
Mass Spectral Analyses of Salmonella Myovirus SPN3US Reveal Conserved and Divergent Themes in Proteolytic Maturation of Large Icosahedral Capsids

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Genome Sequencing of SPN3US Mutant am107
2.2. Preparation of Purified SPN3US Virions and Heads
2.3. Gel Electrophoresis and Mass Spectrometry Analysis of SPN3US Virions and Heads
3. Results
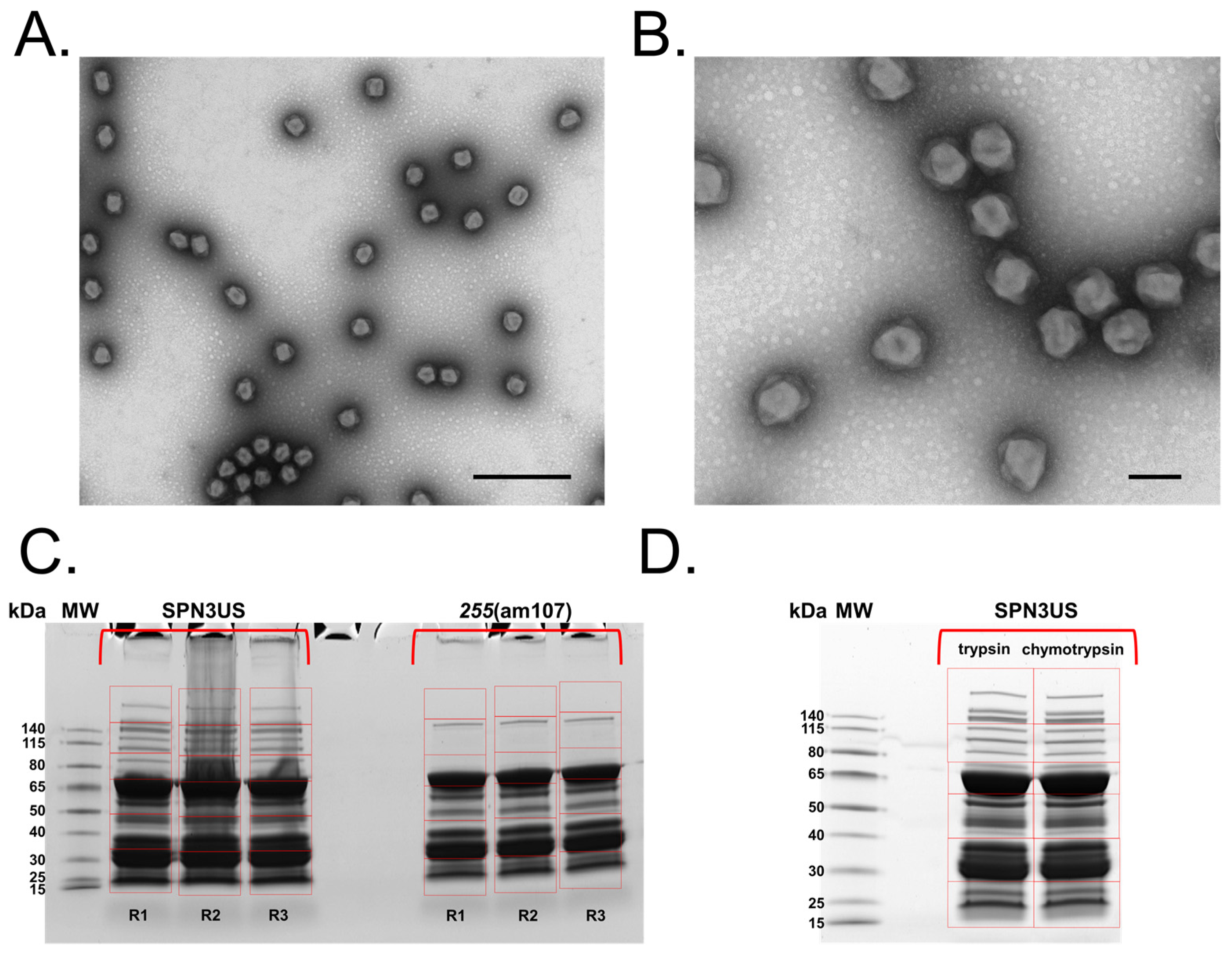
3.1. Isolation and Electron Microscopy of a Tail Tube Mutant of SPN3US
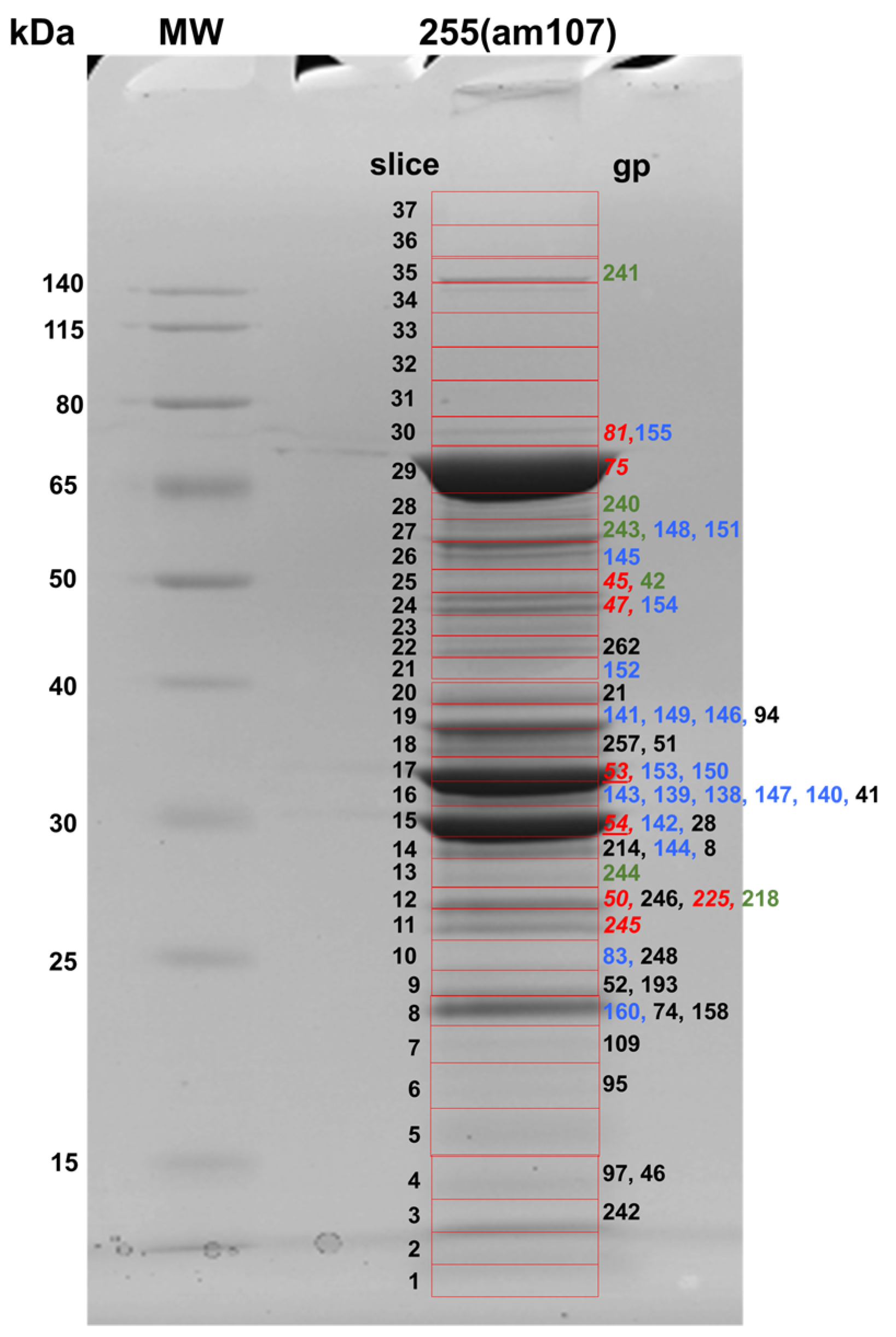
3.2. Proteomic Analyses of the SPN3US Virion and Head
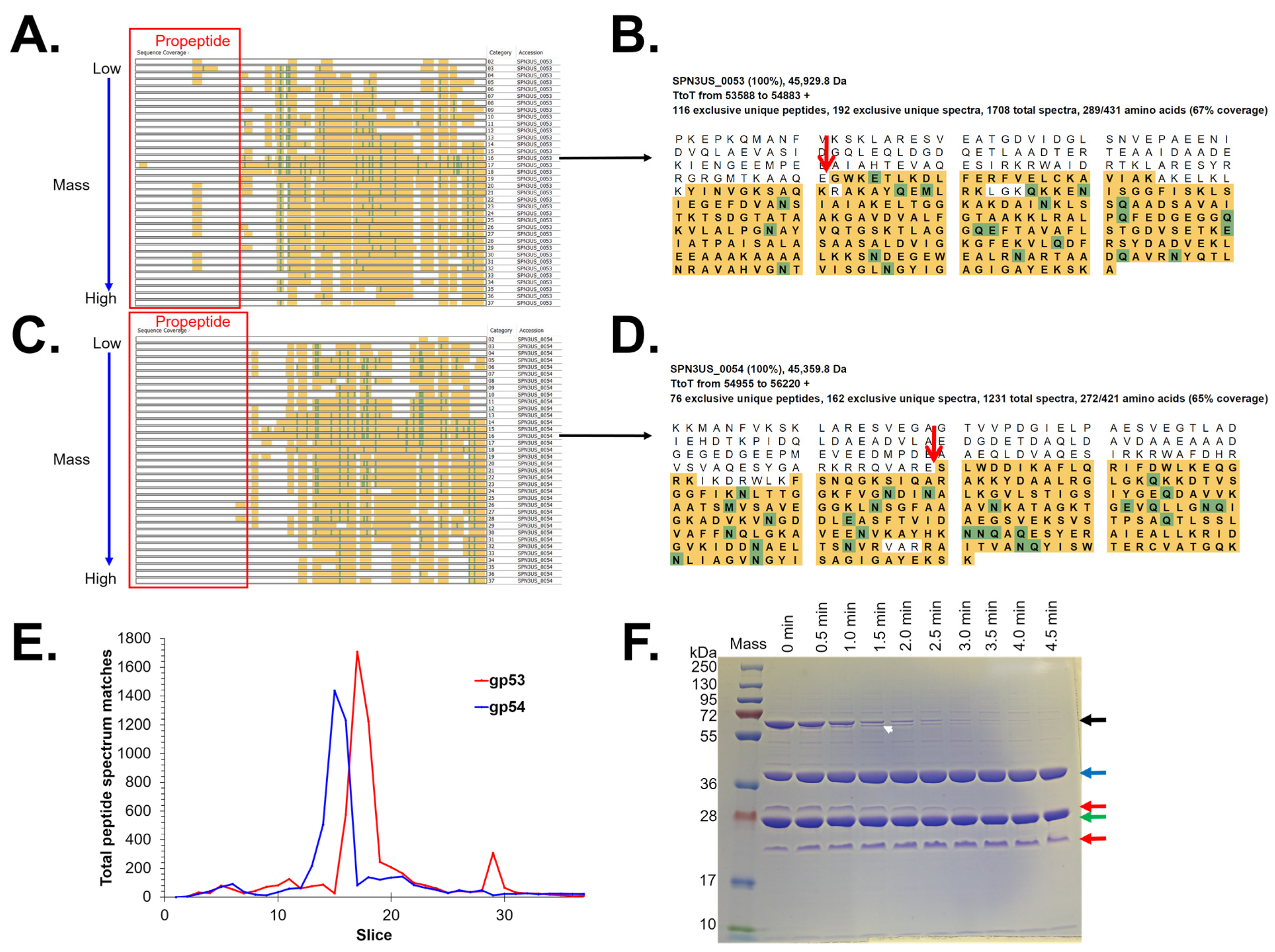
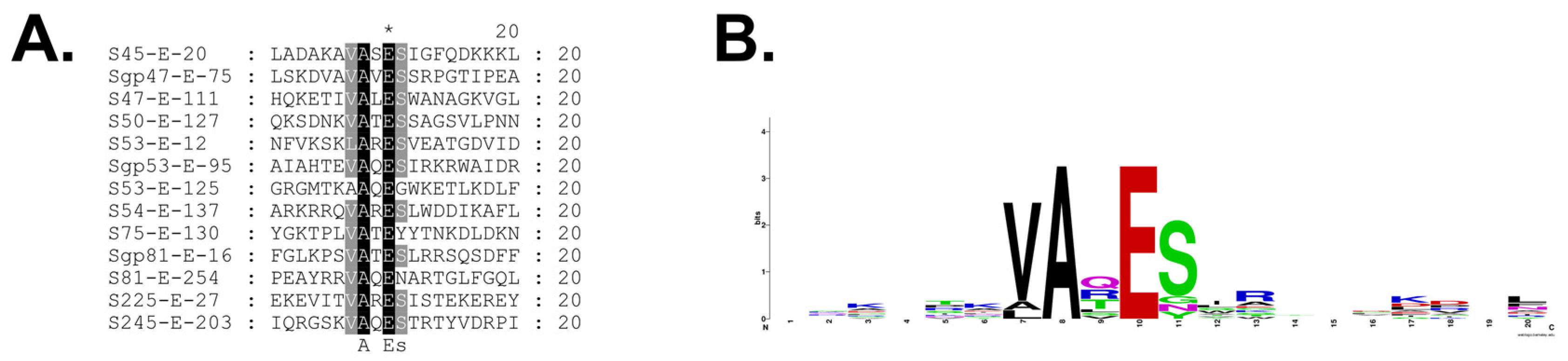
3.3. Identification of Prohead Protease Substrates and Cleavage Sites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aksyuk, A.A.; Rossmann, M.G. Bacteriophage assembly. Viruses 2011, 3, 172–203. [Google Scholar] [CrossRef] [PubMed]
- Dedeo, C.L.; Cingolani, G.; Teschke, C.M. Portal Protein: The Orchestrator of Capsid Assembly for the dsDNA Tailed Bacteriophages and Herpesviruses. Annu. Rev. Virol. 2019, 6, 141–160. [Google Scholar] [CrossRef]
- Prevelige, P.E.; Cortines, J.R. Phage assembly and the special role of the portal protein. Curr. Opin. Virol. 2018, 31, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Maurer, J.B.; Oh, B.; Moyer, C.L.; Duda, R.L. Capsids and Portals Influence Each Other’s Conformation During Assembly and Maturation. J. Mol. Biol. 2020, 432, 2015–2029. