
Immune Molecules’ mRNA Expression in Porcine Alveolar Macrophages Co-Infected with Porcine Reproductive and Respiratory Syndrome Virus and Porcine Circovirus Type 2
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Trial Design
2.3. Immunofluorescence Assay (IFA)
2.4. Quantitative Real-Time RT-PCR Assay (RT-qPCR)
2.4.1. Detection of PRRSV RNA
2.4.2. Detection of PCV2 DNA
2.4.3. Detection of Porcine Immune Molecules
2.5. Viral Titer Assay (TCID50)
2.6. Statistical Analysis
3. Results
3.1. Confirmation of PRRSV or PCV2 Infection in PAMs
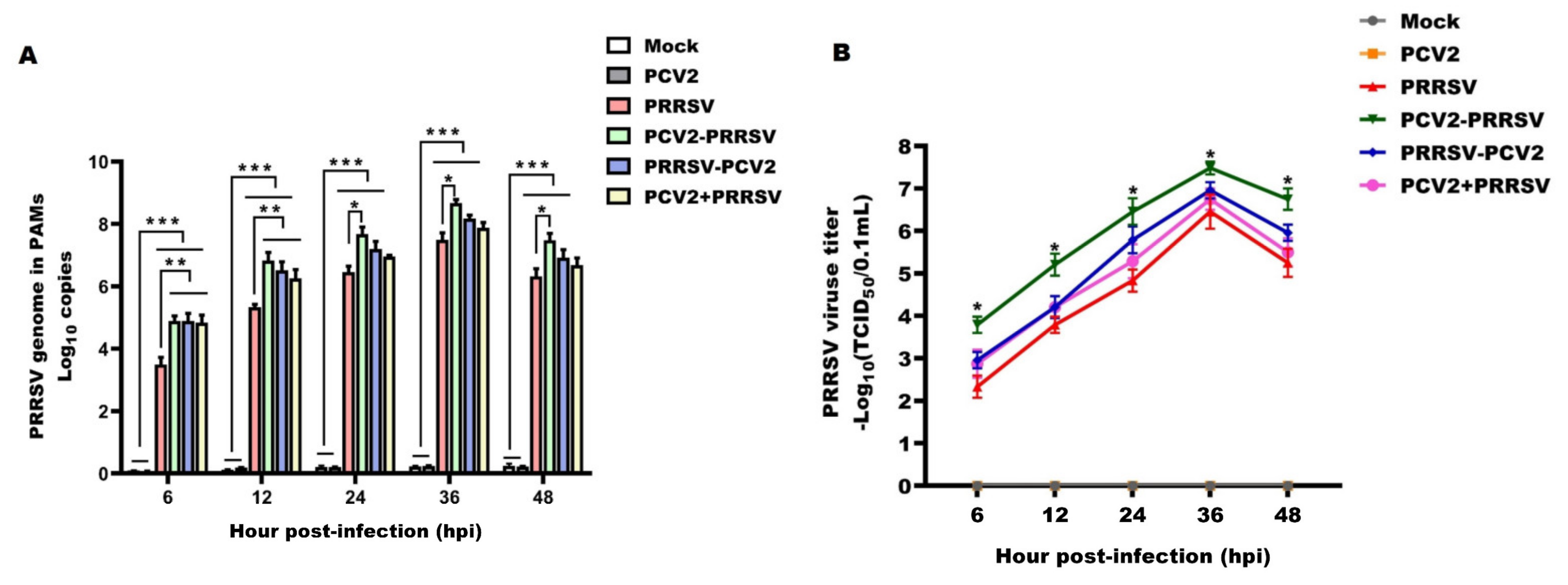
3.2. PRRSV Virologic Parameters
3.3. PCV2 Virologic Parameters
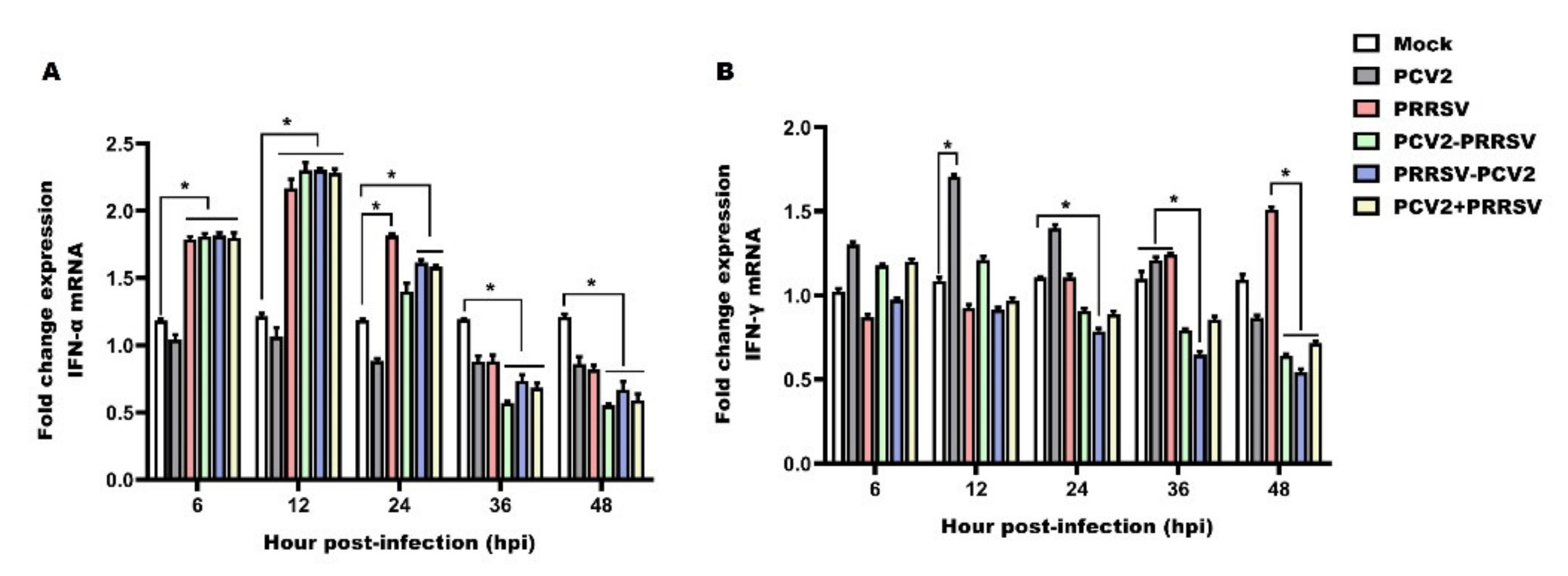
3.4. Detection of mRNA Levels of IFN-α and IFN-γ
3.5. Detection of mRNA Levels of TNF-α and IL-1β
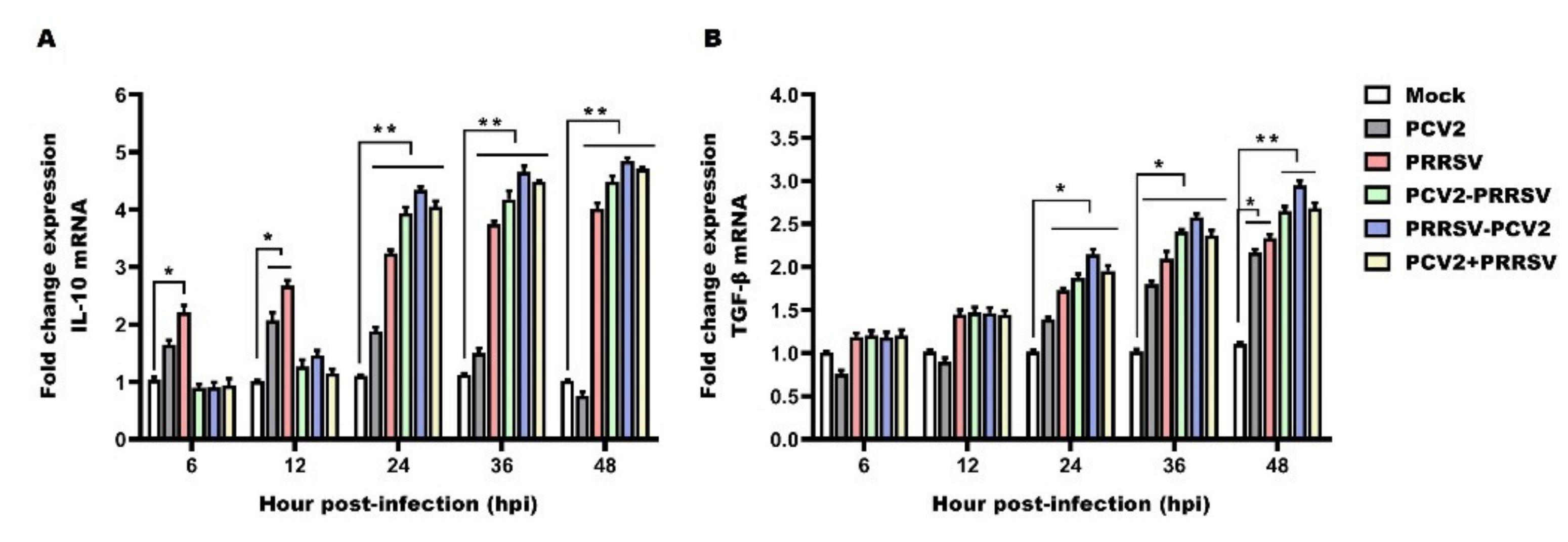
3.6. Detection of mRNA Levels of IL-10 and TGF-β
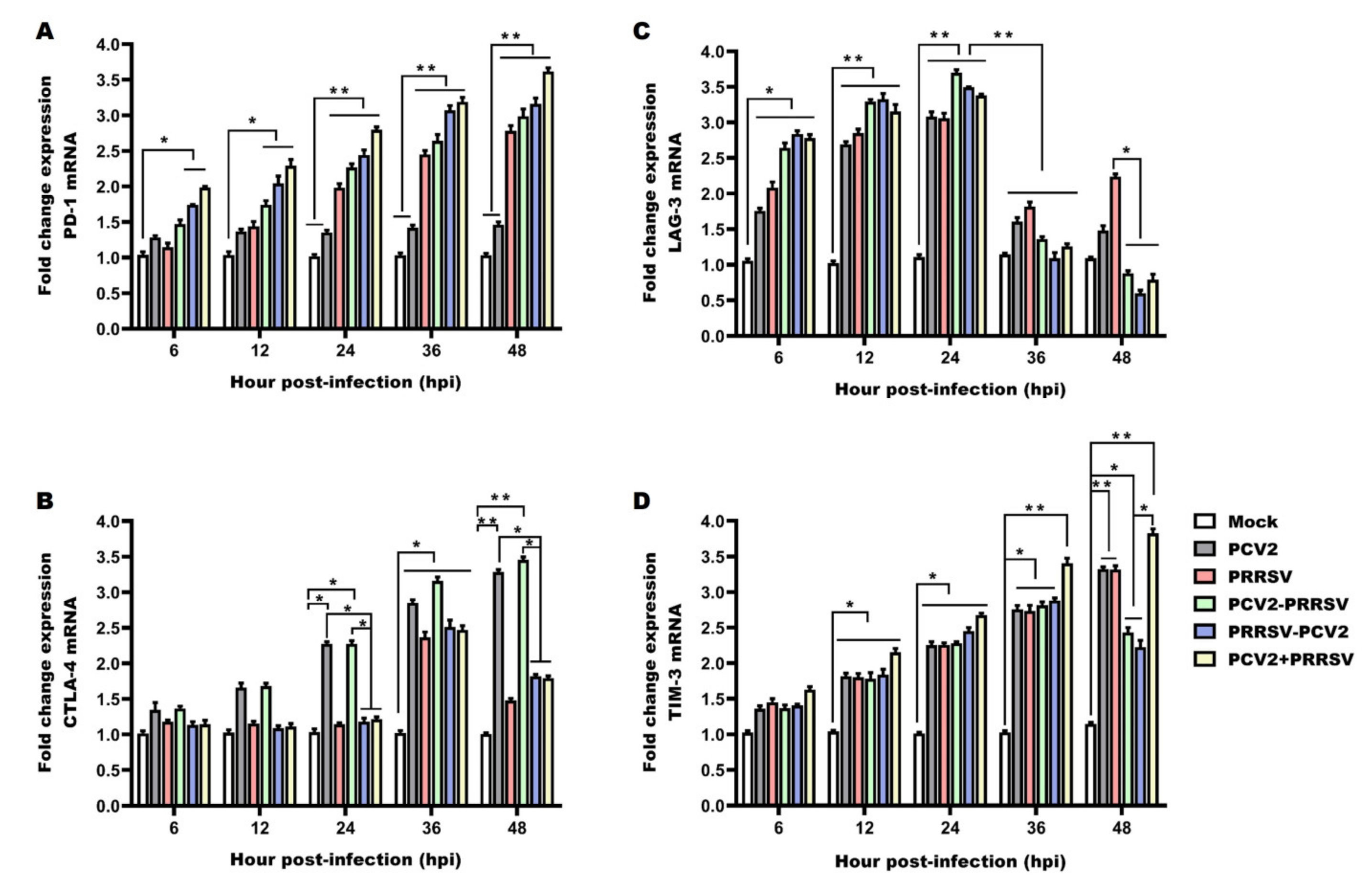
3.7. Detection of mRNA Levels of Immune Checkpoints
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ma, Z.; Han, Z.; Liu, Z.; Meng, F.; Wang, H.; Cao, L.; Li, Y.; Jiao, Q.; Liu, S.; Liu, M. Epidemiological investigation of porcine pseudorabies virus and its coinfection rate in Shandong Province in China from 2015 to 2018. J. Vet. Sci. 2020, 21, e36. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, T.; Zhang, X.; Liu, X.; Ren, L. Co-Infection of Swine with Porcine Circovirus Type 2 and Other Swine Viruses. Viruses 2019, 11, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruedas-Torres, I.; Rodriguez-Gomez, I.M.; Sanchez-Carvajal, J.M.; Larenas-Munoz, F.; Pallares, F.J.; Carrasco, L.; Gomez-Laguna, J. The jigsaw of PRRSV virulence. Vet. Microbiol. 2021, 260, 109168. [Google Scholar] [CrossRef] [PubMed]
- Brinton, M.A.; Gulyaeva, A.A.; Balasuriya, U.B.R.; Dunowska, M.; Faaberg, K.S.; Goldberg, T.; Leung, F.C.C.; Nauwynck, H.J.; Snijder, E.J.; Stadejek, T.; et al. ICTV Virus Taxonomy Profile: Arteriviridae 2021. J. Gen. Virol. 2021, 102, 001632. [Google Scholar] [CrossRef] [PubMed]
- Darwich, L.; Gimeno, M.; Sibila, M.; Diaz, I.; de la Torre, E.; Dotti, S.; Kuzemtseva, L.; Martin, M.; Pujols, J.; Mateu, E. Genetic and immunobiological diversities of porcine reproductive and respiratory syndrome genotype I strains. Vet. Microbiol. 2011, 150, 49–62. [Google Scholar] [CrossRef]
