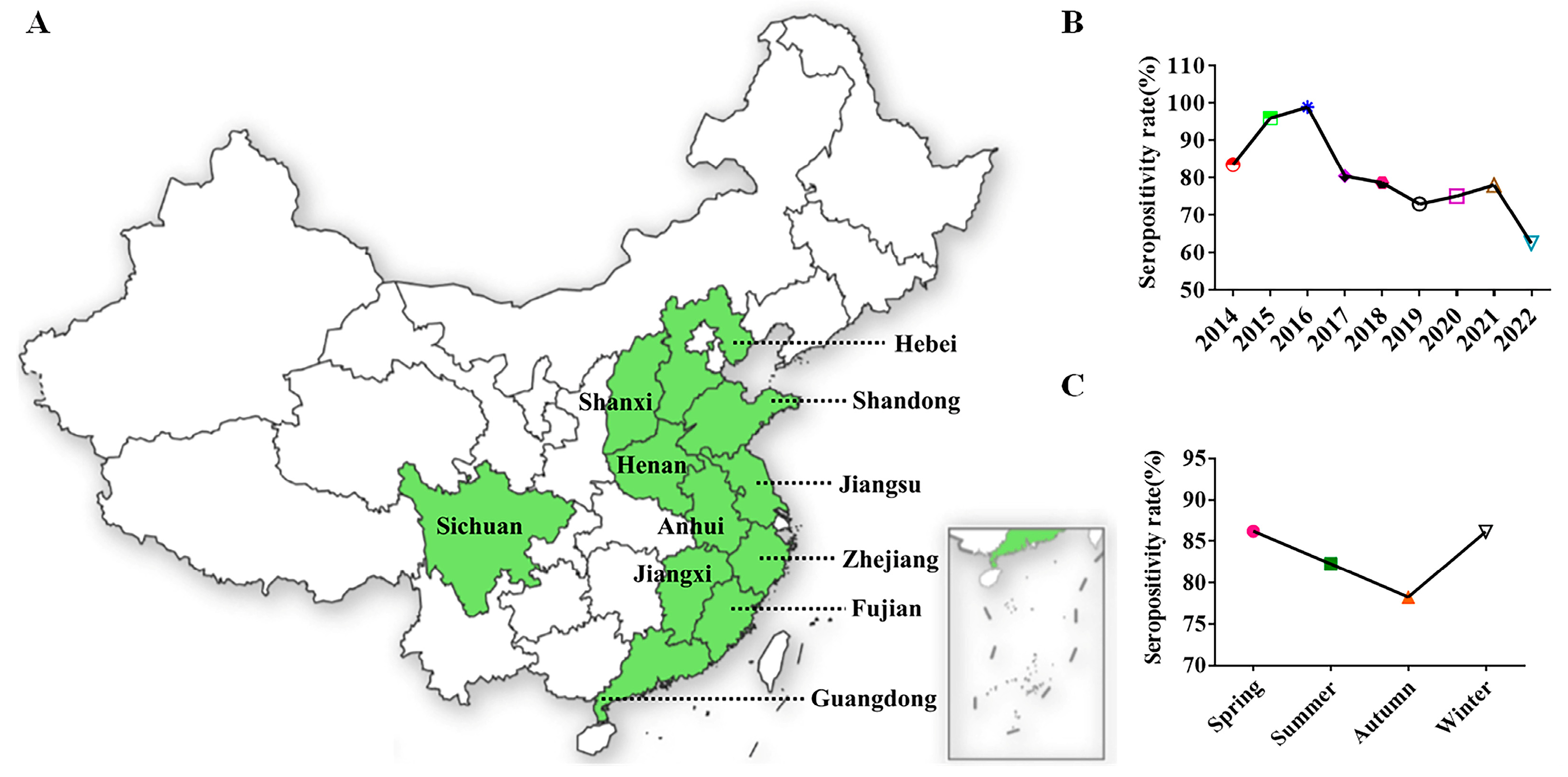
2.8. Retrospective Serological Survey by 3AB ELISA
3. Results
3.1. Purification and Identification of SVA Protein
3.2. Antibody Kinetic Analysis
3.3. Indirect ELISA for SVA 3AB
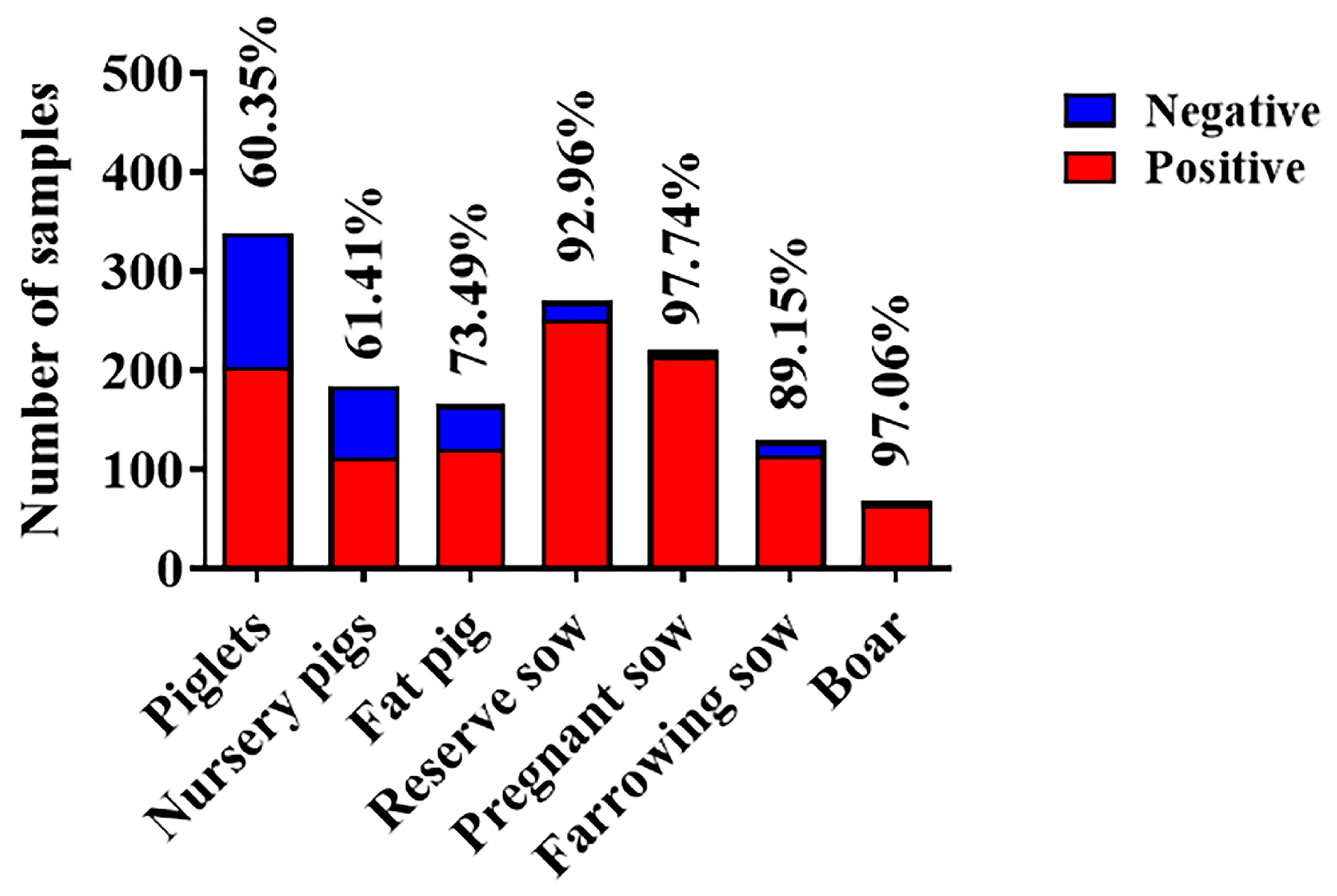
3.4. Retrospective Serological Survey
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hales, L.M.; Knowles, N.J.; Reddy, P.S.; Xu, L.; Hay, C.; Hallenbeck, P.L.Y. Complete Genome Sequence Analysis of Seneca Valley Virus-001, a Novel Oncolytic Picornavirus. J. Gen. Virol. 2008, 89, 1265–1275. [Google Scholar] [CrossRef]
- Pasma, T.; Davidson, S.; Shaw, S.L. Idiopathic Vesicular Disease in Swine in Manitoba. Can. Vet. J. Rev. Vet. Can. 2008, 49, 84–85. [Google Scholar]
- Leme, R.A.; Zotti, E.; Alcântara, B.K.; Oliveira, M.V.; Freitas, L.A.; Alfieri, A.F.; Alfieri, A.A. Senecavirus A: An Emerging Vesicular Infection in Brazilian Pig Herds. Transbound. Emerg. Dis. 2015, 62, 603–611. [Google Scholar] [CrossRef]
- Guo, B.; Piñeyro, P.E.; Rademacher, C.J.; Zheng, Y.; Li, G.; Yuan, J.; Hoang, H.; Gauger, P.C.; Madson, D.M.; Schwartz, K.J.; et al. Novel Senecavirus A in Swine with Vesicular Disease, United States, July. Emerg. Infect. Dis. 2016, 22, 1325–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Vannucci, F.; Knutson, T.P.; Corzo, C.; Marthaler, D.G. Emergence and Whole-Genome Sequence of Senecavirus A in Colombia. Transbound. Emerg. Dis. 2017, 64, 1346–1349. [Google Scholar] [CrossRef] [PubMed]
- Saeng-Chuto, K.; Rodtian, P.; Temeeyasen, G.; Wegner, M.; Nilubol, D. The First Detection of Senecavirus A in Pigs in Thailand. Transbound. Emerg. Dis. 2018, 65, 285–288. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, X.; Bai, Y.; Sun, B.; Xie, Q.; Ma, J. The First Identification and Complete Genome of Senecavirus A Affecting Pig with Idiopathic Vesicular Disease in China. Transbound. Emerg. Dis. 2017, 64, 1633–1640. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Fan, H.; Zhou, E.; Li, L.; Li, S.; Yan, J.; Jiang, P. Pathogenesis of a Senecavirus a Isolate from Swine in Shandong Province, China. Vet. Microbiol. 2020, 242, 108606. [Google Scholar] [CrossRef]