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Huet, A.; Lopez, C.A.; Toropova, K.; Pope, W.H.; Duda, R.L.; Hendrix, R.W.; Conway, J.F. Capsids and Genomes of Jumbo-Sized Bacteriophages Reveal the Evolutionary Reach of the HK97 Fold. mBio 2017, 8, e01579-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, L.M.; Anantharaman, V.; Krishnan, A.; Burroughs, A.M.; Aravind, L. Jumbo Phages: A Comparative Genomic Overview of Core Functions and Adaptions for Biological Conflicts. Viruses 2021, 13, 63. [Google Scholar] [CrossRef]
- Mesyanzhinov, V.V.; Robben, J.; Grymonprez, B.; Kostyuchenko, V.A.; Bourkaltseva, M.V.; Sykilinda, N.N.; Krylov, V.N.; Volckaert, G. The genome of bacteriophage PhiKZ of Pseudomonas aeruginosa. J. Mol. Biol. 2002, 317, 1–19. [Google Scholar] [CrossRef]
- Krylov, V.N.; Bourkaltseva, M.V.; Sykilinda, N.N.; Pleteneva, E.A.; Shaburova, O.V.; Kadykov, V.A.; Miller, S.; Biebl, M. Comparisons of the genomes of new giant phages isolated from environmental Pseudomonas aeruginosa strains of different regions. Russ. J. Genet. 2004, 40, 363–368. [Google Scholar] [CrossRef]
- Hendrix, R.W. Jumbo bacteriophages. In Lesser Known Large dsDNA Viruses; Van Etten, J., Ed.; Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2009; Volume 328, pp. 229–240. [Google Scholar]
