
Potentially Zoonotic Viruses in Wild Rodents, United Arab Emirates, 2019—A Pilot Study
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trapping and Sampling
2.2. Pathological Investigations
2.3. Preparation and Nucleic Acid Extraction of Ticks, Organ Samples, Oro-Pharyngeal Swabs, and Fecal Specimens
2.4. Screening of the Extracted Samples for Viral Nucleic Acids
2.5. Molecular Rodent Species Identification
2.6. Sequencing and Phylogenetic Analyses
3. Results
3.1. Patho-Histological Results
3.2. Viral Nucleic Acid Identifications
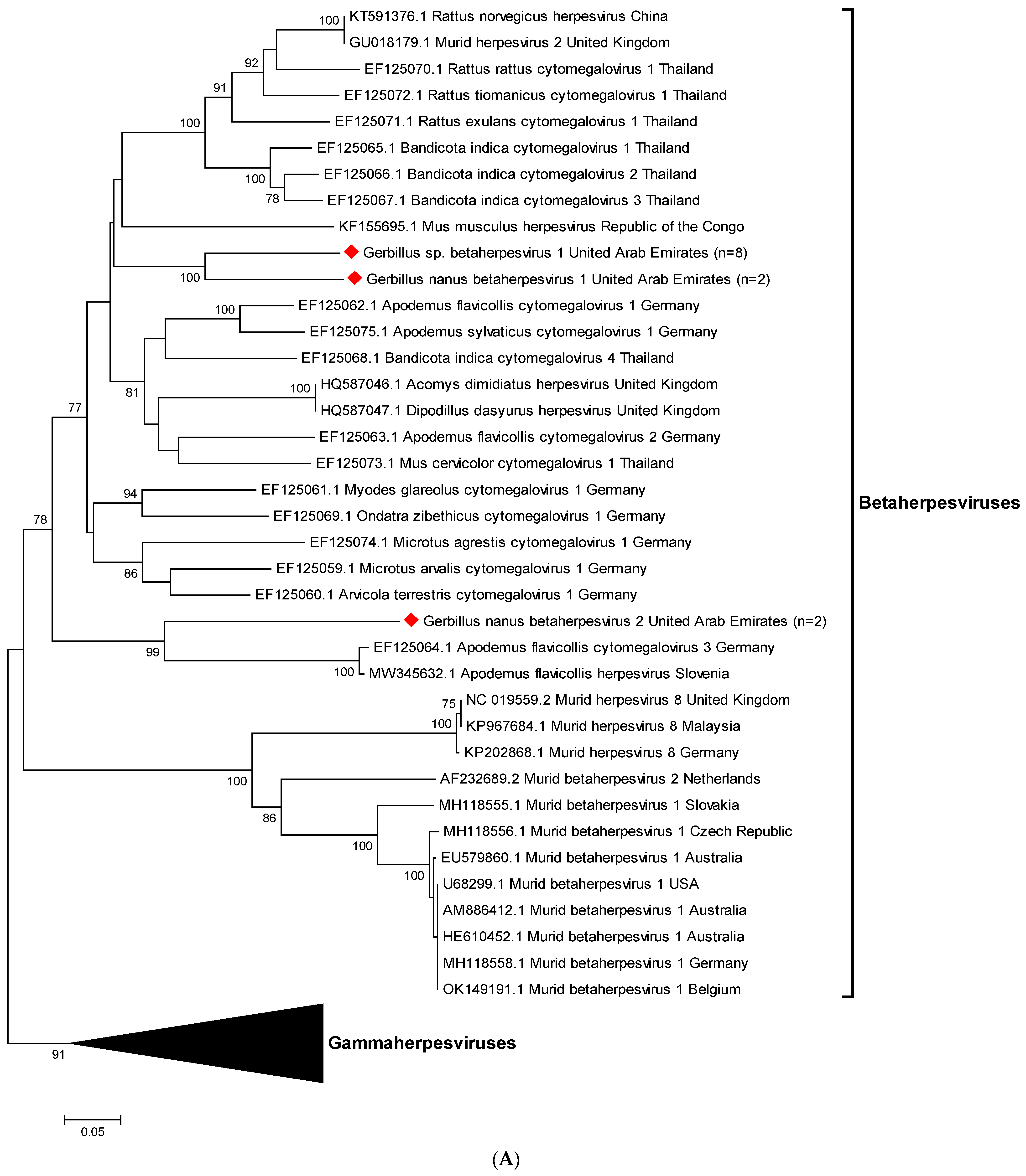
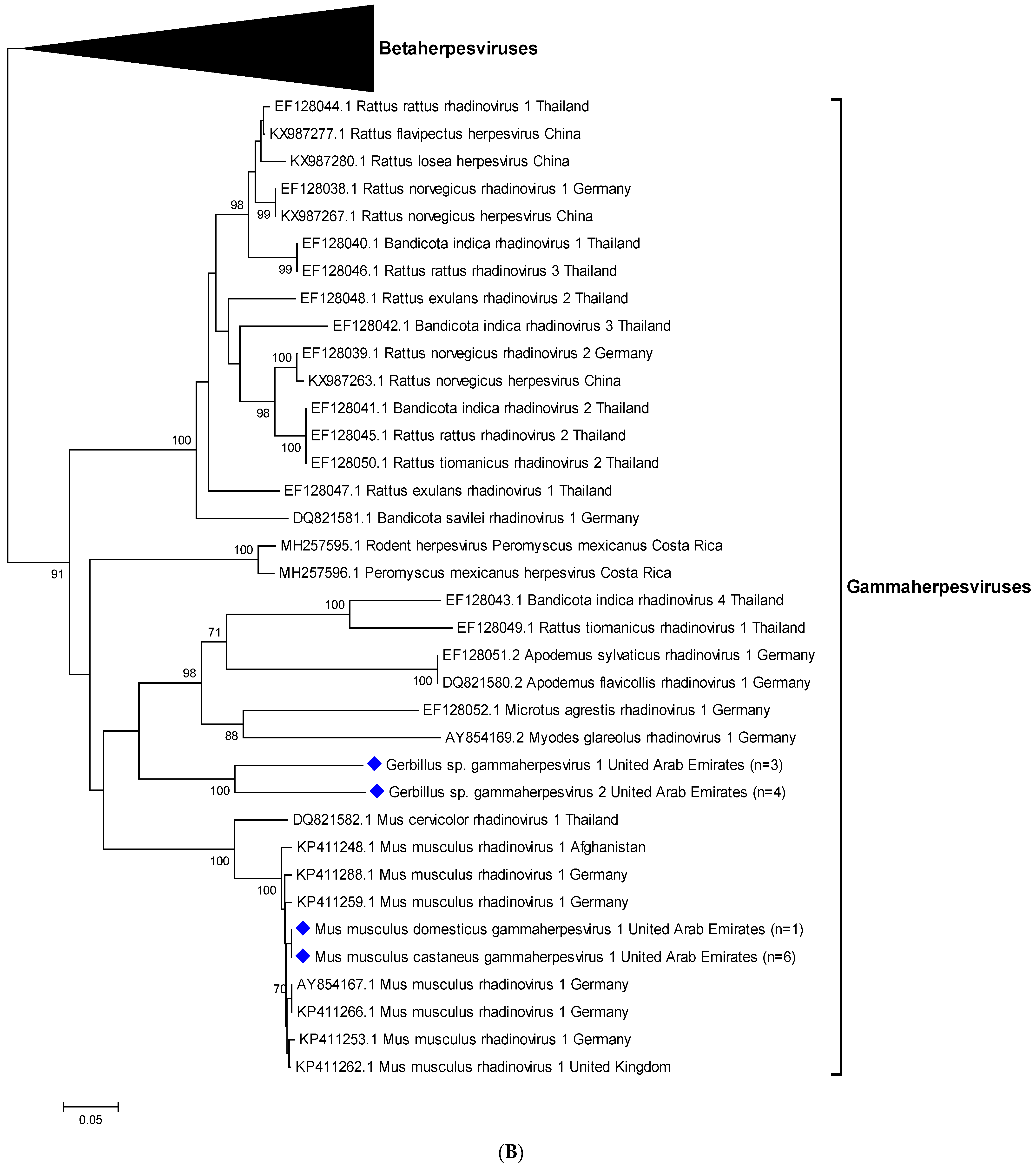
3.3. Sequencing and Phylogenetic Analyses
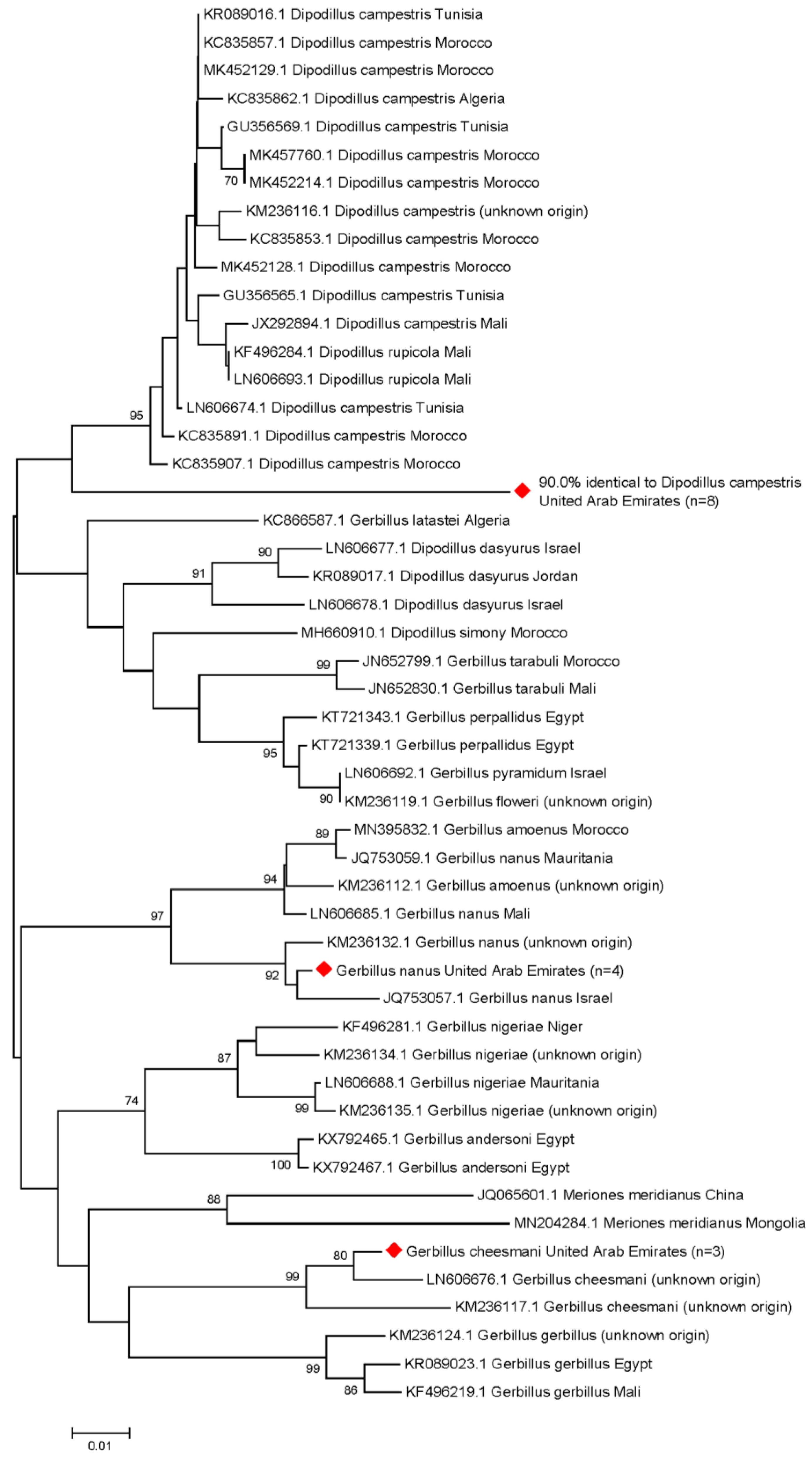
3.4. Molecular Rodent Species Identification
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, Z.; Lu, L.; Du, J.; Yang, L.; Ren, X.; Liu, B.; Jiang, J.; Yang, J.; Dong, J.; Sun, L.; et al. Comparative analysis of rodent and small mammal viromes to better understand the wildlife origin of emerging infectious diseases. Microbiome 2018, 6, 178. [Google Scholar] [CrossRef]
- Wernery, U. Zoonoses in the Arabian Peninsula. Saudi Med. J. 2014, 35, 1455–1462. [Google Scholar] [PubMed]
- Reusken, C.B.; Haagmans, B.L.; Müller, M.A.; Gutierrez, C.; Godeke, G.-J.; Meyer, B.; Muth, D.; Raj, V.S.; Vries, L.S.-D.; Corman, V.M.; et al. Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: A comparative serological study. Lancet Infect. Dis. 2013, 13, 859–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowotny, N.; Kolodziejek, J. Middle East respiratory syndrome coronavirus (MERS-CoV) in dromedary camels, Oman, 2013. Euro Surveill. Bull. Eur. Sur Les Mal. Transm. Eur. Commun. Dis. Bull. 2014, 19, 20781. [Google Scholar] [CrossRef] [Green Version]