- Segalés, J.; Barcellos, D.; Alfieri, A.; Burrough, E.; Marthaler, D. Senecavirus A. Vet. Pathol. 2017, 54, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Bachrach, H.L.; Moore, D.M.; McKercher, P.D.; Polatnick, J. Immune and Antibody Responses to an Isolated Capsid Protein of Foot-and-Mouth Disease Virus. J. Immunol. Baltim. Md 1950 1975, 115, 1636–1641. [Google Scholar] [CrossRef]
- Ludi, A.B.; Morris, A.; Gubbins, S.; Asfor, A.; Afzal, M.; Browning, C.F.; Grazioli, S.; Foglia, E.A.; Wilsden, G.; Burman, A.; et al. Cross-Serotype Reactivity of ELISAs Used to Detect Antibodies to the Structural Proteins of Foot-and-Mouth Disease Virus. Viruses 2022, 14, 1495. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Wang, H.; Wei, H.; Yan, Y.; Zhao, T.; Hu, X.; Luo, P.; Wang, L.; Yu, Y. A Recombinant Truncated FMDV 3AB Protein Used to Better Distinguish between Infected and Vaccinated Cattle. Vaccine 2010, 28, 3435–3439. [Google Scholar] [CrossRef] [PubMed]
- Leme, R.A.; Alfieri, A.F.; Alfieri, A.A. Update on Senecavirus Infection in Pigs. Viruses 2017, 9, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Z.; Li, Z.; Yan, Q.; Li, Y.; Xiong, W.; Wu, K.; Li, X.; Fan, S.; Zhao, M.; Ding, H.; et al. Development of Diagnostic Tests Provides Technical Support for the Control of African Swine Fever. Vaccines 2021, 9, 343. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, J.; Chen, H.; Zhou, J.; Ding, Y.; Liu, Y. An Overview on ELISA Techniques for FMD. Virol. J. 2011, 8, 419. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; van Bruggen, R.; Xu, W. Generation and Diagnostic Application of Monoclonal Antibodies against Seneca Valley Virus. J. Vet. Diagn. Investig. 