- Wensvoort, G.; de Kluyver, E.P.; Luijtze, E.A.; den Besten, A.; Harris, L.; Collins, J.E.; Christianson, W.T.; Chladek, D. Antigenic comparison of Lelystad virus and swine infertility and respiratory syndrome (SIRS) virus. J. Vet. Diagn. Investig. 1992, 4, 134–138. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, J.; Wu, X.; Ma, D.; Zhang, X.; Li, R.; Han, C.; Liu, H.; Yin, X.; Du, Q.; et al. PCV2 targets cGAS to inhibit type I interferon induction to promote other DNA virus infection. PLoS Pathog. 2021, 17, e1009940. [Google Scholar] [CrossRef]
- Link, E.K.; Eddicks, M.; Nan, L.; Ritzmann, M.; Sutter, G.; Fux, R. Discriminating the eight genotypes of the porcine circovirus type 2 with TaqMan-based real-time PCR. Virol. J. 2021, 18, 70. [Google Scholar] [CrossRef]
- Kim, D.; Ha, Y.; Oh, Y.; Chae, C. Prevalence of porcine circovirus types 2a and b in pigs with and without post-weaning multi-systemic wasting syndrome. Vet. J. 2011, 188, 115–117. [Google Scholar] [CrossRef]
- Opriessnig, T.; Halbur, P.G. Concurrent infections are important for expression of porcine circovirus associated disease. Virus. Res. 2012, 164, 20–32. [Google Scholar] [CrossRef]
- Ge, X.; Wang, F.; Guo, X.; Yang, H. Porcine circovirus type 2 and its associated diseases in China. Virus Res 2012, 164, 100–106. [Google Scholar] [CrossRef]
- Pallares, F.J.; Halbur, P.G.; Opriessnig, T.; Sorden, S.D.; Villar, D.; Janke, B.H.; Yaeger, M.J.; Larson, D.J.; Schwartz, K.J.; Yoon, K.J.; et al. Porcine circovirus type 2 (PCV-2) coinfections in US field cases of postweaning multisystemic wasting syndrome (PMWS). J. Vet. Diagn. Investig. 2002, 14, 515–519. [Google Scholar] [CrossRef] [Green Version]
- Niederwerder, M.C.; Jaing, C.J.; Thissen, J.B.; Cino-Ozuna, A.G.; McLoughlin, K.S.; Rowland, R.R. Microbiome associations in pigs with the best and worst clinical outcomes following co-infection with porcine reproductive and respiratory syndrome virus (PRRSV) and porcine circovirus type 2 (PCV2). Vet. Microbiol. 2016, 188, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Drolet, R.; Larochelle, R.; Morin, M.; Delisle, B.; Magar, R. Detection rates of porcine reproductive and respiratory syndrome virus, porcine circovirus type 2, and swine influenza virus in porcine proliferative and necrotizing pneumonia. Vet. Pathol. 2003, 40, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Burgara-Estrella, A.; Montalvo-Corral, M.; Bolanos, A.; Ramirez-Mendoza, H.; Valenzuela, O.; Hernandez, J. Naturally co-infected boars with both porcine reproductive and respiratory syndrome virus and porcine circovirus type 2. Transbound. Emerg. Dis. 2012, 59, 532–538. [Google Scholar] [CrossRef]