- Weidinger, P.; Kolodziejek, J.; Camp, J.V.; Loney, T.; Kannan, D.O.; Ramaswamy, S.; Tayoun, A.A.; Corman, V.M.; Nowotny, N. MERS-CoV in sheep, goats, and cattle, United Arab Emirates, 2019: Virological and serological investigations reveal an accidental spillover from dromedaries. Transbound. Emerg. Dis. 2022, 69, 3066–3072. [Google Scholar] [CrossRef] [PubMed]
- Deyde, V.M.; Khristova, M.L.; Rollin, P.E.; Ksiazek, T.G.; Nichol, S.T. Crimean-Congo hemorrhagic fever virus genomics and global diversity. J. Virol. 2006, 80, 8834–8842. [Google Scholar] [CrossRef] [Green Version]
- Camp, J.V.; Kannan, D.O.; Osman, B.M.; Shah, M.S.; Howarth, B.; Khafaga, T.; Weidinger, P.; Karuvantevida, N.; Kolodziejek, J.; Mazrooei, H.; et al. Crimean-Congo Hemorrhagic Fever Virus Endemicity in United Arab Emirates, 2019. Emerg. Infect. Dis. 2020, 26, 1019–1021. [Google Scholar] [CrossRef]
- Shahhosseini, N.; Wong, G.; Babuadze, G.; Camp, J.V.; Ergonul, O.; Kobinger, G.P.; Chinikar, S.; Nowotny, N. Crimean-Congo hemorrhagic fever virus in Asia, Africa and Europe. Microorganisms 2021, 9, 1907. [Google Scholar] [CrossRef]
- Camp, J.V.; Weidinger, P.; Ramaswamy, S.; Kannan, D.O.; Osman, B.M.; Kolodziejek, J.; Karuvantevida, N.; Abou Tayoun, A.; Loney, T.; Nowotny, N. Association of dromedary camels and camel ticks with reassortant Crimean-Congo hemorrhagic fever virus, United Arab Emirates. Emerg. Infect. Dis. 2021, 27, 2471–2474. [Google Scholar] [CrossRef]
- Madani, T.A.; Abuelzein, E.-T.M.E. Alkhumra hemorrhagic fever virus infection. Arch. Virol. 2021, 166, 2357–2367. [Google Scholar] [CrossRef]
- Zakham, F.; Albalawi, A.E.; Alanazi, A.D.; Truong Nguyen, P.; Alouffi, A.S.; Alaoui, A.; Sironen, T.; Smura, T.; Vapalahti, O. Viral RNA metagenomics of Hyalomma ticks collected from dromedary camels in Makkah province, Saudi Arabia. Viruses 2021, 13, 1396. [Google Scholar] [CrossRef]
