
Insights into the Molecular Epidemiology of Foot-and-Mouth Disease Virus in Russia, Kazakhstan, and Mongolia in Terms of O/ME-SA/Ind-2001e Sublineage Expansion

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
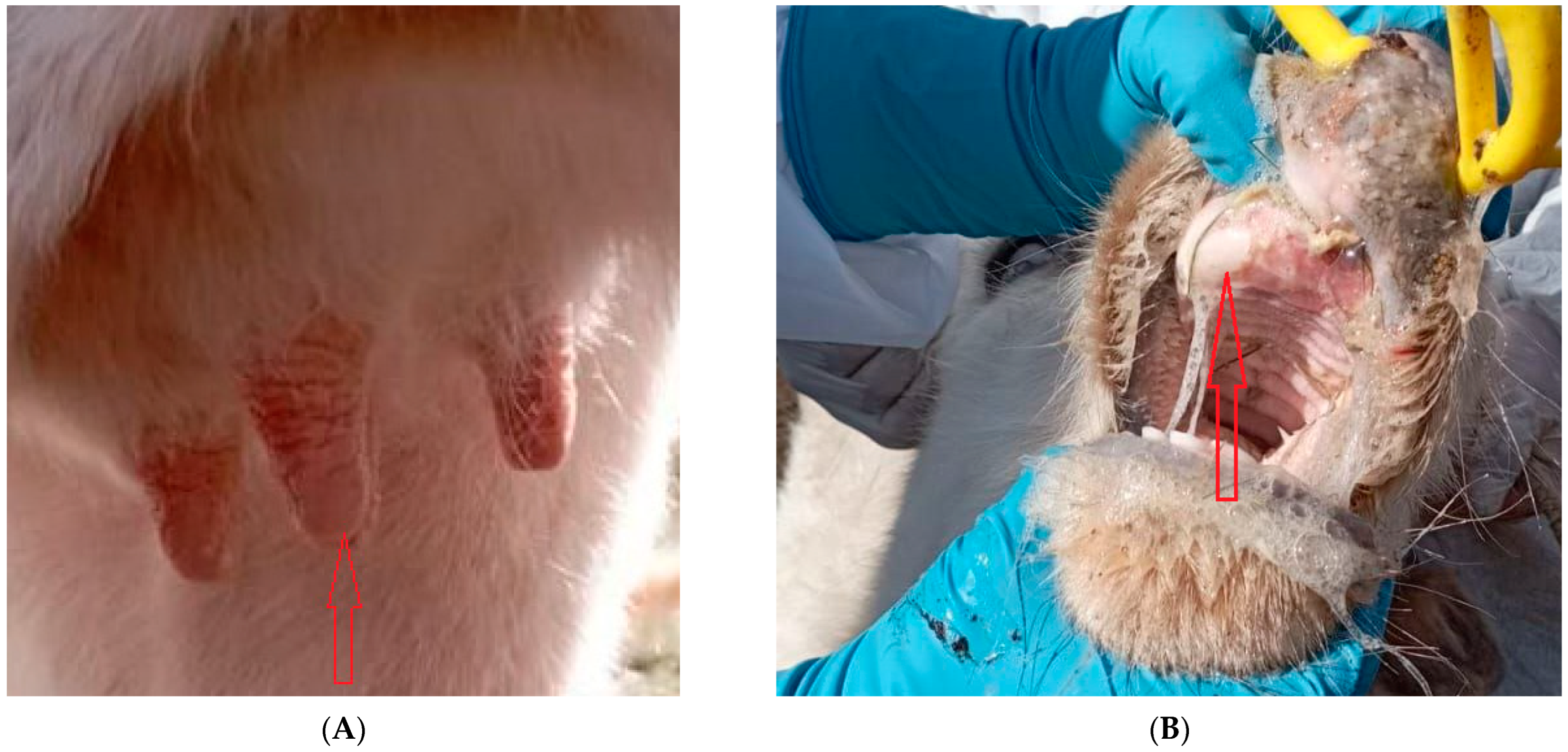
2.1. Sampling and Control Measure Taken during the Outbreak
2.2. Data Collection
2.3. RNA Extraction
2.4. PCR and Sequencing
2.5. Virus Isolation
2.6. Vaccine Matching
3. Results
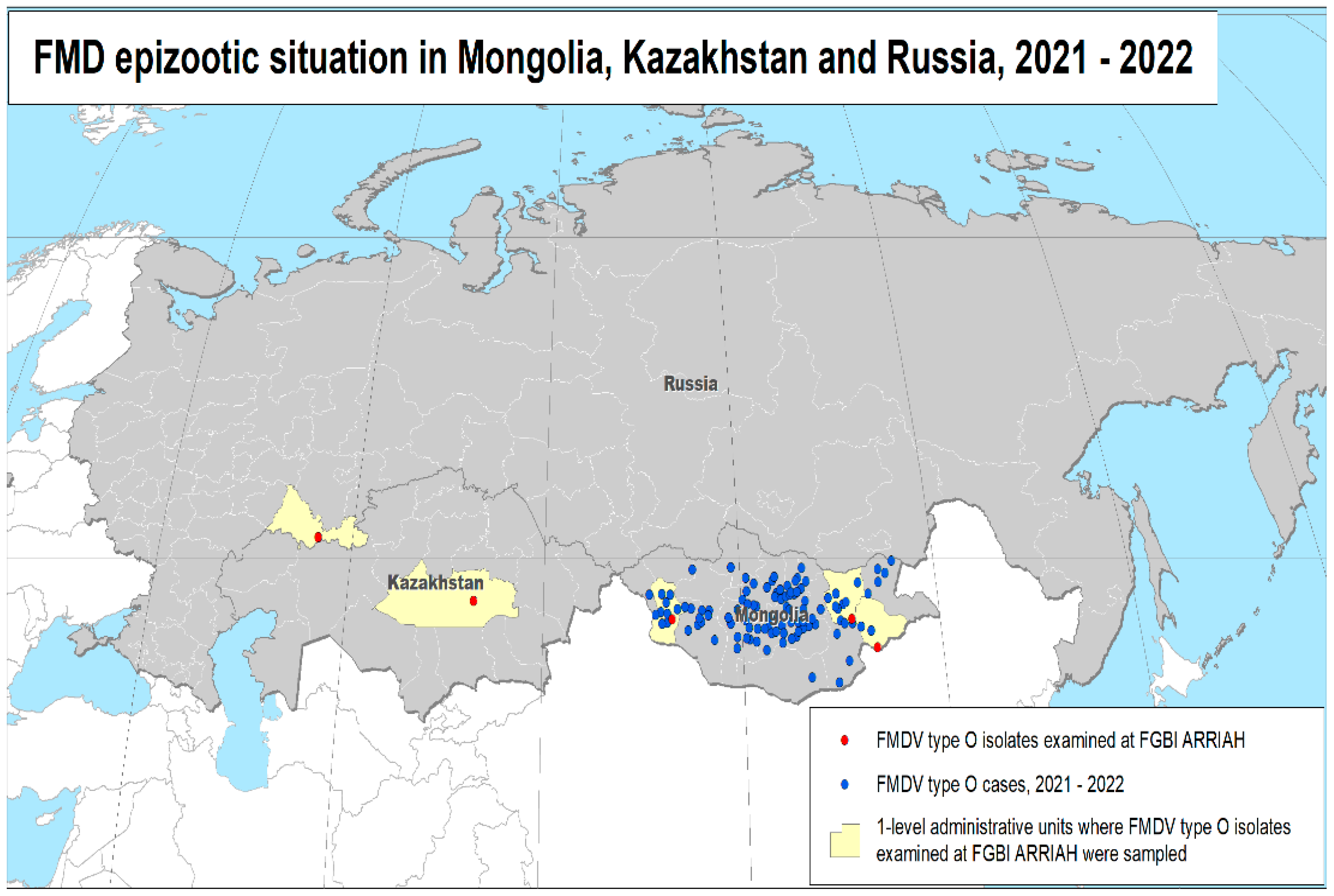
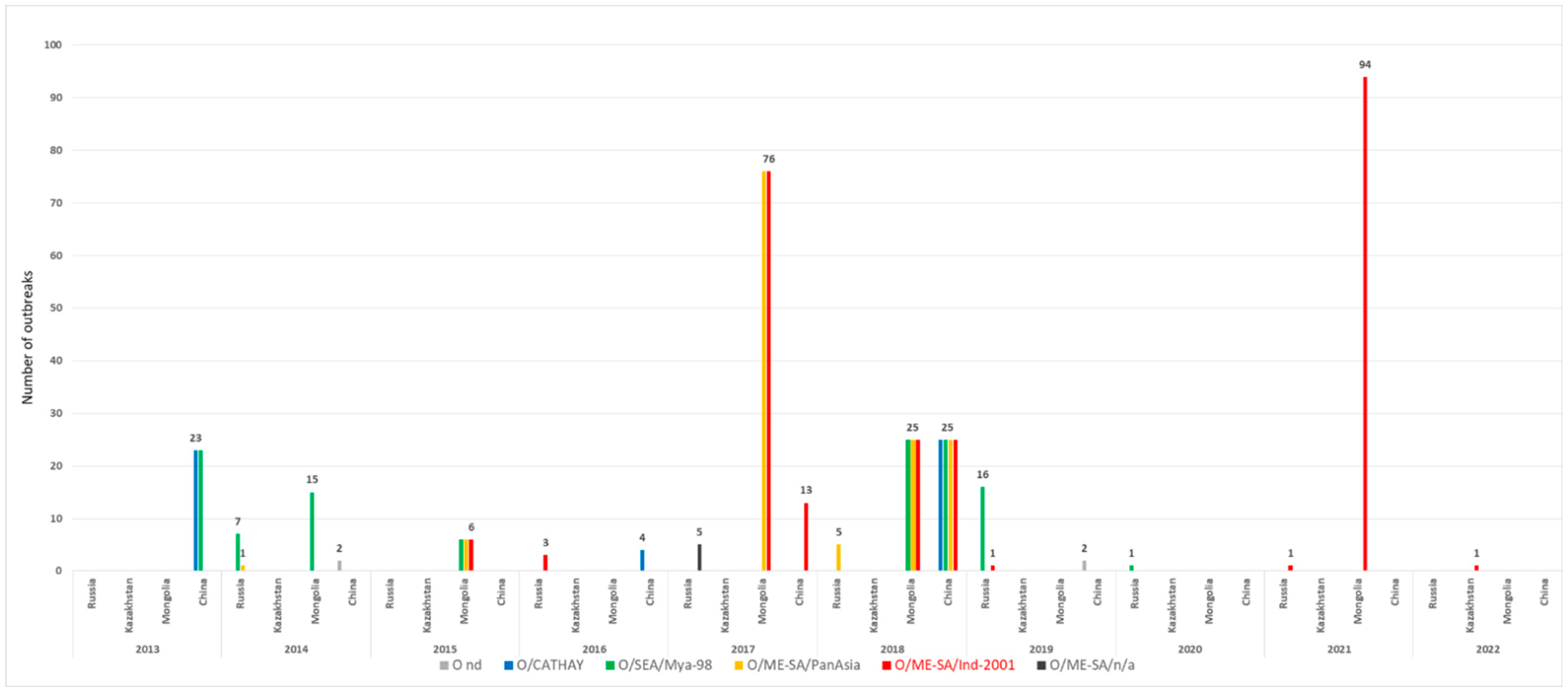
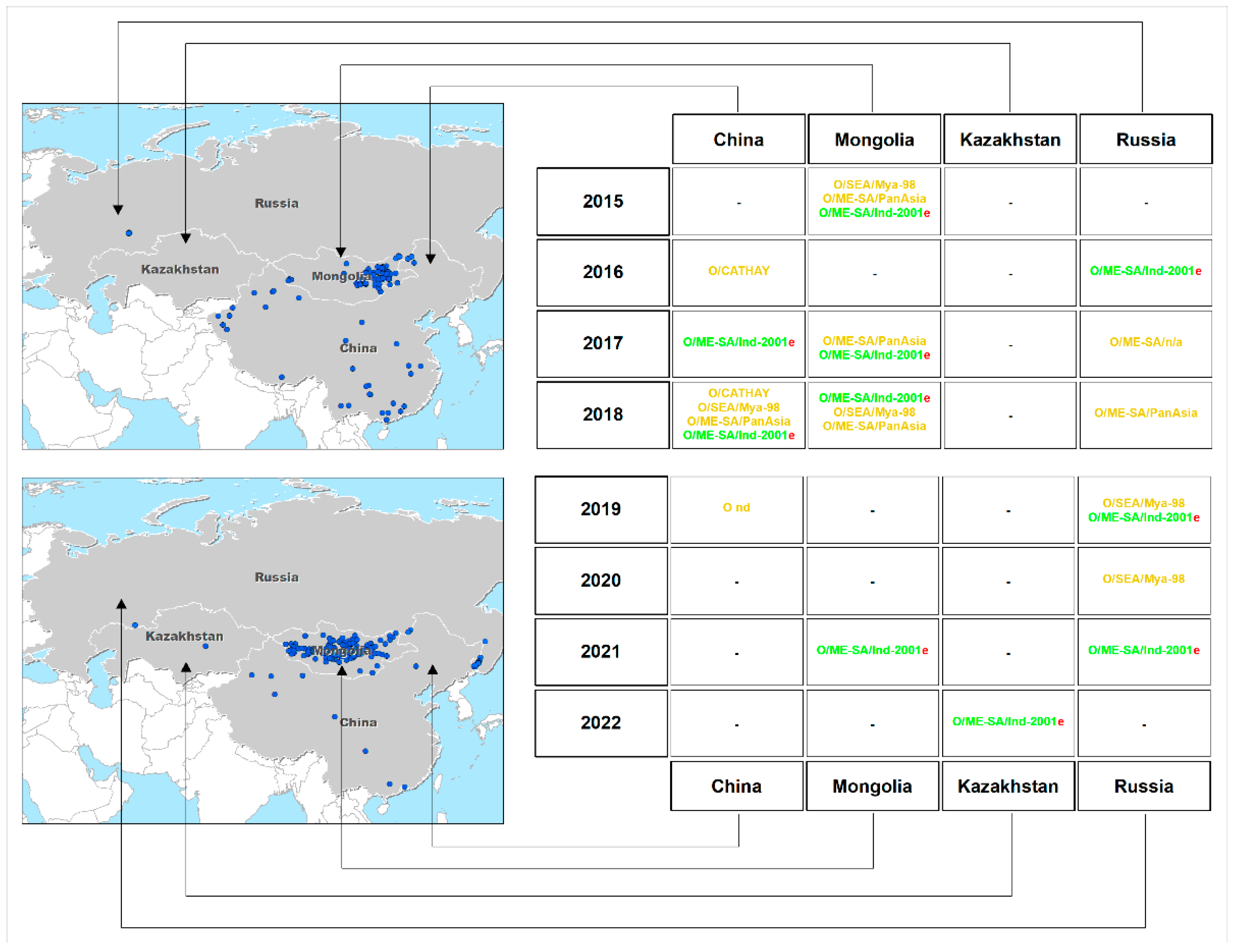
3.1. Time Course of FMD Outbreaks
3.2. VP1 Sequencing
3.3. Vaccine Matching
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jamal, S.M.; Belsham, G.J. Foot-and-mouth disease: Past, present and future. Vet. Res. 2013, 44, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grubman, M.J.; Baxt, B. Foot-and-mouth disease. Clin. Microbiol. Rev. 2004, 17, 465–493. [Google Scholar] [CrossRef] [Green Version]