- Fan, P.; Wei, Y.; Guo, L.; Wu, H.; Huang, L.; Liu, J.; Liu, C. Synergistic effects of sequential infection with highly pathogenic porcine reproductive and respiratory syndrome virus and porcine circovirus type 2. Virol. J. 2013, 10, 265. [Google Scholar] [CrossRef] [Green Version]
- Segales, J.; Domingo, M.; Chianini, F.; Majo, N.; Dominguez, J.; Darwich, L.; Mateu, E. Immunosuppression in postweaning multisystemic wasting syndrome affected pigs. Vet. Microbiol. 2004, 98, 151–158. [Google Scholar] [CrossRef]
- Niederwerder, M.C.; Bawa, B.; Serao, N.V.; Trible, B.R.; Kerrigan, M.A.; Lunney, J.K.; Dekkers, J.C.; Rowland, R.R. Vaccination with a Porcine Reproductive and Respiratory Syndrome (PRRS) Modified Live Virus Vaccine Followed by Challenge with PRRS Virus and Porcine Circovirus Type 2 (PCV2) Protects against PRRS but Enhances PCV2 Replication and Pathogenesis Compared to Results for Nonvaccinated Cochallenged Controls. Clin. Vaccine Immunol. 2015, 22, 1244–1254. [Google Scholar] [CrossRef] [Green Version]
- Barnabei, L.; Laplantine, E.; Mbongo, W.; Rieux-Laucat, F.; Weil, R. NF-kappaB: At the Borders of Autoimmunity and Inflammation. Front. Immunol. 2021, 12, 716469. [Google Scholar] [CrossRef]
- Cai, H.; Liu, G.; Zhong, J.; Zheng, K.; Xiao, H.; Li, C.; Song, X.; Li, Y.; Xu, C.; Wu, H.; et al. Immune Checkpoints in Viral Infections. Viruses 2020, 12, 1051. [Google Scholar] [CrossRef]
- Wykes, M.N.; Lewin, S.R. Immune checkpoint blockade in infectious diseases. Nat. Rev. Immunol. 2018, 18, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sun, Y.; Zhao, S.; Cui, Z.; Chen, Y.; Xu, P.; Chen, J.; Zhang, Y.; Xia, P. Differences in Humoral Immune Response against the Type 2 Porcine Reproductive and Respiratory Syndrome Virus via Different Immune Pathways. Viruses 2022, 14, 1435. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Li, W.; Zhao, S.; Cui, Z.; Chen, Y.; Zhang, Y.N.; Chen, J.; Xia, P. Proteomic Characterization of PAMs with PRRSV-ADE Infection. Viruses 2022, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Shi, X.; Zhang, X.; Wang, A.; Wang, L.; Yang, Y.; Deng, R.; Zhang, G.P. MicroRNA 373 Facilitates the Replication of Porcine Reproductive and Respiratory Syndrome Virus by Its Negative Regulation of Type I Interferon Induction. J. Virol. 2017, 91, e01311-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuire, K.; Glass, E.J. The expanding role of microarrays in the investigation of macrophage responses to pathogens. Vet. Immunol. Immunopathol. 2005, 105, 259–275. [Google Scholar] [CrossRef]