- Charrel, R.N.; Zaki, A.M.; Fakeeh, M.; Yousef, A.I.; de Chesse, R.; Attoui, H.; de Lamballerie, X. Low diversity of Alkhurma hemorrhagic fever virus, Saudi Arabia, 1994–1999. Emerg. Infect. Dis. 2005, 11, 683–688. [Google Scholar] [CrossRef]
- Simmons, J.H.; Riley, L.K. Hantaviruses: An overview. Comp. Med. 2002, 52, 97–110. [Google Scholar] [PubMed]
- Singh, S.; Numan, A.; Sharma, D.; Shukla, R.; Alexander, A.; Jain, G.K.; Ahmad, F.J.; Kesharwani, P. Epidemiology, virology and clinical aspects of hantavirus infections: An overview. Int. J. Environ. Health Res. 2022, 32, 1815–1826. [Google Scholar] [CrossRef]
- Fornůsková, A.; Hiadlovská, Z.; Macholán, M.; Piálek, J.; de Bellocq, J.G. New perspective on the geographic distribution and evolution of lymphocytic choriomeningitis virus, Central Europe. Emerg. Infect. Dis. 2021, 27, 2638–2647. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.J.; Paskey, A.C.; Ebinger, A.; Pfaff, F.; Priemer, G.; Höper, D.; Breithaupt, A.; Heuser, E.; Ulrich, R.G.; Kuhn, J.H.; et al. Relatives of rubella virus in diverse mammals. Nature 2020, 586, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Matiasek, K.; Pfaff, F.; Weissenböck, H.; Wylezich, C.; Kolodziejek, J.; Tengstrand, S.; Ecke, F.; Nippert, S.; Starcky, P.; Litz, B.; et al. Mystery of fatal ‘staggering disease’ unravelled: Novel rustrela virus causes severe encephalomyelitis in domestic cats. Nat. Commun. 2023, 14, 624. [Google Scholar] [CrossRef] [PubMed]
- Kronevi, T.; Nordström, M.; Moreno, W.; Nilsson, P.O. Feline ataxia due to nonsuppurative meningoencephalomyelitis of unknown aetiology. Nord. Vet. 1974, 26, 720–725. [Google Scholar]
