
Determinants of Chikungunya and O’nyong-Nyong Virus Specificity for Infection of Aedes and Anopheles Mosquito Vectors
 , and
, and
Abstract
:1. Introduction
1.1. Vectorial Systems of Arbovirus Transmission
1.1.1. Sympatric Distribution of Aedes and Anopheles but Differences in Pathogen Transmission
1.1.2. O’nyong-Nyong and Chikungunya, Two Closely Related Alphaviruses
1.2. Mosquito Intrinsic Factors and Virus Transmission
1.2.1. Aedes and Anopheles Genetics and Evolution
1.2.2. ONNV and CHIKV Genetics and Evolution
1.3. Vector–Virus Interactions
1.3.1. The Virus Cycle in the Mosquito Vector
Primary Midgut Infection
Midgut Escape Barrier
Viral Dissemination and Transmission
1.3.2. Mosquito Immunity and Antiviral Mechanisms
Mosquito Innate Immunity
Viral Tolerance
1.3.3. Viral Factors Underlying Host Specificity
1.4. Environmental and Human Host Factors
1.4.1. Biotic and Abiotic Factors Influencing Viral Transmission
Biotic Factors
Co-Infection with Other Pathogens
Superinfection Exclusion
Mosquito Virome
Mosquito Bacterial Microbiota
1.4.2. Abiotic Factors
Temperature and Weather
1.5. Role of Human Hosts
Host Population Genetics
2. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Rezza, G.; Chen, R.; Weaver, S.C. O’nyong-nyong fever: A neglected mosquito-borne viral disease. Pathog. Glob. Health 2017, 111, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Fabri, A.A.; Rodrigues, C.D.d.S.; Santos, C.C.d.; Chalhoub, F.L.L.; Sampaio, S.A.; Faria, N.R.d.C.; Torres, M.C.; Fonseca, V.; Brasil, P.; Calvet, G.; et al. Co-Circulation of Two Independent Clades and Persistence of CHIKV-ECSA Genotype during Epidemic Waves in Rio de Janeiro, Southeast Brazil. Pathogens 2020, 9, 984. [Google Scholar] [CrossRef]
- Amin, M.R.; Hasan, M.J.; Khan, M.A.S.; Rafi, M.A.; Islam, M.R.; Shams, T.; Islam, M.J.; Kabir, A.S.M.L.; Sharif, M.; Gozal, D. Chikungunya outbreak in Bangladesh (2017): Sociodemographic and clinical characteristics of patients from three hotspots. Trop. Med. Health 2022, 50, 9. [Google Scholar] [CrossRef] [PubMed]
- Vairo, F.; Coussoud-Mavoungou, M.P.A.; Ntoumi, F.; Castilletti, C.; Kitembo, L.; Haider, N.; Carletti, F.; Colavita, F.; Gruber, C.E.M.; Iannetta, M.; et al. Chikungunya Outbreak in the Republic of the Congo, 2019—Epidemiological, Virological and Entomological Findings of a South-North Multidisciplinary Taskforce Investigation. Viruses 2020, 12, 1020. [Google Scholar] [CrossRef] [PubMed]
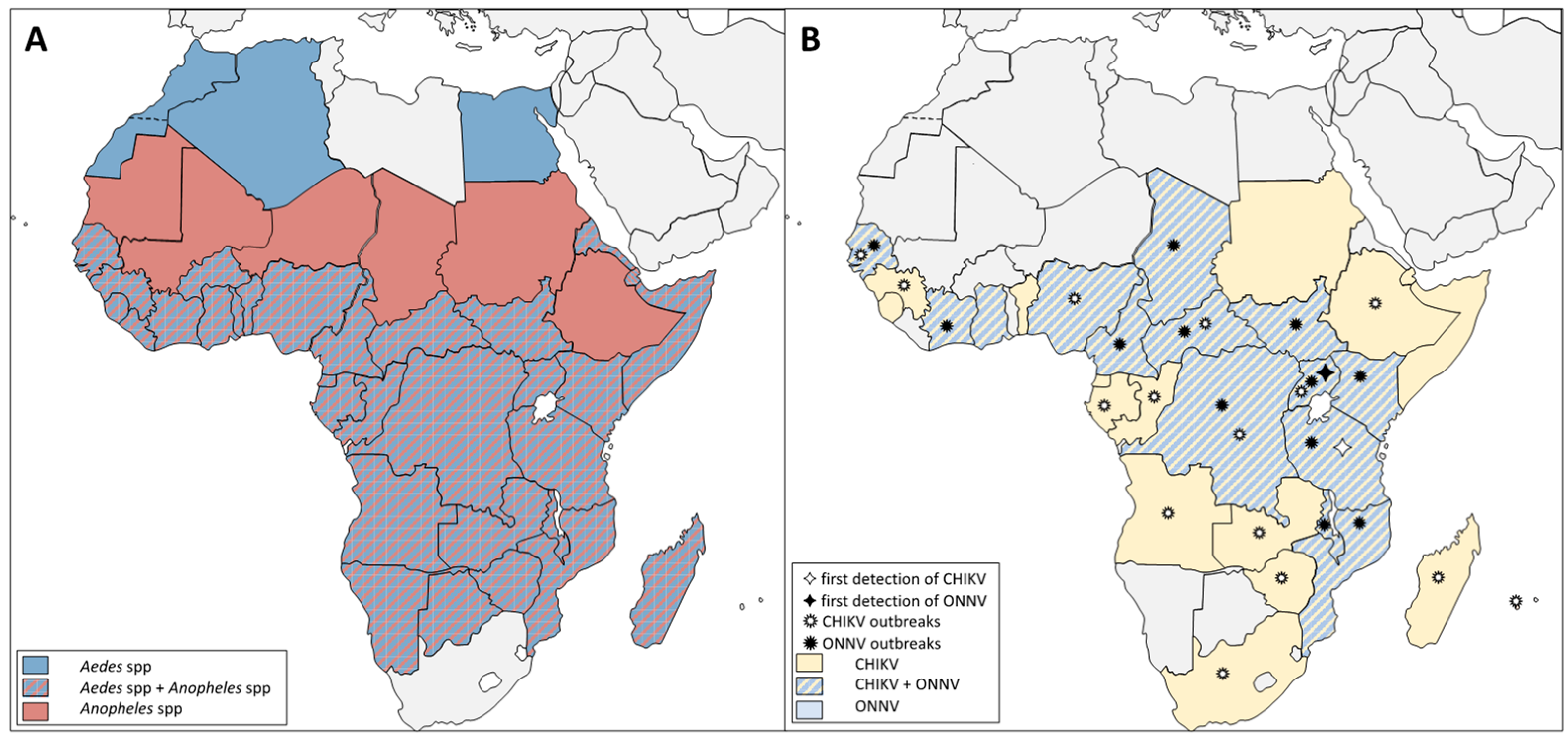
- LaBeaud, A.D.; Banda, T.; Brichard, J.; Muchiri, E.M.; Mungai, P.L.; Mutuku, F.M.; Borland, E.; Gildengorin, G.; Pfeil, S.; Teng, C.Y.; et al. High Rates of O’Nyong Nyong and Chikungunya Virus Transmission in Coastal Kenya. PLoS Negl. Trop. Dis. 2015, 9, e0003436. [Google Scholar] [CrossRef]
- Sérandour, J.; Girel, J.; Boyer, S.; Ravanel, P.; Lemperière, G.; Raveton, M. How human practices have affected vector-borne diseases in the past: A study of malaria transmission in Alpine valleys. Malar. J. 2007, 6, 115. [Google Scholar] [CrossRef] [Green Version]
- Viglietta, M.; Bellone, R.; Blisnick, A.A.; Failloux, A.-B. Vector Specificity of Arbovirus Transmission. Front. Microbiol. 2021, 12, 773211. [Google Scholar] [CrossRef] [PubMed]
- Sinka, M.E. Global Distribution of the Dominant Vector Species of Malaria. In Anopheles Mosquitoes-New Insights into Malaria Vectors; Manguin, S., Ed.; IntechOpen: Rijeka, Croatia, 2013. [Google Scholar] [CrossRef] [Green Version]
- Prudhomme, J.; Fontaine, A.; Lacour, G.; Gantier, J.-C.; Diancourt, L.; Velo, E.; Bino, S.; Reiter, P.; Mercier, A. The native European Aedes geniculatus mosquito species can transmit chikungunya virus. Emerg. Microbes Infect. 2019, 8, 962–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samarawickrema, W.A.; Sone, F.; Cummings, R.F. Natural infections of Wuchereria bancrofti in Aedes (Stegomyia) polynesiensis and Aedes (Finlaya) samoanus in Samoa. Trans. R. Soc. Trop. Med. Hyg. 1987, 81, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Boakye, D.A.; Wilson, M.D.; Appawu, M.A.; Gyapong, J. Vector competence, for Wuchereria bancrofti, of the Anopheles populations in the Bongo district of Ghana. Ann. Trop. Med. Parasitol. 2004, 98, 501–508. [Google Scholar] [CrossRef]
- Sanjuán, R.; Nebot, M.R.; Chirico, N.; Mansky, L.M.; Belshaw, R. Viral Mutation Rates. J. Virol. 2010, 84, 9733–9748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazeille, M.; Moutailler, S.; Coudrier, D.; Rousseaux, C.; Khun, H.; Huerre, M.; Thiria, J.; Dehecq, J.-S.; Fontenille, D.; Schuffenecker, I.; et al. Two Chikungunya Isolates from the Outbreak of La Reunion (Indian Ocean) Exhibit Different Patterns of Infection in the Mosquito, Aedes albopictus. PLoS ONE 2007, 2, e1168. [Google Scholar] [CrossRef]