- Lecoutere, E.; Ceyssens, P.-J.; Miroshnikov, K.A.; Mesyanzhinov, V.V.; Krylov, V.N.; Noben, J.-P.; Robben, J.; Hertveldt, K.; Volckaert, G.; Lavigne, R. Identification and comparative analysis of the structural proteomes of PhiKZ and EL, two giant Pseudomonas aeruginosa bacteriophages. Proteomics 2009, 9, 3215–3219. [Google Scholar] [CrossRef]
- Thomas, J.A.; Weintraub, S.T.; Wu, W.; Winkler, D.C.; Cheng, N.; Steven, A.C.; Black, L.W. Extensive proteolysis of head and inner body proteins by a morphogenetic protease in the giant Pseudomonas aeruginosa phage φKZ. Mol. Microbiol. 2012, 84, 324–339. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.A.; Weintraub, S.T.; Hakala, K.; Serwer, P.; Hardies, S.C. Proteome of the large Pseudomonas myovirus 201φ2-1: Delineation of proteolytically processed virion proteins. Mol. Cell. Proteom. 2010, 9, 940–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heymann, J.B.; Wang, B.; Newcomb, W.W.; Wu, W.; Winkler, D.C.; Cheng, N.; Reilly, E.R.; Hsia, R.C.; Thomas, J.A.; Steven, A.C. The Mottled Capsid of the Salmonella Giant Phage SPN3US, a Likely Maturation Intermediate with a Novel Internal Shell. Viruses 2020, 12, 910. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.A.; Benítez Quintana, A.D.; Bosch, M.A.; Coll De Peña, A.; Aguilera, E.; Coulibaly, A.; Wu, W.; Osier, M.V.; Hudson, A.O.; Weintraub, S.T.; et al. Identification of essential genes in the Salmonella phage SPN3US reveals novel insights into giant phage head structure and assembly. J. Virol. 2016, 90, 10284–10298. [Google Scholar] [CrossRef] [Green Version]