- Nowotny, N.; Weissenböck, H. Description of feline nonsuppurative meningoencephalomyelitis (“staggering disease”) and studies of its etiology. J. Clin. Microbiol. 1995, 33, 1668–1669. [Google Scholar] [CrossRef] [Green Version]
- Mitjà, O.; Ogoina, D.; Titanji, B.K.; Galvan, C.; Muyembe, J.-J.; Marks, M.; Orkin, C.M. Monkeypox. Lancet Lond. Engl. 2023, 401, 60–74. [Google Scholar] [CrossRef]
- Joseph, S.; Kinne, J.; Nagy, P.; Juhász, J.; Barua, R.; Patteril, N.A.G.; Hoffmann, D.; Pfaff, F.; Hoffmann, B.; Wernery, U. Outbreak of a systemic form of camelpox in a dromedary herd (Camelus dromedarius) in the United Arab Emirates. Viruses 2021, 13, 1940. [Google Scholar] [CrossRef]
- Al-Salam, S.; Nowotny, N.; Sohail, M.R.; Kolodziejek, J.; Berger, T.G. Ecthyma contagiosum (orf)–report of a human case from the United Arab Emirates and review of the literature. J. Cutan. Pathol. 2008, 35, 603–607. [Google Scholar] [CrossRef]
- Hawranek, T.; Tritscher, M.; Muss, W.H.; Jecel, J.; Nowotny, N.; Kolodziejek, J.; Emberger, M.; Schaeppi, H.; Hintner, H. Feline orthopoxvirus infection transmitted from cat to human. J. Am. Acad. Dermatol. 2003, 49, 513–518. [Google Scholar] [CrossRef]
- Carroll, D.S.; Emerson, G.L.; Li, Y.; Sammons, S.; Olson, V.; Frace, M.; Nakazawa, Y.; Czerny, C.P.; Tryland, M.; Kolodziejek, J.; et al. Chasing Jenner’s vaccine: Revisiting cowpox virus classification. PLoS ONE 2011, 6, e23086. [Google Scholar] [CrossRef] [Green Version]
- Kuno, G.; Chang, G.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolodziejek, J.; Marinov, M.; Kiss, B.J.; Alexe, V.; Nowotny, N. The complete sequence of a West Nile virus lineage 2 strain detected in a Hyalomma marginatum marginatum tick collected from a song thrush (Turdus philomelos) in eastern Romania in 2013 