- Strauss, J.H.; Strauss, E.G. The alphaviruses: Gene expression, replication, and evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar] [CrossRef]
- Powers, A.M.; Brault, A.C.; Shirako, Y.; Strauss, E.G.; Kang, W.; Strauss, J.H.; Weaver, S.C. Evolutionary Relationships and Systematics of the Alphaviruses. J. Virol. 2001, 75, 10118–10131. [Google Scholar] [CrossRef] [Green Version]
- Zeller, H.; Van Bortel, W.; Sudre, B. Chikungunya: Its History in Africa and Asia and Its Spread to New Regions in 2013–2014. J. Infect. Dis. 2016, 214, S436–S440. [Google Scholar] [CrossRef]
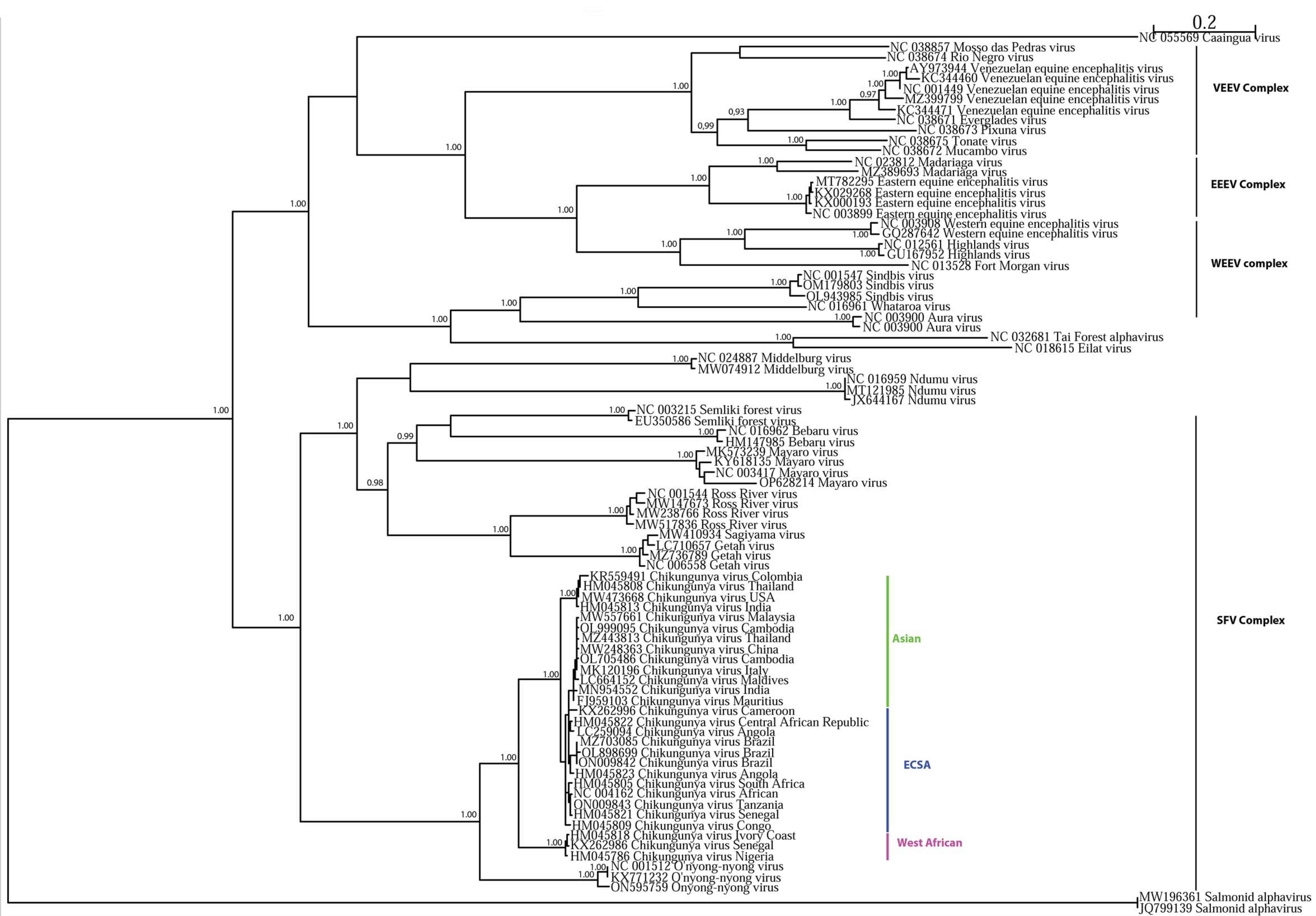
- Lo Presti, A.; Cella, E.; Angeletti, S.; Ciccozzi, M. Molecular epidemiology, evolution and phylogeny of Chikungunya virus: An updating review. Infect. Genet. Evol. 2016, 41, 270–278. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Lambert, A.J. Phylogenetic Analysis of Chikungunya Virus Strains Circulating in the Western Hemisphere. Am. J. Trop. Med. Hyg. 2016, 94, 800–803. [Google Scholar] [CrossRef] [Green Version]
- Bartholomay, L.C.; Cho, W.-L.; Rocheleau, T.A.; Boyle, J.P.; Beck, E.T.; Fuchs, J.F.; Liss, P.; Rusch, M.; Butler, K.M.; Wu, R.C.-C.; et al. Description of the Transcriptomes of Immune Response-Activated Hemocytes from the Mosquito Vectors Aedes aegypti and Armigeres subalbatus. Infect. Immun. 2004, 72, 4114–4126. [Google Scholar] [CrossRef] [Green Version]
- Pezzi, L.; LaBeaud, A.D.; Reusken, C.B.; Drexler, J.F.; Vasilakis, N.; Diallo, M.; Simon, F.; Jaenisch, T.; Gallian, P.; Sall, A.; et al. GloPID-R report on chikungunya, o’nyong-nyong and Mayaro virus, part 2: Epidemiological distribution of o’nyong-nyong virus. Antivir. Res. 2019, 172, 104611. [Google Scholar] [CrossRef]
- Russo, G.; Subissi, L.; Rezza, G. Chikungunya fever in Africa: A systematic review. Pathog. Glob. Health 2020, 114, 136–144. [Google Scholar] [CrossRef]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti Toll Pathway Controls Dengue Virus Infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Souza-Neto, J.; Cosme, R.T.; Rovira, J.; Ortiz, A.; Pascale, J.M.; Dimopoulos, G. Reciprocal Tripartite Interactions between the Aedes aegypti Midgut Microbiota, Innate Immune System and Dengue Virus Influences Vector Competence. PLoS Negl. Trop. Dis. 2012, 6, e1561. [Google Scholar] [CrossRef] [PubMed]
- Saldaña, M.A.; Etebari, K.; Hart, C.E.; Widen, S.G.; Wood, T.G.; Thangamani, S.; Asgari, S.; Hughes, G.L. Zika virus alters the microRNA expression profile and elicits an RNAi response in Aedes aegypti mosquitoes. PLoS Negl. Trop. Dis. 2017, 11, e0005760. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Srivastava, P.; Sirisena, P.; Dubey, S.K.; Kumar, R.; Shrinet, J.; Sunil, S. Mosquito Innate Immunity. Insects 2018, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Adusei, J.A.; Narkwa, P.W.; Owusu, M.; Domfeh, S.A.; Alhassan, M.; Appau, E.; Salam, A.; Mutocheluh, M. Evidence of chikungunya virus infections among febrile patients at three secondary health facilities in the Ashanti and the Bono Regions of Ghana. PLoS Negl. Trop. Dis. 2021, 15, e0009735. [Google Scholar] [CrossRef] [PubMed]
- Nene, V.; Wortman, J.R.; Lawson, D.; Haas, B.; Kodira, C.; Tu, Z.J.; Loftus, B.; Xi, Z.; Megy, K.; Grabherr, M.; et al. Genome Sequence of Aedes aegypti, a Major Arbovirus Vector. Science 2007, 316, 1718–1723. [Google Scholar] [CrossRef] [Green Version]
- Ngoagouni, C.; Kamgang, B.; Kazanji, M.; Paupy, C.; Nakouné, E. Potential of Aedes aegypti and Aedes albopictus populations in the Central African Republic to transmit enzootic chikungunya virus strains. Parasites Vectors 2017, 10, 164. [Google Scholar] [CrossRef] [Green Version]
- Gudo, E.S.; Black, J.F.P.; Cliff, J.L. Chikungunya in Mozambique: A Forgotten History. PLoS Negl. Trop. Dis. 2016, 10, e0005001. [Google Scholar] [CrossRef] [PubMed]
- Fourié, T.; Dia, A.; Savreux, Q.; de Santi, V.P.; de Lamballerie, X.; Leparc-Goffart, I.; Simon, F. Emergence of Indian lineage of ECSA chikungunya virus in Djibouti, 2019. Int. J. Infect. Dis. 2021, 108, 198–201. [Google Scholar] [CrossRef] [PubMed]
- da Silva, A.F.; Machado, L.C.; de Paula, M.B.; Vieira, C.J.D.S.P.; Bronzoni, R.V.D.M.; Santos, M.A.V.D.M.; Wallau, G.L. Culicidae evolutionary history focusing on the Culicinae subfamily based on mitochondrial phylogenomics. Sci. Rep. 2020, 10, 18823. [Google Scholar] [CrossRef] [PubMed]