- Vander Byl, C.; Kropinski, A.M. Sequence of the genome of Salmonella bacteriophage P22. J. Bacteriol. 2000, 182, 6472–6481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popa, M.; McKelvey, T.A.; Hempel, J.; Hendrix, R.W. Bacteriophage HK97 structure: Wholesale covalent cross-linking between the major head shell subunits. J. Virol. 1991, 65, 3227–3237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, B.; Desmond, M.I.; Mallory, S.A.; Benitez, A.D.; Buckley, L.J.; Weintraub, S.T.; Osier, M.V.; Black, L.W.; Thomas, J.A. To Be or Not To Be T4: Evidence of a Complex Evolutionary Pathway of Head Structure and Assembly in Giant Salmonella Virus SPN3US. Front. Microbiol. 2017, 8, 2251. [Google Scholar] [CrossRef] [Green Version]
- Reilly, E.R.; Abajorga, M.K.; Kiser, C.; Mohd Redzuan, N.H.; Haidar, Z.; Adams, L.E.; Diaz, R.; Pinzon, J.A.; Hudson, A.O.; Black, L.W.; et al. A Cut above the Rest: Characterization of the Assembly of a Large Viral Icosahedral Capsid. Viruses 2020, 12, 725. [Google Scholar] [CrossRef]
- Thomas, J.A.; Black, L.W. Mutational analysis of the Pseudomonas aeruginosa myovirus φKZ morphogenetic protease gp175. J. Virol. 2013, 87, 8713–8725. [Google Scholar] [CrossRef] [Green Version]
- Coombs, D.H.; Arisaka, F. T4 tail structure and function. In Molecular Biology of Bacteriophage T4; Karam, J.D., Ed.; American Society for Microbiology: Washington, WA, USA, 1994. [Google Scholar]