revealed closest genetic relationship to strain Volgograd 2007. PLoS ONE 2014, 9, e109905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakonyi, T.; Ivanics, E.; Erdelyi, K.; Ursu, K.; Ferenczi, E.; Weissenböck, H.; Nowotny, N. Lineage 1 and 2 strains of encephalitic West Nile virus, Central Europe. Emerg. Infect. Dis. 2006, 12, 618–623. [Google Scholar] [CrossRef]
- Pollock, C.G. West Nile virus in the Americas. J. Avian. Med. Surg. 2008, 22, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.; Wernery, U.; Teng, J.L.; Wernery, R.; Huang, Y.; Patteril, N.A.; Chan, K.-H.; Elizabeth, S.K.; Fan, R.Y.; Lau, S.K.; et al. First isolation of West Nile virus from a dromedary camel. Emerg. Microbes Infect. 2016, 5, e53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camp, J.V.; Karuvantevida, N.; Chouhna, H.; Safi, E.; Shah, J.N.; Nowotny, N. Mosquito biodiversity and mosquito-borne viruses in the United Arab Emirates. Parasites Vectors 2019, 12, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, M. Herpesviruses: A brief overview. Methods Mol. Biol. Clifton N.J. 2004, 256, 199–219. [Google Scholar]
- Al-Salihi, K.A.; Khalaf, J.M. The emerging SARS-CoV, MERS-CoV, and SARS-CoV-2: An insight into the viruses zoonotic aspects. Vet. World 2021, 14, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Han, H.-J.; Wen, H.; Zhou, C.-M.; Chen, F.-F.; Luo, L.-M.; Liu, J.; Yu, X.-J. Bats as reservoirs of severe emerging infectious diseases. Virus Res. 2015, 205, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Khafaga, T.; Bell, S. Rodent Community Structure and Dynamics in Dubai Desert Conservation Reserve. 2012, 143p. Available online: https://www.ddcr.org/reports/full/Rodents-community-Structure-and-dynamics.pdf (accessed on 12 February 2023).