- Logue, K.; Chan, E.R.; Phipps, T.; Small, S.T.; Reimer, L.; Henry-Halldin, C.; Sattabongkot, J.; Siba, P.M.; Zimmerman, P.A.; Serre, D. Mitochondrial genome sequences reveal deep divergences among Anopheles punctulatus sibling species in Papua New Guinea. Malar. J. 2013, 12, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neafsey, D.E.; Waterhouse, R.M.; Abai, M.R.; Aganezov, S.S.; Alekseyev, M.A.; Allen, J.E.; Amon, J.; Arcà, B.; Arensburger, P.; Artemov, G.; et al. Highly evolvable malaria vectors: The genomes of 16 Anopheles mosquitoes. Science 2015, 347, 1258522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, J.E.; Alves, J.M.; Palmer, W.J.; Day, J.P.; Sylla, M.; Ramasamy, R.; Surendran, S.N.; Black, W.C.T.; Pain, A.; Jiggins, F.M. Population genomics reveals that an anthropophilic population of Aedes aegypti mosquitoes in West Africa recently gave rise to American and Asian populations of this major disease vector. BMC Biol. 2017, 15, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.E.; Evans, B.R.; Zheng, W.; Obas, V.; Barrera-Martinez, L.; Egizi, A.; Zhao, H.; Caccone, A.; Powell, J.R. Human Impacts Have Shaped Historical and Recent Evolution in Aedes aegypti, the Dengue and Yellow Fever Mosquito. Evolution 2014, 68, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Holt, R.A.; Subramanian, G.M.; Halpern, A.; Sutton, G.G.; Charlab, R.; Nusskern, D.R.; Wincker, P.; Clark, A.G.; Ribeiro, J.M.C.; Wides, R.; et al. The Genome Sequence of the Malaria Mosquito Anopheles gambiae. Science 2002, 298, 129–149. [Google Scholar] [CrossRef]
- Severson, D.W.; Behura, S.K. Mosquito Genomics: Progress and Challenges. Annu. Rev. Èntomol. 2012, 57, 143–166. [Google Scholar] [CrossRef]
- Timoshevskiy, V.A.; Kinney, N.A.; Debruyn, B.S.; Mao, C.; Tu, Z.; Severson, D.W.; Sharakhov, I.V.; Sharakhova, M.V. Genomic composition and evolution of Aedes aegypti chromosomes revealed by the analysis of physically mapped supercontigs. BMC Biol. 2014, 12, 27. [Google Scholar] [CrossRef] [Green Version]
- Amarante, A.D.M.; Jupatanakul, N.; Silva, I.C.D.A.D.; Carneiro, V.C.; Vicentino, A.R.R.; Dimopolous, G.; Talyuli, O.A.C.; Fantappié, M.R. The DNA chaperone HMGB1 potentiates the transcriptional activity of Rel1A in the mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 2017, 80, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Joosten, J.; Miesen, P.; Taşköprü, E.; Pennings, B.; Jansen, P.W.T.C.; Huynen, M.A.; Vermeulen, M.; Van Rij, R.P. The Tudor protein Veneno assembles the ping-pong amplification complex that produces viral piRNAs in Aedes mosquitoes. Nucleic Acids Res. 2019, 47, 2546–2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keene, K.M.; Foy, B.D.; Sanchez-Vargas, I.; Beaty, B.J.; Blair, C.D.; Olson, K.E. RNA interference acts as a natural antiviral response to O’nyong-nyong virus (Alphavirus; Togaviridae) infection of Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2004, 101, 17240–17245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morazzani, E.M.; Wiley, M.R.; Murreddu, M.G.; Adelman, Z.N.; Myles, K.M. Production of Virus-Derived Ping-Pong-Dependent piRNA-like Small RNAs in the Mosquito Soma. PLoS Pathog. 2012, 8, e1002470. [Google Scholar] [CrossRef]
- Weaver, S.C.; Forrester, N.L. Chikungunya: Evolutionary history and recent epidemic spread. Antivir. Res. 2015, 120, 32–39. [Google Scholar] [CrossRef]
- Johnson, B.K. O’nyong-nyong Virus Disease. In The Arboviruses: Epidemiology and Ecology; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Bessaud, M.; Peyrefitte, C.N.; Pastorino, B.A.; Gravier, P.; Tock, F.; Boete, F.; Tolou, H.J.; Grandadam, M. O’nyong-nyong Virus, Chad. Emerg. Infect. Dis. 2006, 12, 1248–1250. [Google Scholar] [CrossRef]
- Vazeille, M.; Zouache, K.; Vega-Rúa, A.; Thiberge, J.-M.; Caro, V.; Yébakima, A.; Mousson, L.; Piorkowski, G.; Dauga, C.; Vaney, M.-C.; et al. Importance of mosquito “quasispecies” in selecting an epidemic arthropod-borne virus. Sci. Rep. 2016, 6, 29564. [Google Scholar] [CrossRef] [Green Version]
- Mandary, M.B.; Masomian, M.; Poh, C.L. Impact of RNA Virus Evolution on Quasispecies Formation and Virulence. Int. J. Mol. Sci. 2019, 20, 4657. [Google Scholar] [CrossRef] [Green Version]
- Weaver, S.C.; Forrester, N.L.; Liu, J.; Vasilakis, N. Population bottlenecks and founder effects: Implications for mosquito-borne arboviral emergence. Nat. Rev. Microbiol. 2021, 19, 184–195. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Ludwig, M.L.; Rwaguma, E.B.; Lutwama, J.J.; Kram, T.M.; Karabatsos, N.; Cropp, B.C.; Miller, B.R. Emergence of Epidemic O’nyong-nyong Fever in Uganda after a 35-Year Absence: Genetic Characterization of the Virus. Virology 1998, 252, 258–268. [Google Scholar] [CrossRef] [Green Version]