- Gilpin, D.F.; McCullough, K.; Meehan, B.M.; McNeilly, F.; McNair, I.; Stevenson, L.S.; Foster, J.C.; Ellis, J.A.; Krakowka, S.; Adair, B.M.; et al. In vitro studies on the infection and replication of porcine circovirus type 2 in cells of the porcine immune system. Vet. Immunol. Immunopathol. 2003, 94, 149–161. [Google Scholar] [CrossRef]
- Allan, G.M.; McNeilly, F.; Ellis, J.; Krakowka, S.; Meehan, B.; McNair, I.; Walker, I.; Kennedy, S. Experimental infection of colostrum deprived piglets with porcine circovirus 2 (PCV2) and porcine reproductive and respiratory syndrome virus (PRRSV) potentiates PCV2 replication. Arch. Virol. 2000, 145, 2421–2429. [Google Scholar] [CrossRef]
- Wellenberg, G.J.; Stockhofe-Zurwieden, N.; Boersma, W.J.; De Jong, M.F.; Elbers, A.R. The presence of co-infections in pigs with clinical signs of PMWS in The Netherlands: A case-control study. Res. Vet. Sci. 2004, 77, 177–184. [Google Scholar] [CrossRef]
- Harms, P.A.; Sorden, S.D.; Halbur, P.G.; Bolin, S.R.; Lager, K.M.; Morozov, I.; Paul, P.S. Experimental reproduction of severe disease in CD/CD pigs concurrently infected with type 2 porcine circovirus and porcine reproductive and respiratory syndrome virus. Vet. Pathol. 2001, 38, 528–539. [Google Scholar] [CrossRef]
- Rovira, A.; Balasch, M.; Segales, J.; Garcia, L.; Plana-Duran, J.; Rosell, C.; Ellerbrok, H.; Mankertz, A.; Domingo, M. Experimental inoculation of conventional pigs with porcine reproductive and respiratory syndrome virus and porcine circovirus 2. J. Virol. 2002, 76, 3232–3239. [Google Scholar] [CrossRef] [Green Version]
- Sinha, A.; Shen, H.G.; Schalk, S.; Beach, N.M.; Huang, Y.W.; Meng, X.J.; Halbur, P.G.; Opriessnig, T. Porcine reproductive and respiratory syndrome virus (PRRSV) influences infection dynamics of porcine circovirus type 2 (PCV2) subtypes PCV2a and PCV2b by prolonging PCV2 viremia and shedding. Vet. Microbiol. 2011, 152, 235–246. [Google Scholar] [CrossRef]
- Chang, H.W.; Pang, V.F.; Chen, L.J.; Chia, M.Y.; Tsai, Y.C.; Jeng, C.R. Bacterial lipopolysaccharide induces porcine circovirus type 2 replication in swine alveolar macrophages. Vet. Microbiol. 2006, 115, 311–319. [Google Scholar] [CrossRef]
- Chang, H.W.; Jeng, C.R.; Liu, J.J.; Lin, T.L.; Chang, C.C.; Chia, M.Y.; Tsai, Y.C.; Pang, V.F. Reduction of porcine reproductive and respiratory syndrome virus (PRRSV) infection in swine alveolar macrophages by porcine circovirus 2 (PCV2)-induced interferon-alpha. Vet. Microbiol. 2005, 108, 167–177. [Google Scholar] [CrossRef]