- Mason, P.W.; Grubman, M.J.; Baxt, B. Molecular basis of pathogenesis of FMDV. Virus Res. 2003, 91, 9–32. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.A.; Hernández, J.; Piccone, M.E.; Palma, E.L.; Domingo, E.; Knowles, N.; Mateu, M.G. Two mechanisms of antigenic diversification of foot-and-mouth disease virus. Virology 1991, 184, 695–706. [Google Scholar] [CrossRef]
- Singh, C.P.; Verma, A.K.; Pal, B.C. Prevalence of protected animals against foot and mouth disease in Uttar Pradesh. Haryana Vet. 2008, 47, 107–109. [Google Scholar]
- Singh, R.; Pandey, A.B.; Chandra, D.K.; Singh, P.; Mehrortra, M.L. Epidemiology of malignant form of foot-and-mouth disease in susceptible cattle and buffalo population of Punjab and Uttar Pradesh. Indian J. Anim. Sci. 2008, 3–7, 78. [Google Scholar]
- Hegde, R.; Kumar, N.; Prabhakaran, K.; Wadhwan, V.M.; Kakker, N.; Lokhande, T.; Sharma, K.; Kanani, A.; Limaye, K.N.; Pn, A.; et al. Foot and Mouth Disease (FMD) incidence in cattle and buffaloes and its associated farm-level economic costs in endemic India. Prev. Vet. Med. 2021, 190, 105318. [Google Scholar]