- Sycheva, L.V.; Shneider, M.M.; Sykilinda, N.N.; Ivanova, M.A.; Miroshnikov, K.A.; Leiman, P.G. Crystal structure and location of gp131 in the bacteriophage phiKZ virion. Virology 2012, 434, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Conway, J.F.; Duda, R.L.; Cheng, N.; Hendrix, R.W.; Steven, A.C. Proteolytic and conformational control of virus capsid maturation: The bacteriophage HK97 system. J. Mol. Biol. 1995, 253, 86–99. [Google Scholar] [CrossRef]
- Cheng, H.; Shen, N.; Pei, J.; Grishin, N.V. Double-stranded DNA bacteriophage prohead protease is homologous to herpesvirus protease. Protein Sci. 2004, 13, 2260–2269. [Google Scholar] [CrossRef] [Green Version]
- Fokine, A.; Chipman, P.R.; Leiman, P.G.; Mesyanzhinov, V.V.; Rao, V.B.; Rossmann, M.G. Molecular architecture of the prolate head of bacteriophage T4. Proc. Natl. Acad. Sci. USA 2004, 101, 6003–6008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Showe, M.K.; Isobe, E.; Onorato, L. Bacteriophage T4 prehead proteinase: I. Purification and properties of a bacteriophage enzyme which cleaves the capsid precursor proteins. J. Mol. Biol. 1976, 107, 35–54. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, P.T. N-Terminal Methionine Processing. Curr. Protoc. Protein Sci. 2017, 88, 6.14.11–16.14.13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omotajo, D.; Tate, T.; Cho, H.; Choudhary, M. Distribution and diversity of ribosome binding sites in prokaryotic genomes. BMC Genom. 