- Opriessnig, T.; Ramamoorthy, S.; Madson, D.M.; Patterson, A.R.; Pal, N.; Carman, S.; Meng, X.J.; Halbur, P.G. Differences in virulence among porcine circovirus type 2 isolates are unrelated to cluster type 2a or 2b and prior infection provides heterologous protection. J. Gen. Virol. 2008, 89 Pt 10, 2482–2491. [Google Scholar] [CrossRef]
- Yin, S.H.; Xiao, C.T.; Gerber, P.F.; Beach, N.M.; Meng, X.J.; Halbur, P.G.; Opriessnig, T. Concurrent porcine circovirus type 2a (PCV2a) or PCV2b infection increases the rate of amino acid mutations of porcine reproductive and respiratory syndrome virus (PRRSV) during serial passages in pigs. Virus Res. 2013, 178, 445–451. [Google Scholar] [CrossRef]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Chang, H.W.; Jeng, C.R.; Lin, T.L.; Lin, C.M.; Wan, C.H.; Pang, V.F. The effect of infection order of porcine circovirus type 2 and porcine reproductive and respiratory syndrome virus on dually infected swine alveolar macrophages. BMC Vet. Res. 2012, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- Albina, E.; Carrat, C.; Charley, B. Interferon-alpha response to swine arterivirus (PoAV), the porcine reproductive and respiratory syndrome virus. J. Interferon. Cytokine Res. 1998, 18, 485–490. [Google Scholar] [CrossRef]
- Li, W.; Liu, S.; Wang, Y.; Deng, F.; Yan, W.; Yang, K.; Chen, H.; He, Q.; Charreyre, C.; Audoneet, J.C. Transcription analysis of the porcine alveolar macrophage response to porcine circovirus type 2. BMC Genom. 2013, 14, 353. [Google Scholar] [CrossRef] [Green Version]
- Shi, K.C.; Guo, X.; Ge, X.N.; Liu, Q.; Yang, H.C. Cytokine mRNA expression profiles in peripheral blood mononuclear cells from piglets experimentally co-infected with porcine reproductive and respiratory syndrome virus and porcine circovirus type 2. Vet. Microbiol. 2010, 140, 155–160. [Google Scholar] [CrossRef]
- Chen, A.Y.; Wolchok, J.D.; Bass, A.R. TNF in the era of immune checkpoint inhibitors: Friend or foe? Nat. Rev. Rheumatol. 2021, 17, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Interleukin-1 in the pathogenesis and treatment of inflammatory diseases. Blood 2011, 117, 3720–3732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thanawongnuwech, R.; Thacker, B.; Halbur, P.; Thacker, E.L. Increased production of proinflammatory cytokines following infection with porcine reproductive and respiratory syndrome virus and Mycoplasma hyopneumoniae. Clin. Diagn. Lab. Immunol. 2004, 11, 901–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniam, S.; Beura, L.K.; Kwon, B.; Pattnaik, A.K.; Osorio, F.A. Amino acid residues in the non-structural protein 1 of porcine reproductive and respiratory syndrome virus involved in down-regulation of TNF-alpha expression in vitro and attenuation in vivo. Virology 2012, 432, 241–249. [Google Scholar] [CrossRef]