- McGrath, D. Are Desert Rodents a Suitable Indicator for Assessing Change in Ecosystem Health in the United Arab Emirate’s Inland Deserts? University of Oxford: Oxford, UK, 2018. [Google Scholar]
- Corman, V.M.; Müller, M.A.; Costabel, U.; Timm, J.; Binger, T.; Meyer, B.; Kreher, P.; Lattwein, E.; Eschbach-Bludau, M.; Nitsche, A.; et al. Assays for laboratory confirmation of novel human coronavirus (hCoV-EMC) infections. Eurosurveillance 2012, 17, 20334. [Google Scholar] [CrossRef] [Green Version]
- Klempa, B.; Fichet-Calvet, E.; Lecompte, E.; Auste, B.; Aniskin, V.; Meisel, H.; Denys, C.; Koivogui, L.; ter Meulen, J.; Krüger, D.H. Hantavirus in African wood mouse, Guinea. Emerg. Infect. Dis. 2006, 12, 838–840. [Google Scholar] [CrossRef]
- Li, Y.; Meyer, H.; Zhao, H.; Damon, I.K. GC content-based pan-pox universal PCR assays for poxvirus detection. J. Clin. Microbiol. 2010, 48, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.; Landt, O.; Kaiser, M.; Faye, O.; Koppe, T.; Lass, U.; Sall, A.A.; Niedrig, M. Development of one-step quantitative reverse transcription PCR for the rapid detection of flaviviruses. Virol. J. 2013, 10, 58. [Google Scholar] [CrossRef] [Green Version]
- VanDevanter, D.R.; Warrener, P.; Bennett, L.; Schultz, E.R.; Coulter, S.; Garber, R.L.; Rose, T.M. Detection and analysis of diverse herpesviral species by consensus primer PCR. J. Clin. Microbiol. 1996, 34, 1666–1671. [Google Scholar] [CrossRef] [Green Version]
- Kitano, T.; Umetsu, K.; Tian, W.; Osawa, M. Two universal primer sets for species identification among vertebrates. Int. J. Leg. Med. 2007, 121, 423–427. [Google Scholar] [CrossRef]
- Ndiaye, A.; Tatard, C.; Stanley, W.; Granjon, L. Taxonomic hypotheses regarding the genus Gerbillus (Rodentia, Muridae, Gerbillinae) based on molecular analyses of museum specimens. ZooKeys 2016, 566, 145–155. [Google Scholar]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Herbert, P.D.N. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Granjon, L. Gerbillus campestris. Available online: https://www.iucnredlist.org/species/45088/22465830 (accessed on 28 September 2022).
- Lado, S.; Elbers, J.P.; Plasil, M.; Loney, T.; Weidinger, P.; Camp, J.V.; Kolodziejek, J.; Futas, J.; Kannan, D.A.; Orozco-terWengel, P.; et al. Innate and adaptive immune genes associated with MERS-CoV infection in dromedaries. Cells 2021, 10, 1291. [Google Scholar] [CrossRef] [PubMed]
- Lado, S.; Futas, J.; Plasil, M.; Loney, T.; Weidinger, P.; Camp, J.V.; Kolodziejek, J.; Kannan, D.O.; Horin, P.; Nowotny, N.; et al. Crimean–Congo hemorrhagic fever virus past infections are associated with two innate immune response candidate genes in dromedaries. Cells 2021, 11, 8. [Google Scholar] [CrossRef]
- Gargili, A.; Estrada-Pena, A.; Spengler, J.R.; Lukashev, A.; Nuttall, P.A.; Bente, D.A. The role of ticks in the maintenance and transmission of Crimean-Congo hemorrhagic fever virus: A review of published field and laboratory studies. Antivir. Res. 2017, 144, 93–119. [Google Scholar] [CrossRef]
- Mohd, H.A.; Al-Tawfiq, J.A.; Memish, Z.A. Middle East respiratory syndrome coronavirus (MERS-CoV) origin and animal reservoir. Virol. J. 2016, 13, 87. [Google Scholar] [CrossRef] [Green Version]