- Carrillo, C.; Tulman, E.R.; Delhon, G.; Lu, Z.; Carreno, A.; Vagnozzi, A.; Kutish, G.F.; Rock, D.L. Comparative genomics of foot-and-mouth disease virus. J. Virol. 2005, 79, 6487–6504. [Google Scholar] [CrossRef] [Green Version]
- Brito, B.P.; Perez, A.M.; Capozzo, A.V. Accuracy of traditional and novel serology tests for predicting cross-protection in foot-and-mouth disease vaccinated cattle. Vaccine 2014, 32, 433–436. [Google Scholar] [CrossRef]
- Cao, Y.; Lu, Z.; Li, D.; Fan, P.; Sun, P.; Bao, H.; Fu, Y.; Li, P.; Bai, X.; Chen, Y.; et al. Evaluation of cross-protection against three topotypes of serotype O foot-and-mouth disease virus in pigs vaccinated with multi-epitope protein vaccine incorporated with poly(I:C). Vet. Microbiol. 2014, 168, 294–301. [Google Scholar] [CrossRef]
- Kitching, R.P.; Knowles, N.J.; Samuel, A.R.; Donaldson, A.I. Development of foot-and-mouth disease virus strain characterization—A review. Trop. Anim. Health Prod. 1989, 21, 153–166. [Google Scholar] [CrossRef]