- Darwich, L.; Pie, S.; Rovira, A.; Segales, J.; Domingo, M.; Oswald, I.P.; Mateu, E. Cytokine mRNA expression profiles in lymphoid tissues of pigs naturally affected by postweaning multisystemic wasting syndrome. J. Gen. Virol. 2003, 84 Pt 8, 2117–2125. [Google Scholar] [CrossRef]
- Sipos, W.; Duvigneau, J.C.; Willheim, M.; Schilcher, F.; Hartl, R.T.; Hofbauer, G.; Exel, B.; Pietschmann, P.; Schmoll, F. Systemic cytokine profile in feeder pigs suffering from natural postweaning multisystemic wasting syndrome (PMWS) as determined by semiquantitative RT-PCR and flow cytometric intracellular cytokine detection. Vet. Immunol. Immunopathol. 2004, 99, 63–71. [Google Scholar] [CrossRef]
- Batlle, E.; Massague, J. Transforming Growth Factor-beta Signaling in Immunity and Cancer. Immunity 2019, 50, 924–940. [Google Scholar] [CrossRef]
- Larson, C.; Oronsky, B.; Carter, C.A.; Oronsky, A.; Knox, S.J.; Sher, D.; Reid, T.R. TGF-beta: A master immune regulator. Expert. Opin. Ther. Targets 2020, 24, 427–438. [Google Scholar] [CrossRef]
- Richmond, O.; Cecere, T.E.; Erdogan, E.; Meng, X.J.; Pineyro, P.; Subramaniam, S.; Todd, S.M.; LeRoith, T. The PD-L1/CD86 ratio is increased in dendritic cells co-infected with porcine circovirus type 2 and porcine reproductive and respiratory syndrome virus, and the PD-L1/PD-1 axis is associated with anergy, apoptosis, and the induction of regulatory T-cells in porcine lymphocytes. Vet. Microbiol. 2015, 180, 223–229. [Google Scholar] [CrossRef]
- Andersson, M.; Ahlberg, V.; Jensen-Waern, M.; Fossum, C. Intestinal gene expression in pigs experimentally co-infected with PCV2 and PPV. Vet. Immunol. Immunopathol. 2011, 142, 72–80. [Google Scholar] [CrossRef]
- Feng, W.H.; Tompkins, M.B.; Xu, J.S.; Zhang, H.X.; McCaw, M.B. Analysis of constitutive cytokine expression by pigs infected in-utero with porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 2003, 94, 35–45. [Google Scholar] [CrossRef]
- Suradhat, S.; Thanawongnuwech, R. Upregulation of interleukin-10 gene expression in the leukocytes of pigs infected with porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 2003, 84 Pt 10, 2755–2760. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, P.; Xie, C.; Ha, Z.; Shi, N.; Zhang, H.; Li, Z.; Han, J.; Xie, Y.; Qiu, X.; et al. Pathogenicity by Coinfection and Sequential Infection with NADC30-like PRRSV and PCV2 in Post-Weaned Pigs. Viruses 2022, 14, 193. [Google Scholar] [CrossRef]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [Green Version]