- Myles, K.M.; Kelly, C.L.H.; Ledermann, J.P.; Powers, A.M. Effects of an Opal Termination Codon Preceding the nsP4 Gene Sequence in the O’Nyong-Nyong Virus Genome on Anopheles gambiae Infectivity. J. Virol. 2006, 80, 4992–4997. [Google Scholar] [CrossRef] [Green Version]
- Götte, B.; Liu, L.; McInerney, G.M. The Enigmatic Alphavirus Non-Structural Protein 3 (nsP3) Revealing Its Secrets at Last. Viruses 2018, 10, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.E.; Long, K.M.; Whitmore, A.C.; Sanders, W.; Thurlow, L.R.; Brown, J.A.; Morrison, C.R.; Vincent, H.; Peck, K.M.; Browning, C.; et al. Disruption of the Opal Stop Codon Attenuates Chikungunya Virus-Induced Arthritis and Pathology. Mbio 2017, 8, e01456-17. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.C.; Kam, Y.-W.; Lin, R.P.T.; Ng, M.M.-L.; Ng, L.F.; Chu, J.H.J. Comparative analysis of the genome sequences and replication profiles of chikungunya virus isolates within the East, Central and South African (ECSA) lineage. Virol. J. 2013, 10, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, A.W.E.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue Barriers to Arbovirus Infection in Mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourya, D.T.; Ranadive, S.N.; Gokhale, M.D.; Barde, P.V.; Padbidri, V.S.; Banerjee, K. Putative chikungunya virus-specific receptor proteins on the midgut brush border membrane of Aedes aegypti mosquito. Indian J. Med. Res. 1998, 107, 10–14. [Google Scholar] [PubMed]
- Fongsaran, C.; Jirakanwisal, K.; Kuadkitkan, A.; Wikan, N.; Wintachai, P.; Thepparit, C.; Ubol, S.; Phaonakrop, N.; Roytrakul, S.; Smith, D.R. Involvement of ATP synthase β subunit in chikungunya virus entry into insect cells. Arch. Virol. 2014, 159, 3353–3364. [Google Scholar] [CrossRef]
- Paingankar, M.S.; Gokhale, M.D.; Deobagkar, D.N. Dengue-2-virus-interacting polypeptides involved in mosquito cell infection. Arch. Virol. 2010, 155, 1453–1461. [Google Scholar] [CrossRef]
- Rider, M.A.; Zou, J.; VanLandingham, D.; Nuckols, J.T.; Higgs, S.; Zhang, Q.; Lacey, M.; Kim, J.; Wang, G.; Hong, Y.S. Quantitative Proteomic Analysis of the Anopheles gambiae (Diptera: Culicidae) Midgut Infected with O’nyong–Nyong Virus. J. Med. Èntomol. 2013, 50, 1077–1088. [Google Scholar] [CrossRef] [Green Version]
- Colpitts, T.M.; Cox, J.; Nguyen, A.; Feitosa, F.; Krishnan, M.N.; Fikrig, E. Use of a tandem affinity purification assay to detect interactions between West Nile and dengue viral proteins and proteins of the mosquito vector. Virology 2011, 417, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Bernard, E.; Solignat, M.; Gay, B.; Chazal, N.; Higgs, S.; Devaux, C.; Briant, L. Endocytosis of Chikungunya Virus into Mammalian Cells: Role of Clathrin and Early Endosomal Compartments. PLoS ONE 2010, 5, e11479. [Google Scholar] [CrossRef] [Green Version]
- Kantor, A.M.; Grant, D.G.; Balaraman, V.; White, T.A.; Franz, A.W.E. Ultrastructural Analysis of Chikungunya Virus Dissemination from the Midgut of the Yellow Fever Mosquito, Aedes aegypti. Viruses 2018, 10, 571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rückert, C.; Ebel, G.D. How Do Virus–Mosquito Interactions Lead to Viral Emergence? Trends Parasitol. 2018, 34, 310–321. [Google Scholar] [CrossRef]
- Dong, S.; Balaraman, V.; Kantor, A.M.; Lin, J.; Grant, D.G.; Held, N.L.; Franz, A.W.E. Chikungunya virus dissemination from the midgut of Aedes aegypti is associated with temporal basal lamina degradation during bloodmeal digestion. PLoS Negl. Trop. Dis. 2017, 11, e0005976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, S.; Kantor, A.M.; Lin, J.; Passarelli, A.L.; Clem, R.J.; Franz, A.W.E. Infection pattern and transmission potential of chikungunya virus in two New World laboratory-adapted Aedes aegypti strains. Sci. Rep. 2016, 6, 24729. [Google Scholar] [CrossRef] [Green Version]
- Carissimo, G.; Pondeville, E.; McFarlane, M.; Dietrich, I.; Mitri, C.; Bischoff, E.; Antoniewski, C.; Bourgouin, C.; Failloux, A.-B.; Kohl, A.; et al. Antiviral immunity of Anopheles gambiae is highly compartmentalized, with distinct roles for RNA interference and gut microbiota. Proc. Natl. Acad. Sci. USA 2015, 112, E176–E185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro, V.V.S.; Navegantes-Lima, K.C.; De Lemos, A.B.; da Silva, G.L.; de Souza Gomes, R.; Reis, J.F.; Junior, L.C.R.; Da Silva, O.S.; Romão, P.R.T.; Monteiro, M.C. Aedes–Chikungunya Virus Interaction: Key Role of Vector Midguts Microbiota and Its Saliva in the Host Infection. Front. Microbiol. 2019, 10, 492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogels, C.B.; Goöertz, G.P.; Pijlman, G.P.; Koenraadt, C.J. Vector competence of European mosquitoes for West Nile virus. Emerg. Microbes Infect. 2017, 6, e96. [Google Scholar] [CrossRef] [Green Version]