- Samuel, A.R.; Knowles, N.J. Foot-and-mouth disease type O viruses exhibit genetically and geographically distinct evolutionary lineages (topotypes). J. Gen. Virol. 2001, 82, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Hamid, N.F.; Hussein, N.M.; Wadsworth, J.; Radford, A.D.; Knowles, N.J.; King, D.P. Phylogeography of foot-and-mouth disease virus types O and A in Malaysia and surrounding countries. Infect. Genet. Evol. 2011, 11, 320–328. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho Ferreira, H.C.; Pauszek, S.J.; Ludi, A.; Huston, C.L.; Pacheco, J.M.; Le, V.T.; Nguyen, P.T.; Bui, H.H.; Nguyen, T.D.; Nguyen, T.; et al. An integrative analysis of foot-and-mouth disease virus carriers in Vietnam achieved through targeted surveillance and molecular epidemiology. Transbound. Emerg. Dis. 2017, 64, 547–563. [Google Scholar] [CrossRef] [PubMed]
- Brito, B.; Pauszek, S.J.; Eschbaumer, M.; Stenfeldt, C.; de Carvalho Ferreira, H.C.; Vu, L.T.; Phuong, N.T.; Hoang, B.H.; Tho, N.D.; Dong, P.V.; et al. Phylodynamics of foot-and-mouth disease virus O/PanAsia in Vietnam 2010–2014. Vet. Res. 2017, 48, 24. [Google Scholar] [CrossRef] [Green Version]
- Hemadri, D.; Tosh, C.; Sanyal, A.; Venkataramanan, R. Emergence of a new strain of type O foot-and-mouth disease virus: Its phylogenetic and evolutionary relationship with the PanAsia pandemic strain. Virus Genes 2002, 25, 23–34. [Google Scholar] [CrossRef]
- Knowles, N.J.; Bachanek-Bankowska, K.; Wadsworth, J.; Mioulet, V.; Valdazo-González, B.; Eldaghayes, I.M.; Dayhum, A.S.; Kammon, A.M.; Sharif, M.A.; Waight, S.; et al. Outbreaks of foot-and-mouth disease in Libya and Saudi Arabia during 2013 due to an exotic O/ME-SA/Ind-2001 lineage virus. Transbound. Emerg. Dis. 2016, 63, e431–e435. [Google Scholar] [CrossRef] [Green Version]
- WRLFMD. Genotyping Report, Serotype O (Bahrain, 2015). Available online: https://www.wrlfmd.org/fmd_genotyping/2015/WRLFMD-2015-00004%20O%20Bahrain%202015.pdf (accessed on 19 August 2022).