2012, 24, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Gimenez-Lirola, L.G.; Rademacher, C.; Linhares, D.; Harmon, K.; Rotolo, M.; Sun, Y.; Baum, D.H.; Zimmerman, J.; Piñeyro, P. Serological and Molecular Detection of Senecavirus A Associated with an Outbreak of Swine Idiopathic Vesicular Disease and Neonatal Mortality. J. Clin. Microbiol. 2016, 54, 2082–2089. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Lv, J.; Zhang, Z.; Pan, L. Development of an Indirect ELISA Using a Novel Linear Epitope at the C-Terminal Region of the VP2 Protein to Specifically Detect Antibodies against Senecavirus A. Virol. J. 2022, 19, 204. [Google Scholar] [CrossRef]
- Gao, Y.; Xia, T.; Bai, J.; Zhang, L.; Zheng, H.; Jiang, P. Preparation of Monoclonal Antibodies against the Viral P54 Protein and a Blocking ELISA for Detection of the Antibody against African Swine Fever Virus. Viruses 2022, 14, 2335. [Google Scholar] [CrossRef]
- Tesfagaber, W.; Wang, L.; Tsegay, G.; Hagoss, Y.T.; Zhang, Z.; Zhang, J.; Huangfu, H.; Xi, F.; Li, F.; Sun, E.; et al. Characterization of Anti-P54 Monoclonal Antibodies and Their Potential Use for African Swine Fever Virus Diagnosis. Pathogens 2021, 10, 178. [Google Scholar] [CrossRef]
- Budiman, C.; Bando, K.; Angkawidjaja, C.; Koga, Y.; Takano, K.; Kanaya, S. Engineering of Monomeric FK506-Binding Protein 22 with Peptidyl Prolyl Cis-Trans Isomerase. Importance of a V-Shaped Dimeric Structure for Binding to Protein Substrate. FEBS J. 2009, 276, 4091–4101. [Google Scholar] [CrossRef]
- Budiman, C.; Razak, R.A.; Unggit, A.R.A.; Razali, R.; Suzery, M.; Mokhtar, R.A.M.; Lee, P.-C.; Utomo, D.H. Catalytic Properties of Caseinolytic Protease Subunit of Plasmodium Knowlesi and Its Inhibition by a Member of δ-Lactone, Hyptolide. Molecules 2022, 27, 3787. [Google Scholar] [CrossRef]