- Tchankouo-Nguetcheu, S.; Khun, H.; Pincet, L.; Roux, P.; Bahut, M.; Huerre, M.; Guette, C.; Choumet, V. Differential Protein Modulation in Midguts of Aedes aegypti Infected with Chikungunya and Dengue 2 Viruses. PLoS ONE 2010, 5, e13149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanLandingham, D.L.; Lehane, M.J.; Hong, C.; Powers, A.M.; McElroy, K.L.; Tsetsarkin, K.; Klingler, K.; Higgs, S. Differential Infectivities of O’nyong-Nyong and Chikungunya Virus Isolates in Anopheles gambiae and Aedes aegypti Mosquitoes. Am. J. Trop. Med. Hyg. 2005, 72, 616–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFarlane, M.; Arias-Goeta, C.; Martin, E.; O’Hara, Z.; Lulla, A.; Mousson, L.; Rainey, S.M.; Misbah, S.; Schnettler, E.; Donald, C.L.; et al. Characterization of Aedes aegypti Innate-Immune Pathways that Limit Chikungunya Virus Replication. PLoS Negl. Trop. Dis. 2014, 8, e2994. [Google Scholar] [CrossRef] [PubMed]
- Angleró-Rodríguez, Y.I.; MacLeod, H.J.; Kang, S.; Carlson, J.S.; Jupatanakul, N.; Dimopoulos, G. Aedes aegypti Molecular Responses to Zika Virus: Modulation of Infection by the Toll and Jak/Stat Immune Pathways and Virus Host Factors. Front. Microbiol. 2017, 8, 2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldock, J.; Olson, K.E.; Christophides, G.K. Anopheles gambiae Antiviral Immune Response to Systemic O’nyong-nyong Infection. PLoS Negl. Trop. Dis. 2012, 6, e1565. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.; Jan, E.; Sarnow, P.; Schneider, D. The Imd Pathway Is Involved in Antiviral Immune Responses in Drosophila. PLoS ONE 2009, 4, e7436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Vargas, I.; Scott, J.C.; Poole-Smith, B.K.; Franz, A.W.E.; Barbosa-Solomieu, V.; Wilusz, J.; Olson, K.E.; Blair, C.D. Dengue Virus Type 2 Infections of Aedes aegypti Are Modulated by the Mosquito’s RNA Interference Pathway. PLOS Pathog. 2009, 5, e1000299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, G.; Liu, L.; Wang, P.; Zhang, Y.; Zhao, Y.O.; Colpitts, T.M.; Feitosa, F.; Anderson, J.F.; Fikrig, E. An In Vivo Transfection Approach Elucidates a Role for Aedes aegypti Thioester-Containing Proteins in Flaviviral Infection. PLoS ONE 2011, 6, e22786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagueux, M.; Perrodou, E.; Levashina, E.A.; Capovilla, M.; Hoffmann, J.A. Constitutive expression of a complement-like protein in Toll and JAK gain-of-function mutants of Drosophila. Proc. Natl. Acad. Sci. USA 2000, 97, 11427–11432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraiture, M.; Baxter, R.H.G.; Steinert, S.; Chelliah, Y.; Frolet, C.; Quispe-Tintaya, W.; Hoffmann, J.A.; Blandin, S.A.; Levashina, E.A. Two Mosquito LRR Proteins Function as Complement Control Factors in the TEP1-Mediated Killing of Plasmodium. Cell Host Microbe 2009, 5, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Povelones, M.; Waterhouse, R.M.; Kafatos, F.C.; Christophides, G.K. Leucine-Rich Repeat Protein Complex Activates Mosquito Complement in Defense Against Plasmodium Parasites. Science 2009, 324, 258–261. [Google Scholar] [CrossRef] [Green Version]
- Mitri, C.; Bischoff, E.; Takashima, E.; Williams, M.; Eiglmeier, K.; Pain, A.; Guelbeogo, W.M.; Gnémé, A.; Brito-Fravallo, E.; Holm, I.; et al. An Evolution-Based Screen for Genetic Differentiation between Anopheles Sister Taxa Enriches for Detection of Functional Immune Factors. PLoS Pathog. 2015, 11, e1005306. [Google Scholar] [CrossRef]
- Waterhouse, R.M.; Povelones, M.; Christophides, G.K. Sequence-structure-function relations of the mosquito leucine-rich repeat immune proteins. BMC Genom. 2010, 11, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Alto, B.W.; Shin, D. Transcriptional Profile of Aedes aegypti Leucine-Rich Repeat Proteins in Response to Zika and Chikungunya Viruses. Int. J. Mol. Sci. 2019, 20, 615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medzhitov, R.; Schneider, D.S.; Soares, M.P. Disease Tolerance as a Defense Strategy. Science 2012, 335, 936–941. [Google Scholar] [CrossRef] [Green Version]
- Goic, B.; Stapleford, K.A.; Frangeul, L.; Doucet, A.J.; Gausson, V.; Blanc, H.; Schemmel-Jofre, N.; Cristofari, G.; Lambrechts, L.; Vignuzzi, M.; et al. Virus-derived DNA drives mosquito vector tolerance to arboviral infection. Nat. Commun. 2016, 7, 12410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goic, B.; Vodovar, N.; Mondotte, J.A.; Monot, C.; Frangeul, L.; Blanc, H.; Gausson, V.; Vera-Otarola, J.; Cristofari, G.; Saleh, M.-C. RNA-mediated interference and reverse transcription control the persistence of RNA viruses in the insect model Drosophila. Nat. Immunol. 2013, 14, 396–403. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Swevers, L.; Kolliopoulou, A.; Smagghe, G. Arboviruses and the Challenge to Establish Systemic and Persistent Infections in Competent Mosquito Vectors: The Interaction with the RNAi Mechanism. Front. Physiol. 2019, 10, 890. [Google Scholar] [CrossRef]