2015, 16, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Campbell, A.; Karlin, S. Correlations between Shine-Dalgarno sequences and gene features such as predicted expression levels and operon structures. J. Bacteriol. 2002, 184, 5733–5745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, L.; Tarkowski, H.-J. The N terminus of the head protein of T4 bacteriophage directs proteins to the GroEL chaperonin. J. Mol. Biol. 2005, 345, 375–386. [Google Scholar] [CrossRef]
- Mullaney, J.M.; Black, L.W. Capsid targeting sequence targets foreign proteins into bacteriophage T4 and permits proteolytic processing. J. Mol. Biol. 1996, 261, 372–385. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
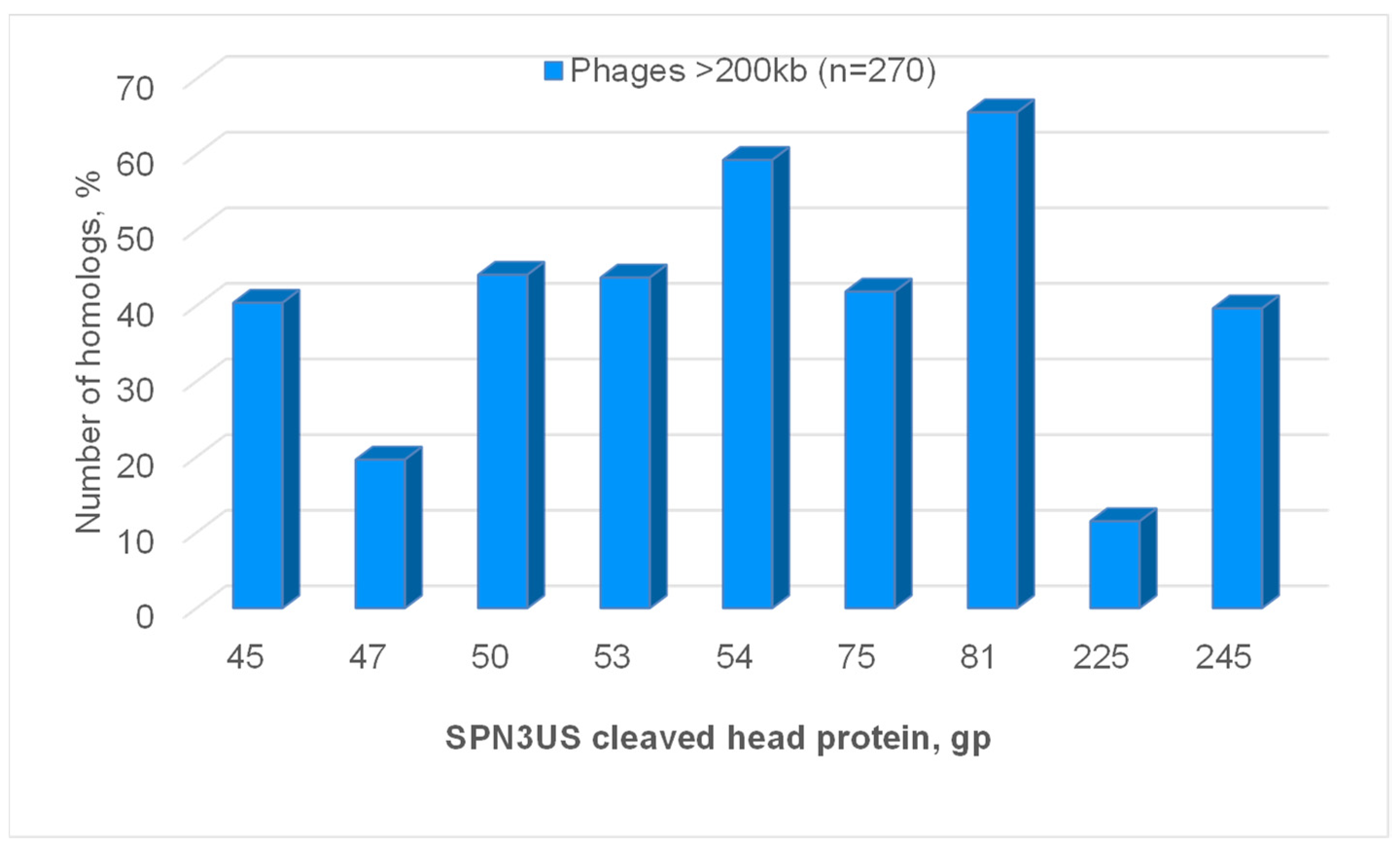{kind=link}
| Virions—Single Gradient Purified 1 | Virions—Double Gradient Purified 2 | Heads—Double Gradient Purified 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| R1 | R2 | R3 | R1 | R2 | R3 | |||
| Number of Proteins | Salmonella | 24 | 51 | 52 | 55 | 46 | 51 | 46 |
| Phage | 86 | 98 | 98 | 100 | 78 | 80 | 77 | |
| Number of PSM | Salmonella | 459 | 338 | 384 | 421 | 238 | 260 | 243 |
| Phage | 8507 | 26,892 | 26,216 | 26,569 | 19,288 | 18,990 | 18,496 | |
| Salmonella PSM/Phage PSM, % | 5.40 | 1.26 | 1.46 | 1.58 | 1.23 | 1.37 | 1.31 | |