- Gabe, C.M.; Brookes, S.J.; Kirkham, J. Preparative SDS PAGE as an Alternative to His-Tag Purification of Recombinant Amelogenin. Front. Physiol. 2017, 8, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Liu, L.; Du, G.; Liu, S. A Multifunctional Tag with the Ability to Benefit the Expression, Purification, Thermostability and Activity of Recombinant Proteins. J. Biotechnol. 2018, 283, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Burgess, R.R. Chapter 17 Refolding Solubilized Inclusion Body Proteins. In Methods in Enzymology; Burgess, R.R., Deutscher, M.P., Eds.; Guide to Protein Purification, 2nd ed.; Academic Press: Cambridge, MA, USA, 2009; Volume 463, pp. 259–282. [Google Scholar]
- Padhiar, A.A.; Chanda, W.; Joseph, T.P.; Guo, X.; Liu, M.; Sha, L.; Batool, S.; Gao, Y.; Zhang, W.; Huang, M.; et al. Comparative Study to Develop a Single Method for Retrieving Wide Class of Recombinant Proteins from Classical Inclusion Bodies. Appl. Microbiol. Biotechnol. 2018, 102, 2363–2377. [Google Scholar] [CrossRef]
- Fan, H.; Zhu, H.; Li, S.; Shi, M.; Zhou, E.; Wang, X.; Jiang, P.; Bai, J. Identification of Linear B Cell Epitopes on VP1 and VP2 Proteins of Senecavirus A (SVA) Using Monoclonal Antibodies. Vet. Microbiol. 2020, 247, 108753. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Xie, Y.; Guan, Z.; Zhang, Y.; Li, Y.; Yang, Y.; Zhang, J.; Li, Z.; Qiu, Y.; Li, B.; et al. Seroprevalence of Getah Virus in Pigs in Eastern China Determined with a Recombinant E2 Protein-Based Indirect ELISA. Viruses 2022, 14, 2173. [Google Scholar] [CrossRef] [PubMed]
- Abade Dos Santos, F.A.; Santos, N.; Carvalho, C.L.; Martinez-Haro, M.; Gortázar, C.; García-Bocanegra, I.; Capucci, L.; Duarte, M.; Alves, P.C. Retrospective Serological and Molecular Survey of Myxoma or Antigenically Related Virus in the Iberian Hare, Lepus Granatensis. Transbound. Emerg. Dis. 2022, 69, 3637–3650. [Google Scholar] [CrossRef]
- Silveira, S.; Falkenberg, S.M.; Elderbrook, M.J.; Sondgeroth, K.S.; Dassanayake, R.P.; Neill, J.D.; Ridpath, J.F.; Canal, C.W. Serological Survey for Antibodies against Pestiviruses in Wyoming Domestic Sheep. Vet. Microbiol. 2018, 219, 96–99. [Google Scholar] [CrossRef]