- Yue, F.; Zhu, Y.P.; Zhang, Y.F.; Sun, G.P.; Yang, Y.; Guo, D.G.; Wang, A.G.; Li, B.W.; Yin, M.; Cheng, A.C.; et al. Up-regulated expression of PD-1 and its ligands during acute Classical Swine Fever virus infection in swine. Res. Vet. Sci. 2014, 97, 251–256. [Google Scholar] [CrossRef]
- Muhlbauer, M.; Fleck, M.; Schutz, C.; Weiss, T.; Froh, M.; Blank, C.; Scholmerich, J.; Hellerbrand, C. PD-L1 is induced in hepatocytes by viral infection and by interferon-alpha and -gamma and mediates T cell apoptosis. J. Hepatol. 2006, 45, 520–528. [Google Scholar] [CrossRef]
- Richmond, O.; Cecere, T.E.; Erdogan, E.; Meng, X.J.; Pineyro, P.; Subramaniam, S.; Todd, S.M.; LeRoith, T. PD-L1 expression is increased in monocyte derived dendritic cells in response to porcine circovirus type 2 and porcine reproductive and respiratory syndrome virus infections. Vet. Immunol. Immunopathol. 2015, 168, 24–29. [Google Scholar] [CrossRef]
- Linsley, P.S.; Greene, J.L.; Tan, P.; Bradshaw, J.; Ledbetter, J.A.; Anasetti, C.; Damle, N.K. Coexpression and functional cooperation of CTLA-4 and CD28 on activated T lymphocytes. J. Exp. Med. 1992, 176, 1595–1604. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, S.; Kataoka, H.; Hara, S.; Yokosuka, T.; Takase, K.; Yamasaki, S.; Kobayashi, W.; Saito, Y.; Saito, T. In vivo overexpression of CTLA-4 suppresses lymphoproliferative diseases and thymic negative selection. Eur. J. Immunol. 2005, 35, 399–407. [Google Scholar] [CrossRef]
- Ruedas-Torres, I.; Rodriguez-Gomez, I.M.; Sanchez-Carvajal, J.M.; Guil-Luna, S.; Larenas-Munoz, F.; Pallares, F.J.; Carrasco, L.; Gomez-Laguna, J. Up-Regulation of Immune Checkpoints in the Thymus of PRRSV-1-Infected Piglets in a Virulence-Dependent Fashion. Front. Immunol. 2021, 12, 671743. [Google Scholar] [CrossRef]
- Verhagen, J.; Genolet, R.; Britton, G.J.; Stevenson, B.J.; Sabatos-Peyton, C.A.; Dyson, J.; Luescher, I.F.; Wraith, D. C, CTLA-4 controls the thymic development of both conventional and regulatory T cells through modulation of the TCR repertoire. Proc. Natl. Acad. Sci. USA 2013, 110, E221–E230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, F.; Cheng, A.; Zhu, Y.; Li, P.; Zhang, Y.; Sun, G.; Wang, M.; Wang, X. Overexpression of programmed death ligands in naturally occurring postweaning multisystemic wasting syndrome. Viral. Immunol. 2015, 28, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Anderson, A.C.; Kuchroo, V.K. TIM-3 and its regulatory role in immune responses. Curr. Top Microbiol. Immunol. 2011, 350, 1–15. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | The Infection Orders of PRRSV and PCV2 |