- Valdazo-González, B.; Knowles, N.J.; King, D.P. Genome sequences of foot-and-mouth disease virus O/ME-SA/Ind-2001 lineage from outbreaks in Libya, Saudi Arabia, and Bhutan during 2013. Genome Announc. 2013, 2, e00242-14. [Google Scholar] [CrossRef] [Green Version]
- Bachanek-Bankowska, K.; Wadsworth, J.; Gray, A.; Abouchoaib, N.; King, D.P.; Knowles, N.J. Genome sequence of foot-and-mouth disease virus Serotype O isolated from Morocco in 2015. Genome Announc. 2016, 4, e01746-15. [Google Scholar] [CrossRef] [Green Version]
- Bachanek-Bankowska, K.; Di Nardo, A.; Wadsworth, J.; Mioulet, V.; Pezzoni, G.; Grazioli, S.; Brocchi, E.; Kafle, S.C.; Hettiarachchi, R.; Kumarawadu, P.L.; et al. Reconstructing the evolutionary history of pandemic foot-and-mouth disease viruses: The impact of recombination within the emerging O/ME-SA/Ind-2001 lineage. Sci. Rep. 2018, 8, 14693. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Abila, R.; Rodtian, P.; King, D.P.; Knowles, N.J.; Ngo, L.T.; Le, V.T.; Khounsy, S.; Bounma, P.; Lwin, S.; et al. Emergence of an exotic strain of serotype O foot-and-mouth disease virus O/ME-SA/Ind-2001d in South-East Asia in 2015. Transbound. Emerg. Dis. 2018, 65, e104–e112. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, S.; Lee, H.; Lim, D.R.; Lee, J.W.; Bunnary, S.; Tum, S.; Lee, D.S.; Hwang, H.; Jeong, S.; Nah, J.; et al. Identification of the O/ME-SA/Ind-2001e sublineage of foot-and-mouth disease virus in Cambodia. Front. Vet. Sci. 2021, 8, 749966. [Google Scholar] [CrossRef] [PubMed]
- Jamal, S.M.; Khan, S.; Knowles, N.J.; Wadsworth, J.; Hicks, H.M.; Mioulet, V.; Bin-Tarif, A.; Ludi, A.B.; Shah, S.A.A.; Abubakar, M.; et al. Foot-and-mouth disease viruses of the O/ME-SA/Ind-2001e sublineage in Pakistan. Transbound. Emerg. Dis. 2021, 68, 3126–3135. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.wrlfmd.org/western-and-central-asia/russia#panel-7722 (accessed on 30 December 2022).
- WRLFMD. Genotyping Report, Serotype O (Kazakhstan, 2011). Available online: https://www.wrlfmd.org/sites/world/files/quick_media/WRLMEG-2011-00023-KAZ-GTR-O-O_001.pdf (accessed on 23 October 2022).
- WRLFMD. Genotyping Report, Serotype A (Kazakhstan, 2013). Available online: https://www.wrlfmd.org/sites/world/files/quick_media/WRLMEG-2013-00017-KAZ-GTR-O-A_001.pdf (accessed on 23 October 2022).
- Valdazo-González, B.; Polihronova, L.; Alexandrov, T.; Normann, P.; Knowles, N.J.; Hammond, J.M.; Georgiev, G.K.; Özyörük, F.; Sumption, K.J.; Belsham, G.J.; et al. Reconstruction of the transmission history of RNA virus outbreaks using full genome sequences: Foot-and-mouth disease virus in Bulgaria in 2011. PLoS ONE 2012, 7, e49650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE/Food and Agriculture Organization. Foot-And-Mouth Disease Reference Laboratory Network Annual Report. 2021. Available online: https://www//wrlfmd.org/sites/world/files/quick_media/OIEFAO%20FMD%20Ref%20Lab%20Network%20Report%202021.pdf (accessed on 23 October 2022).
- WRLFMD Quarterly Report; October–December 2021. Available online: https://www.wrlfmd.org/sites/world/files/quick_media/OIE-FAO%20FMD%20Ref%20Lab%20Report%20Oct-Dec%202021.pdf (accessed on 23 October 2022).
- Tyulegenov, S.B.; Zhakupbayev, A.; Berdikulov, M.; Karibayev, T.; Yessembekova, G.N.; Sultanov, A.A.; Perez, A.M.; Abdrakhmanov, S.K. Foot-and-mouth disease in Kazakhstan. Transbound. Emerg. Dis. 2022, 69, 1712–1714. [Google Scholar] [CrossRef] [PubMed]
- Knowles, N.J.; Wadsworth, J.; Bachanek-Bankowska, K.; King, D.P. VP1 sequencing protocol for foot and mouth disease virus molecular epidemiology. Rev. Sci. Tech. 2016, 35, 741–755. [Google Scholar] [CrossRef] [Green Version]
- OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, Chapter 3.1.8. 2021. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.01.08_FMD.pdf (accessed on 23 October 2022).