- Lewis, S.H.; Salmela, H.; Obbard, D.J. Duplication and Diversification of Dipteran Argonaute Genes, and the Evolutionary Divergence of Piwi and Aubergine. Genome Biol. Evol. 2016, 8, 507–518. [Google Scholar] [CrossRef]
- Olson, K.E.; Bonizzoni, M. Nonretroviral integrated RNA viruses in arthropod vectors: An occasional event or something more? Curr. Opin. Insect Sci. 2017, 22, 45–53. [Google Scholar] [CrossRef]
- Palatini, U.; Miesen, P.; Carballar-Lejarazu, R.; Ometto, L.; Rizzo, E.; Tu, Z.; van Rij, R.P.; Bonizzoni, M. Comparative genomics shows that viral integrations are abundant and express piRNAs in the arboviral vectors Aedes aegypti and Aedes albopictus. BMC Genom. 2017, 18, 512. [Google Scholar] [CrossRef] [Green Version]
- Fujino, K.; Horie, M.; Honda, T.; Merriman, D.K.; Tomonaga, K. Inhibition of Borna disease virus replication by an endogenous bornavirus-like element in the ground squirrel genome. Proc. Natl. Acad. Sci. USA 2014, 111, 13175–13180. [Google Scholar] [CrossRef] [Green Version]
- Buckley, S.M., II. Multiplication of Chikungunya and O’nyong-nyong Viruses in Singh’s Aedes Cell Lines. Curr. Top. Microbiol. Immunol. 1971, 55, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; McGee, C.E.; Volk, S.M.; Van Landingham, D.L.; Weaver, S.C.; Higgs, S. Epistatic Roles of E2 Glycoprotein Mutations in Adaption of Chikungunya Virus to Aedes albopictus and Ae. Aegypti Mosquitoes. PLoS ONE 2009, 4, e6835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matusali, G.; Colavita, F.; Bordi, L.; Lalle, E.; Ippolito, G.; Capobianchi, M.R.; Castilletti, C. Tropism of the Chikungunya Virus. Viruses 2019, 11, 175. [Google Scholar] [CrossRef] [Green Version]
- Brault, A.C.; Foy, B.D.; Myles, K.M.; Kelly, C.L.H.; Higgs, S.; Weaver, S.C.; Olson, K.E.; Miller, B.R.; Powers, A.M. Infection patterns of o’nyong nyong virus in the malaria-transmitting mosquito, Anopheles gambiae. Insect Mol. Biol. 2004, 13, 625–635. [Google Scholar] [CrossRef]
- VanLandingham, D.L.; Higgs, S.; Hong, C.; Klingler, K.A.; McElroy, K.L.; Lehane, M.J.; Tsetsarkin, K. Determinants of Vector Specificity of O’nyong Nyong and Chikungunya Viruses in Anopheles and Aedes Mosquitoes. Am. J. Trop. Med. Hyg. 2006, 74, 663–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lello, L.S.; Utt, A.; Bartholomeeusen, K.; Wang, S.; Rausalu, K.; Kendall, C.; Coppens, S.; Fragkoudis, R.; Tuplin, A.; Alphey, L.; et al. Cross-utilisation of template RNAs by alphavirus replicases. PLoS Pathog. 2020, 16, e1008825. [Google Scholar] [CrossRef]
- Saxton-Shaw, K.D.; Ledermann, J.P.; Borland, E.M.; Stovall, J.L.; Mossel, E.C.; Singh, A.J.; Wilusz, J.; Powers, A.M. O’nyong nyong Virus Molecular Determinants of Unique Vector Specificity Reside in Non-Structural Protein. PLoS Negl. Trop. Dis. 2013, 7, e1931. [Google Scholar] [CrossRef] [Green Version]
- Malet, H.; Coutard, B.; Jamal, S.; Dutartre, H.; Papageorgiou, N.; Neuvonen, M.; Ahola, T.; Forrester, N.; Gould, E.A.; Lafitte, D.; et al. The Crystal Structures of Chikungunya and Venezuelan Equine Encephalitis Virus nsP3 Macro Domains Define a Conserved Adenosine Binding Pocket. J. Virol. 2009, 83, 6534–6545. [Google Scholar] [CrossRef] [Green Version]
- Shin, G.; Yost, S.A.; Miller, M.T.; Elrod, E.J.; Grakoui, A.; Marcotrigiano, J. Structural and functional insights into alphavirus polyprotein processing and pathogenesis. Proc. Natl. Acad. Sci. USA 2012, 109, 16534–16539. [Google Scholar] [CrossRef] [Green Version]
- Vihinen, H.; Saarinen, J. Phosphorylation Site Analysis of Semliki Forest Virus Nonstructural Protein J. Biol. Chem. 2000, 275, 27775–27783. [Google Scholar] [CrossRef] [Green Version]
- Agback, P.; Shernyukov, A.; Dominguez, F.; Agback, T.; Frolova, E.I. Novel NMR Assignment Strategy Reveals Structural Heterogeneity in Solution of the nsP3 HVD Domain of Venezuelan Equine Encephalitis Virus. Molecules 2020, 25, 5824. [Google Scholar] [CrossRef]