|---|---|
| Mock | no infected virus |
| PCV2 | infection with PCV2 alone |
| PRRSV | infection with PRRSV alone |
| PCV2–PRRSV | infection with PCV2 prior to PRRSV |
| PRRSV–PCV2 | infection with PRRSV prior to PCV2 |
| PCV2 + PRRSV | PCV2 and PRRSV co-infection |
| Genes | Primer Sequence (5’–3’) |
|---|---|
| PCV2 | F 5’-CCTCACCTATGACCCCTAT-3’ |
| R 5’TGTTGTTTGGTTGGAAGTA-3’ | |
| PRRSV | F 5’-AAACCAGTCCAGAGGCAAGG-3’ |
| R 5’GCAAACTAAACTCCACAGTGTAA-3’ | |
| IFN-α | F 5’-GGATCAGCAGCTCAGGG-3’ |
| R 5’GAGGGTGAGTCTGTGGAAGTA-3’ | |
| IFN-γ | F 5’-AAAGATAACCAGCCCATTC-3’ |
| R 5’-GTCATTCAGTTTCCCAGA-3’ | |
| TNF-α | F 5’-TGGTGGTGCCGACAGATGG-3’ |
| R 5’-GGCTGATGGTGTGAGTGAGGAA-3’ | |
| IL-1β | F 5’-ATGCTGAAGGCTCTCCACCTC-3’ |
| R 5’-TTGTTGCTATCATCTCCTTGCAC-3’ | |
| IL-10 | F 5’-GCATCCACTTCCCAACCA-3’ |
| R 5’GCAACAAGTCGCCCATCT-3’ | |
| TGF-β | F 5’-CTTACTGAGCATCTTGGACCTTA-3’ |
| R 5’CCACTGAGCCACAATGGAAA-3’ | |
| PD-1 | F 5’-AGCCCAAGCACTTCATCCTC-3’ |
| R 5’-TGTGGAAGTCTCGTCCGTTG-3’ | |
| LAG-3 | F 5’-CTCCTCCTGCTCCTTTTGGTT-3’ |
| R 5’-CAGCTCCCCAGTCTTGCTCT-3’ | |
| CTLA-4 | F 5’-TCTTCATCCCTGTCTTCTCCAAA-3’ |
| R 5’-GCAGACCCATACTCACACACAAA-3’ | |
| TIM-3 | F 5’-TTCGACGGGAGCAGTAAAGC-3’ |
| R 5’-AGGGCAGGACACAGTCAAAG-3’ | |
| β-actin | F 5’-CGGGACATCAAGGAGAAGC-3’ |
| R 5’-CTCGTTGCCGATGGTGATG-3’ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Z.; Zhou, L.; Hu, X.; Zhao, S.; Xu, P.; Li, W.; Chen, J.; Zhang, Y.; Xia, P. Immune Molecules’ mRNA Expression in Porcine Alveolar Macrophages Co-Infected with Porcine Reproductive and Respiratory Syndrome Virus and Porcine Circovirus Type 2. Viruses 2023, 15, 777. https://doi.org/10.3390/v15030777
Cui Z, Zhou L, Hu X, Zhao S, Xu P, Li W, Chen J, Zhang Y, Xia P. Immune Molecules’ mRNA Expression in Porcine Alveolar Macrophages Co-Infected with Porcine Reproductive and Respiratory Syndrome Virus and Porcine Circovirus Type 2. Viruses. 2023; 15(3):777. https://doi.org/10.3390/v15030777
Chicago/Turabian StyleCui, Zhiying, Likun Zhou, Xingxing Hu, Shijie Zhao, Pengli Xu, Wen Li, Jing Chen, Yina Zhang, and Pingan Xia. 2023. "Immune Molecules’ mRNA Expression in Porcine Alveolar Macrophages Co-Infected with Porcine Reproductive and Respiratory Syndrome Virus and Porcine Circovirus Type 2" Viruses 15, no. 3: 777. https://doi.org/10.3390/v15030777