- Rweyemamu, M.M. Antigenic variation in foot-and-mouth disease: Studies based on the virus neutralization reaction. J. Biol. Stand. 1984, 12, 323–337. [Google Scholar] [CrossRef]
- Available online: https://www.foot-and-mouth.org/oiefao-fmd-reference-laboratory-network/oiefao-fmd-laboratories-network-annual-reports (accessed on 23 October 2022).
- Report-Management. Available online: https://wahis.woah.org/#/ (accessed on 30 December 2022).
- Bolortsetseg, S.; Enkhtuvshin, S.; Nyamsuren, D.; Weisman, W.; Fine, A.; Yang, A.; Joly, D.O. Serosurveillance for foot-and-mouth disease in Mongolian gazelles (Procapra gutturosa) and livestock on the Eastern Steppe of Mongolia. J. Wildl. Dis. 2012, 48, 33–38. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seq-> | O/Orenburg/RUS/2021 | O/KAZ/2022 | MOG/Sukhbaatar/2022 | MOG/Khentii/2022 | MOG/Khovd/2022 |
|---|---|---|---|---|---|
| O/Orenburg/1/RUS/2021 | ID | 0.992 | 0.989 | 0.967 | 0.962 |
| O/KAZ/2022 | 0.992 | ID | 0.987 | 0.965 | 0.96 |
| MOG/Sukhbaatar/2022 | 0.989 | 0.987 | ID | 0.971 | 0.967 |
| MOG/Khentii/2022 | 0.967 | 0.965 | 0.971 | ID | 0.99 |
| MOG/Khovd/2022 | 0.962 | 0.96 | 0.967 | 0.99 | ID |
| FMDV Isolates | Postvaccination Monovalent Sera against ARRIAH Production Strains, r1 Value | ||||
|---|---|---|---|---|---|
| O1 Manisa (ME–SA) | O No. 1734 /Primorsky/2000 (O/PanAsia) | O No. 2102 /Zabaikalsky/2010 (O/Mya-98) | O No. 2212 /Primorsky/2014 (O/Mya-98) | O No. 2311/ Zabaikalsky/2016 (O/Ind-2001) | |
| O/Orenburg/2021 | 0.28 | 0.04 | 0.08 | 0.7 | 1.0 |
| O/Kazakhstan/2022 | 0.23 | 0.22 | 0.05 | 0.87 | 1.0 |
| O/Mongolia/2021 | 0.28 | 0.44 | 0.06 | 0.66 | 1.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikiforov, V.; Shcherbakov, A.; Chvala, I.; Kremenchugskaya, S.; Korennoy, F.; Mayorova, T.; Timina, A.; Tyulegenov, S.; Abdrakhmanov, S.; Berdikulov, M.; et al. Insights into the Molecular Epidemiology of Foot-and-Mouth Disease Virus in Russia, Kazakhstan, and Mongolia in Terms of O/ME-SA/Ind-2001e Sublineage Expansion. Viruses 2023, 15, 598. https://doi.org/10.3390/v15030598
Nikiforov V, Shcherbakov A, Chvala I, Kremenchugskaya S, Korennoy F, Mayorova T, Timina A, Tyulegenov S, Abdrakhmanov S, Berdikulov M, et al. Insights into the Molecular Epidemiology of Foot-and-Mouth Disease Virus in Russia, Kazakhstan, and Mongolia in Terms of O/ME-SA/Ind-2001e Sublineage Expansion. Viruses. 2023; 15(3):598. https://doi.org/10.3390/v15030598
Chicago/Turabian StyleNikiforov, Viktor, Alexey Shcherbakov, Ilya Chvala, Svetlana Kremenchugskaya, Fedor Korennoy, Tamara Mayorova, Anna Timina, Samat Tyulegenov, Sarsenbay Abdrakhmanov, Maksat Berdikulov, and et al. 2023. "Insights into the Molecular Epidemiology of Foot-and-Mouth Disease Virus in Russia, Kazakhstan, and Mongolia in Terms of O/ME-SA/Ind-2001e Sublineage Expansion" Viruses 15, no. 3: 598. https://doi.org/10.3390/v15030598