- Göertz, G.P.; Lingemann, M.; Geertsema, C.; Abma-Henkens, M.H.C.; Vogels, C.B.F.; Koenraadt, C.J.M.; van Oers, M.M.; Pijlman, G.P. Conserved motifs in the hypervariable domain of chikungunya virus nsP3 required for transmission by Aedes aegypti mosquitoes. PLoS Negl. Trop. Dis. 2018, 12, e0006958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kedersha, N.; Panas, M.D.; Achorn, C.A.; Lyons, S.; Tisdale, S.; Hickman, T.; Thomas, M.; Lieberman, J.; McInerney, G.M.; Ivanov, P.; et al. G3BP–Caprin1–USP10 complexes mediate stress granule condensation and associate with 40S subunits. J. Cell Biol. 2016, 212, 845–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholte, F.E.M.; Tas, A.; Albulescu, I.C.; Žusinaite, E.; Merits, A.; Snijder, E.J.; van Hemert, M.J. Stress Granule Components G3BP1 and G3BP2 Play a Proviral Role Early in Chikungunya Virus Replication. J. Virol. 2015, 89, 4457–4469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fros, J.J.; Geertsema, C.; Zouache, K.; Baggen, J.; Domeradzka, N.; van Leeuwen, D.M.; Flipse, J.; Vlak, J.M.; Failloux, A.-B.; Pijlman, G.P. Mosquito Rasputin interacts with chikungunya virus nsP3 and determines the infection rate in Aedes albopictus. Parasites Vectors 2015, 8, 464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowee, G.; Bakker, J.W.; Geertsema, C.; Ros, V.I.D.; Göertz, G.P.; Fros, J.J.; Pijlman, G.P. A Tale of 20 Alphaviruses; Inter-species Diversity and Conserved Interactions between Viral Non-structural Protein 3 and Stress Granule Proteins. Front. Cell Dev. Biol. 2021, 9, 625711. [Google Scholar] [CrossRef]
- Abdullah, N.; Ahemad, N.; Aliazis, K.; Khairat, J.E.; Lee, T.C.; Ahmad, S.A.A.; Adnan, N.A.A.; Macha, N.O.; Hassan, S.S. The Putative Roles and Functions of Indel, Repetition and Duplication Events in Alphavirus Non-Structural Protein 3 Hypervariable Domain (nsP3 HVD) in Evolution, Viability and Re-Emergence. Viruses 2021, 13, 1021. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Srivastava, P.; Mathur, K.; Shrinet, J.; Dubey, S.K.; Chinnappan, M.; Kaur, I.; Nayak, D.; Chattopadhyay, S.; Bhatnagar, R.K.; et al. Chikungunya virus non-structural protein nsP3 interacts with Aedes aegypti DEAD-box helicase RM62F. VirusDisease 2021, 32, 657–665. [Google Scholar] [CrossRef]
- Saron, W.; Rathore, A.; Lim, T.; Ooi, E.E.; Low, J.; Abraham, S.; St. John, A.L. Flavivirus serocomplex cross-reactive immunity is protective by activating heterologous memory CD4 T cells. Int. J. Infect. Dis. 2019, 79, 12–13. [Google Scholar] [CrossRef] [Green Version]
- Wichit, S.; Gumpangseth, N.; Hamel, R.; Yainoy, S.; Arikit, S.; Punsawad, C.; Missé, D. Chikungunya and Zika Viruses: Co-Circulation and the Interplay between Viral Proteins and Host Factors. Pathogens 2021, 10, 448. [Google Scholar] [CrossRef]
- Rückert, C.; Weger-Lucarelli, J.; Garcia-Luna, S.M.; Young, M.C.; Byas, A.D.; Murrieta, R.A.; Fauver, J.R.; Ebel, G.D. Impact of simultaneous exposure to arboviruses on infection and transmission by Aedes aegypti mosquitoes. Nat. Commun. 2017, 8, 15412. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.B.; Vrati, S.; Kalia, M. Pathobiology of Japanese encephalitis virus infection. Mol. Asp. Med. 2021, 81, 100994. [Google Scholar] [CrossRef] [PubMed]
- Roth, A.; Mercier, A.; Lepers, C.; Hoy, D.; Duituturaga, S.; Benyon, E.; Guillaumot, L.; Souarès, Y. Concurrent outbreaks of dengue, chikungunya and Zika virus infections–an unprecedented epidemic wave of mosquito-borne viruses in the Pacific 2012–2014. Eurosurveillance 2014, 19, 20929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothan, H.A.; Bidokhti, M.R.M.; Byrareddy, S.N. Current concerns and perspectives on Zika virus co-infection with arboviruses and HIV. J. Autoimmun. 2018, 89, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Phommanivong, V.; Kanda, S.; Shimono, T.; Lamaningao, P.; Darcy, A.W.; Mishima, N.; Phaytanavanh, B.; Nishiyama, T. Co-circulation of the dengue with chikungunya virus during the 2013 outbreak in the southern part of Lao PDR. Trop. Med. Health 2016, 44, 24. [Google Scholar] [CrossRef] [Green Version]
- Caron, M.; Paupy, C.; Grard, G.; Becquart, P.; Mombo, I.; Nso, B.B.B.; Kassa, F.K.; Nkoghe, D.; Leroy, E.M. Recent Introduction and Rapid Dissemination of Chikungunya Virus and Dengue Virus Serotype 2 Associated with Human and Mosquito Coinfections in Gabon, Central Africa. Clin. Infect. Dis. 2012, 55, e45–e53. [Google Scholar] [CrossRef] [Green Version]