| gp | Predicted Mass (kDa) | Mature Mass (kDa) | Propeptide Mass (kDa) | Cleavage Site (s) 1 | WT 10-Slice 2 | WT 6-Slice, Replicates 3,4 | Heads 6-Slice, Replicates 3,4 | Heads 37-Slice | WT 6-Slice | WT Chym 6-Slice | Protein Function/Comment |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Digestion enzyme: | Trypsin | Chymotrypsin | |||||||||
| 45 | 50.3 | 48.2 | 2.2 | ASE-20 | Y | Y(3) | Y(3) | Y | Y | Y | Head, unknown function |
| 47 | 63.3 | 50.7 | 12.5 | AVE-75 ALE-111 * | Y N | Y(2) Y(3#) | Y(1) Y(3#) | Y Y | Y Y | N N | Head protein, essential but unknown function |
| 50 | 41.5 | 25.6 | 15.8 | ATE-127 | Y | Y(3) | Y(3) | Y | Y | Y | Head |
| 53 | 45.2 | 31.5 | 13.8 | ARE-12 * AQE-95 * AQE-125 | N N Y | Y(3) Y(3) Y(3) | N Y(3) Y(3) | N Y Y | N Y Y | N Y N | High abundance E protein |
| 54 | 45.1 | 30.3 | 14.8 | AQE-124 * ARE-137 * | N N | N Y(3) | N Y(3) | N Y | N Y | Y# N | High abundance E protein |
| 75 | 83.9 | 70.4 | 13.5 | ATE-130 | Y | Y(3) | Y(3) | Y | Y | Y | Major capsid protein |
| 81 | 100.2 | 72.3 | 27.9 | ATE-161 AQE-254 * | Y N | Y(3#) N | Y(3) N | N N | Y N | Y Y | Portal |
| 225 | 25.1 | 22.3 | 2.8 | ARE-24 * | N | Y (3) | Y (3) | Y | Y | Y | Head, low abundance |
| 245 | 30.7 | 23.4 | 7.2 | AQE-203 | N | N | N | N | N | Y | Prohead protease |
| Erwinia Phage PhiEaH2 (JX316028) | Cronobacter phage CR5 (JX094500.1) | Pseudomonas Phage 201phi2-1 (EU197055) | Pseudomonas Phage PhiKZ (AF399011.1) | ||
|---|---|---|---|---|---|
| Nucleotide | Percent identity (%) | 75 | 69 | 83 | 85 |