- Corman, V.M.; Ithete, N.L.; Richards, L.R.; Schoeman, M.C.; Preiser, W.; Drosten, C.; Drexler, J.F. Rooting the phylogenetic tree of Middle East respiratory syndrome coronavirus by characterization of a conspecific virus from an African bat. J. Virol. 2014, 88, 11297–11303. [Google Scholar] [CrossRef] [Green Version]
- Dudas, G.; Carvalho, L.M.; Rambaut, A.; Bedford, T. MERS-CoV spillover at the camel-human interface. eLife 2018, 7, e31257. [Google Scholar] [CrossRef]
- Hemida, M.G.; Alhammadi, M.; Almathen, F.; Alnaeem, A. Exploring the potential roles of some rodents in the transmission of the Middle East respiratory syndrome coronavirus. J. Med. Virol. 2021, 93, 5328–5332. [Google Scholar] [CrossRef]
- Hemida, M.G.; Alhammadi, M.; Almathen, F.; Alnaeem, A. Lack of detection of the Middle East respiratory syndrome coronavirus (MERS-CoV) nucleic acids in some Hyalomma dromedarii infesting some Camelus dromedary naturally infected with MERS-CoV. BMC Res. Notes 2021, 14, 96. [Google Scholar] [CrossRef]
- Kashuba, C.; Hsu, C.; Krogstad, A.; Franklin, C. Small mammal virology. Vet. Clin. N. America. Exot. Anim. Pract. 2005, 8, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Yama, I.N.; Cazaux, B.; Britton-Davidian, J.; Moureau, G.; Thirion, L.; de Lamballerie, X.; Dobigny, G.; Charrel, R.N. Isolation and characterization of a new strain of lymphocytic choriomeningitis virus from rodents in southwestern France. Vector Borne Zoonotic Dis. Larchmt. N. Y. 2012, 12, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T. Flavivirus encephalitis. N. Engl. J. Med. 2004, 351, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Nawa, M. Development of a new cell system for the infectivity assay of dengue viruses: Plaque formation and virus growth of prototype and wild-type dengue virus strains in a newly established cell line, GK. Microbiol. Immunol. 1984, 28, 765–776. [Google Scholar] [CrossRef]
- Süss, J.; Béziat, P.; Ramelow, C.; Kahl, O. Tick-borne encephalitis virus (TBEV)-specific RT-PCR for characterization of natural foci of TBE and for other applications. Zent. Bakteriol. 1997, 286, 125–138. [Google Scholar] [CrossRef]
- Ehlers, B.; Küchler, J.; Yasmum, N.; Dural, G.; Voigt, S.; Schmidt-Chanasit, J.; Jäkel, T.; Matuschka, F.-R.; Richter, D.; Essbauer, S.; et al. Identification of novel rodent herpesviruses, including the first gammaherpesvirus of Mus musculus. J. Virol. 2007, 81, 8091–8100. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Virus (Family) | Primer/Probe Name | Sequence (5′-3′) | Amplicon Length | PCR Kit/Conditions | Reference | |
|---|---|---|---|---|---|---|
| MERS-CoV (Coronaviridae) | Orf1a-F Orf1a-R Orf1a-P | CCACTACTCCCATTTCGTCAG CAGTATGTGTAGTGCGCATATAAGCA TTGCAAATTGGCTTGCCCCCACT | 83 bp | Quantabio qScript XLT 1-Step RT-qPCR ToughMix | RT-qPCR; Corman et al. [37] | |
| CCHFV (Nairoviridae) | - | Data not provided by manufacturer | - | 50 °C for 10 min; 95 °C for 2 min; 45x 95 °C for 15 s, 55 °C for 45 s, and 72 °C for 15 s | RealStar RT-PCR Kit 1.0; altona Diagnostics | |
| AHFV (Flaviviridae) | NS3-5439F NS3-5565R NS3-5474P | CAGGGGAGACAGAATTGGGAAG TCATGAGCACCAAAGCGCAC TGAAGCCCATTGGACCGATCCACATAGCAT | 84 bp | Quantabio qScript XLT 1-Step RT-qPCR ToughMix | RT-qPCR; in-house method | |
| Hantaviruses (Hantaviridae) | HAN-L-F1 HAN-L-R1 | ATGTAYGTBAGTGCWGATGC AACCADTCWGTYCCRTCATC | 452 bp | QIAGEN OneStep RT-PCR Kit | Nested RT-PCR; Klempa et al. [38] | |