- Yi, W.; Zhu, H.; Wu, Y.; Li, Q.; Lou, W.; Zhao, H.; Pan, Z. The Recombinant Erns and Truncated E2-Based Indirect Enzyme-Linked Immunosorbent Assays to Distinguishably Test Specific Antibodies against Classical Swine Fever Virus and Bovine Viral Diarrhea Virus. Virol. J. 2022, 19, 121. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, Z.; Yang, F.; Cao, W.; Tian, H.; Zhang, K.; Zheng, H.; Liu, X. Review of Seneca Valley Virus: A Call for Increased Surveillance and Research. Front. Microbiol. 2018, 9, 940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Zhu, Z.; Cao, W.; Liu, H.; Zhang, K.; Tian, H.; Liu, X.; Zheng, H. Immunogenicity and Protective Efficacy of an Inactivated Cell Culture-Derived Seneca Valley Virus Vaccine in Pigs. Vaccine 2018, 36, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Saporiti, V.; Fritzen, J.T.T.; Feronato, C.; Leme, R.A.; Lobato, Z.I.P.; Alfieri, A.F.; Alfieri, A.A. A Ten Years (2007–2016) Retrospective Serological Survey for Seneca Valley Virus Infection in Major Pig Producing States of Brazil. Vet. Res. Commun. 2017, 41, 317–321. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Nucleotide Sequence (5′→3′) | Restriction Sites |
|---|---|---|
| SVA-3AB-F | 5′ GAGGAATTCAGCCCTAACGA 3′ | EcoR I |
| SVA-3AB-R | 5′ GCGCTCGAGTCACTGTTGCATTTC 3′ | Xho I |
| SVA-VP1-F | 5′ CGCGGATCCTCCACCGACAACGCCGAGACT 3′ | BamH I |
| SVA-VP1-R | 5′ GCTCGAGTTGCATCAGCATC 3′ | Xho l |
| SVA-2C-F | 5′ GAGGAATTCGGACCCATGGATA 3′ | EcoR I |
| SVA-2C-R | 5′ GCGCTCGAGCTGTAGAACCAGA 3′ | Xhol I |
| SVA-3C-F | 5′ GAGGAATTCCAGCCCAACGTGGACA 3′ | EcoR I |
| SVA-3C-R | 5′ GCGCTCGAGTTGCATTGTAGCC 3′ | Xho l |
| SVA-3D-F | 5′ GAGGAATTCGGACTGATGACTG 3′ | EcoR I |
| SVA-3D-R | 5′ CTCAAGCTTGTCGAACAAGGCCCTC 3′ | Hind III |
| SVA-L-F | 5′ GAGGAATTCCAGAACTCTAATTT 3′ | EcoR I |
| SVA-L-R | 5′ TATCTCGAGCTGTAGTTCGTATACGATG 3′ | Xho I |
| Detection Methods | VNT Results | |||
|---|---|---|---|---|
| Positive | Negative | Total | ||