| Query coverage (%) | 65 | 23 | <0.1 | <0.1 | |
| SPN3US proteins | All proteins (264) | 220 | 160 | 77 | 72 |
| Head proteins (55) | 49 | 48 | 26 | 23 | |
| Head proteins minus the 21 Paralog B proteins (34) | 31 | 32 | 18 | 19 | |
| Processed proteins (9) | 9 | 9 | 7 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scheuch, A.; Moran, S.A.M.; Faraone, J.N.; Unwin, S.R.; Vu, G.; Benítez, A.D.; Mohd Redzuan, N.H.; Molleur, D.; Pardo, S.; Weintraub, S.T.; et al. Mass Spectral Analyses of Salmonella Myovirus SPN3US Reveal Conserved and Divergent Themes in Proteolytic Maturation of Large Icosahedral Capsids. Viruses 2023, 15, 723. https://doi.org/10.3390/v15030723
Scheuch A, Moran SAM, Faraone JN, Unwin SR, Vu G, Benítez AD, Mohd Redzuan NH, Molleur D, Pardo S, Weintraub ST, et al. Mass Spectral Analyses of Salmonella Myovirus SPN3US Reveal Conserved and Divergent Themes in Proteolytic Maturation of Large Icosahedral Capsids. Viruses. 2023; 15(3):723. https://doi.org/10.3390/v15030723
Chicago/Turabian StyleScheuch, Aaron, Sara A. M. Moran, Julia N. Faraone, Sophia R. Unwin, Gialinh Vu, Andrea Denisse Benítez, Nurul Humaira Mohd Redzuan, Dana Molleur, Sammy Pardo, Susan T. Weintraub, and et al. 2023. "Mass Spectral Analyses of Salmonella Myovirus SPN3US Reveal Conserved and Divergent Themes in Proteolytic Maturation of Large Icosahedral Capsids" Viruses 15, no. 3: 723. https://doi.org/10.3390/v15030723