| HAN-L-F2 HAN-L-R2 | TGCWGATGCHACIAARTGGTC GCRTCRTCWGARTGRTGDGCAA | 390 bp | QIAGEN OneStep RT-PCR Kit: 50 °C for 1 min; 95 °C for 1 min; 40x 94 °C for 30 s, 55 °C for 30 s and 72 °C for 30 s; 72 °C for 7 min | |||
| LCMV (Arenaviridae) | LCM-S-F LCM-S-R | CTGTGAGYGCYTGCACAACATC GATCCTAGGCATTTGATTGCGC | 650 bp | QIAGEN OneStep RT-PCR Kit | RT-PCR; in-house method | |
| RusV (Matonaviridae) | RusV-234F RusV-323R RusV-256P | CCCCGTGTTCCTAGGCAC TCGCCCCATTCWACCCAATT GTGAGCGACCACCCAGCACTCCA | 51 bp | Quantabio qScript XLT 1-Step RT-qPCR ToughMix | RT-qPCR; Matiasek et al. [17] | |
| Poxviruses (Poxviridae) | Pan-Pox-F Pan-Pox-R | ACACCAAAAACTCATATAACTTCT CCTATTTTACTCCTTAGTAAATGAT | 220 bp | QIAGEN Fast Cycling PCR Kit: 95 °C for 5 min; 50x 96 °C for 5 s, 50 °C for 5 s, 68 °C for 10 s; 68 °C for 1 min | Pan-Pox PCR (low GC); Li et al. [39] | |
| Flaviviruses (Flaviviridae) | Flavi-S Flavi-AS2 | TACAACATGATGGGGAARAGAGARAA GTGTCCCAGCCNGCKGTGTCATCWGC | 260 bp | QIAGEN OneStep RT-PCR Kit | Universal RT-PCR; Patel et al. [40] | |
| Herpesviruses (Herpesviridae) | HerpesCons-F1 HerpesCons-F2 HerpesCons-R | GAYTTYGCNAGYYTNTAYCC TCCTGGACAAGCAGCARNYSGCNMTNAA GTCTTGCTCACCAGNTCNACNCCYTT | 700 bp | QIAGEN Fast Cycling PCR Kit: 95 °C for 5 min; 50x 96 °C for 5 s, 50 °C for 5 s, 68 °C for 25 s; 72 °C for 1 min | Nested PCR; VanDevanter et al. [41] | |
| HerpesCons-nF HerpesCons-nR | TGTAACTCGGTGTAYGGNTTYACNGGNGT CACAGAGTCCGTRTCNCCRTADAT | 215–315 bp | QIAGEN Fast Cycling PCR Kit: 95 °C for 5 min; 50x 96 °C for 5 s, 50 °C for 5 s, 68 °C for 8 s; 72 °C for 1 min |
| Primer Name | Sequence (5′-3′) | Amplicon Length | PCR Conditions | Reference |
|---|---|---|---|---|
| L2513-F H2714-R | GCCTGTTTACCAAAAACATCAC CTCCATAGGGTCTTCTCGTCTT | 243 bp | 95 °C for 5 min; 50x 96 °C for 5 s, 57 °C for 5 s, 68 °C for 15 s; 68 °C for 1 min | Kitano et al. [42] |
| GERBCYTB-F2 GERBCYTB-R3 | GCAAACGGAGCCTCAATATT CATTCTACRATTGTTGGGCCA | 239 bp | 95 °C for 5 min; 50x 96 °C for 5 s, 55 °C for 5 s, 68 °C for 30 s; 72 °C for 1 min | Ndiaye et al. [43] |
| VF1d VR1d | TGTAAAACGACGGCCAGTTYT-CNACHAAYCAYAAAGAYATYGG CAGGAAACAGCTATGACTANA-CYTCNGGRTGNCCRAARAATCA | 659 bp | 95 °C for 2 min; 5x 96 °C for 10 s, 50 °C for 10 s, 68 °C for 30 s; 35x 96 °C for 10 s, 54 °C for 10 s, 68 °C for 30 s; 72 °C for 1 min | Modified from Ivanova et al. [44] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weidinger, P.; Kolodziejek, J.; Khafaga, T.; Loney, T.; Howarth, B.; Sher Shah, M.; Abou Tayoun, A.; Alsheikh-Ali, A.; Camp, J.V.; Nowotny, N. Potentially Zoonotic Viruses in Wild Rodents, United Arab Emirates, 2019—A Pilot Study. Viruses 2023, 15, 695. https://doi.org/10.3390/v15030695
Weidinger P, Kolodziejek J, Khafaga T, Loney T, Howarth B, Sher Shah M, Abou Tayoun A, Alsheikh-Ali A, Camp JV, Nowotny N. Potentially Zoonotic Viruses in Wild Rodents, United Arab Emirates, 2019—A Pilot Study. Viruses. 2023; 15(3):695. https://doi.org/10.3390/v15030695
Chicago/Turabian StyleWeidinger, Pia, Jolanta Kolodziejek, Tamer Khafaga, Tom Loney, Brigitte Howarth, Moayyed Sher Shah, Ahmad Abou Tayoun, Alawi Alsheikh-Ali, Jeremy V. Camp, and Norbert Nowotny. 2023. "Potentially Zoonotic Viruses in Wild Rodents, United Arab Emirates, 2019—A Pilot Study" Viruses 15, no. 3: 695. https://doi.org/10.3390/v15030695