| 3AB indirect ELISA results | Positive | 42 | 4 | 46 |
| Negative | 4 | 50 | 54 | |
| Data analysis | Total | 46 | 54 | 100 |
| Sensitivity 1 | 91.30% | - | - | |
| Specificity 2 | - | 92.59% | - | |
| Coincidence rate 3 | - | - | 92.00% | |
| Province | Positive 1 | Negative 1 | Total 1 | Ratio (%) |
|---|---|---|---|---|
| Shandong | 730 | 170 | 900 | 81.11 |
| Jiangsu | 762 | 150 | 912 | 83.55 |
| Jiangxi | 512 | 119 | 631 | 73.25 |
| Zhejiang | 559 | 140 | 699 | 79.97 |
| Henan | 298 | 88 | 386 | 77.20 |
| Hebei | 125 | 18 | 143 | 87.41 |
| Anhui | 49 | 5 | 54 | 90.74 |
| Sichuan | 35 | 0 | 35 | 100.00 |
| Guangdong | 119 | 5 | 124 | 95.97 |
| Fujian | 12 | 5 | 17 | 70.59 |
| Shanxi | 29 | 0 | 29 | 100.00 |
| Total | 3230 | 700 | 3930 | 82.19 |
| 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | |
|---|---|---|---|---|---|---|---|---|---|
| Shandong | 84.30 | 95.15 | 98.97 | 93.88 | 74.58 | 67.57 | 71.43 | - | 39.29 |
| Jiangxi | 96.67 | 96.67 | 99.03 | 82.05 | 80.00 | 74.00 | 75.00 | 66.67 | 37.50 |
| Jiangsu | 86.89 | 92.31 | 98.92 | 85.92 | 81.00 | - | - | 55.21 | 75.00 |
| Zhejiang | 54.24 | 98.67 | 98.60 | 69.88 | 86.67 | 60.26 | - | - | 81.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, J.; Gao, Y.; Li, J.; Li, M.; Guo, C.; Bai, J.; Jiang, P. The Establishment and Application of Indirect 3AB-ELISA for the Detection of Antibodies against Senecavirus A. Viruses 2023, 15, 861. https://doi.org/10.3390/v15040861
Yan J, Gao Y, Li J, Li M, Guo C, Bai J, Jiang P. The Establishment and Application of Indirect 3AB-ELISA for the Detection of Antibodies against Senecavirus A. Viruses. 2023; 15(4):861. https://doi.org/10.3390/v15040861
Chicago/Turabian StyleYan, Junfang, Yanni Gao, Jian Li, Minjing Li, Chengyi Guo, Juan Bai, and Ping Jiang. 2023. "The Establishment and Application of Indirect 3AB-ELISA for the Detection of Antibodies against Senecavirus A" Viruses 15, no. 4: 861. https://doi.org/10.3390/v15040861