- Göertz, G.P.; Vogels, C.B.F.; Geertsema, C.; Koenraadt, C.J.M.; Pijlman, G.P. Mosquito co-infection with Zika and chikungunya virus allows simultaneous transmission without affecting vector competence of Aedes aegypti. PLoS Negl. Trop. Dis. 2017, 11, e0005654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieme, C.; Zmarlak, N.M.; Brito-Fravallo, E.; Travaillé, C.; Pain, A.; Cherrier, F.; Genève, C.; Alvarez, E.C.; Riehle, M.M.; Vernick, K.D.; et al. Exposure of Anopheles mosquitoes to trypanosomes reduces reproductive fitness and enhances susceptibility to Plasmodium. PLoS Negl. Trop. Dis. 2020, 14, e0008059. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.M.; Ndenga, B.A.; Mutuku, F.M.; Okuta, V.; Ronga, C.O.; Chebii, P.K.; Maina, P.; Jembe, Z.; Sahoo, M.K.; Huang, C.; et al. No Evidence of O’nyong-Nyong Viremia among Children with Febrile Illness in Kenya (2015–2018). Am. J. Trop. Med. Hyg. 2021, 104, 1435–1437. [Google Scholar] [CrossRef]
- Torres-Ruesta, A.; Teo, T.-H.; Chan, Y.-H.; Amrun, S.N.; Yeo, N.K.-W.; Lee, C.Y.-P.; Nguee, S.Y.-T.; Tay, M.Z.; Nosten, F.; Fong, S.-W.; et al. Malaria abrogates O’nyong–nyong virus pathologies by restricting virus infection in nonimmune cells. Life Sci. Alliance 2022, 5, e202101272. [Google Scholar] [CrossRef]
- Boussier, J.; Levi, L.; Weger-Lucarelli, J.; Poirier, E.Z.; Vignuzzi, M.; Albert, M.L. Chikungunya virus superinfection exclusion is mediated by a block in viral replication and does not rely on non-structural protein 2. PLoS ONE 2020, 15, e0241592. [Google Scholar] [CrossRef] [PubMed]
- Kantor, A.M.; Lin, J.; Wang, A.; Thompson, D.C.; Franz, A.W.E. Infection Pattern of Mayaro Virus in Aedes aegypti (Diptera: Culicidae) and Transmission Potential of the Virus in Mixed Infections with Chikungunya Virus. J. Med. Èntomol. 2019, 56, 832–843. [Google Scholar] [CrossRef] [PubMed]
- Minkeu, F.N.; Vernick, K.D. A Systematic Review of the Natural Virome of Anopheles Mosquitoes. Viruses 2018, 10, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakrzewski, M.; Rašić, G.; Darbro, J.; Krause, L.; Poo, Y.S.; Filipović, I.; Parry, R.; Asgari, S.; Devine, G.; Suhrbier, A. Mapping the virome in wild-caught Aedes aegypti from Cairns and Bangkok. Sci. Rep. 2018, 8, 4690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguiar, E.R.; Olmo, R.P.; Paro, S.; Ferreira, F.V.; Faria, I.J.D.S.D.; Todjro, Y.M.H.; Lobo, F.P.; Kroon, E.G.; Meignin, C.; Gatherer, D.; et al. Sequence-independent characterization of viruses based on the pattern of viral small RNAs produced by the host. Nucleic Acids Res. 2015, 43, 6191–6206. [Google Scholar] [CrossRef]
- Thannesberger, J.; Rascovan, N.; Eisenmann, A.; Klymiuk, I.; Zittra, C.; Fuehrer, H.-P.; Scantlebury-Manning, T.; Gittens-St. Hilaire, M.; Austin, S.; Landis, R.C.; et al. Highly Sensitive Virome Characterization of Aedes aegypti and Culex pipiens Complex from Central Europe and the Caribbean Reveals Potential for Interspecies Viral Transmission. Pathogens 2020, 9, 686. [Google Scholar] [CrossRef]
- Fauver, J.R.; Grubaugh, N.D.; Krajacich, B.J.; Weger-Lucarelli, J.; Lakin, S.M.; Fakoli, L.S., III; Bolay, F.K.; Diclaro, J.W., II; Dabiré, K.R.; Foy, B.D.; et al. West African Anopheles gambiae mosquitoes harbor a taxonomically diverse virome including new insect-specific flaviviruses, mononegaviruses, and totiviruses. Virology 2016, 498, 288–299. [Google Scholar] [CrossRef] [Green Version]
- Belda, E.; Nanfack-Minkeu, F.; Eiglmeier, K.; Carissimo, G.; Holm, I.; Diallo, M.; Diallo, D.; Vantaux, A.; Kim, S.; Sharakhov, I.V.; et al. De novo profiling of RNA viruses in Anopheles malaria vector mosquitoes from forest ecological zones in Senegal and Cambodia. BMC Genom. 2019, 20, 664. [Google Scholar] [CrossRef] [Green Version]
- de Almeida, J.P.; Aguiar, E.R.; Armache, J.N.; Olmo, R.P.; Marques, J.T. The virome of vector mosquitoes. Curr. Opin. Virol. 2021, 49, 7–12. [Google Scholar] [CrossRef]
- Carissimo, G.; Eiglmeier, K.; Reveillaud, J.; Holm, I.; Diallo, M.; Diallo, D.; Vantaux, A.; Kim, S.; Ménard, D.; Siv, S.; et al. Identification and Characterization of Two Novel RNA Viruses from Anopheles gambiae Species Complex Mosquitoes. PLoS ONE 2016, 11, e0153881. [Google Scholar] [CrossRef] [Green Version]
- Bolling, B.G.; Olea-Popelka, F.J.; Eisen, L.; Moore, C.G.; Blair, C.D. Transmission dynamics of an insect-specific flavivirus in a naturally infected Culex pipiens laboratory colony and effects of co-infection on vector competence for West Nile virus. Virology 2012, 427, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Nasar, F.; Erasmus, J.H.; Haddow, A.D.; Tesh, R.B.; Weaver, S.C. Eilat virus induces both homologous and heterologous interference. Virology 2015, 484, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, M.J.; Frydman, H.M.; Connor, J.H. Dual Insect specific virus infection limits Arbovirus replication in Aedes mosquito cells. Virology 2018, 518, 406–413. [Google Scholar] [CrossRef]
- Baidaliuk, A.; Miot, E.F.; Lequime, S.; Moltini-Conclois, I.; Delaigue, F.; Dabo, S.; Dickson, L.B.; Aubry, F.; Merkling, S.H.; Cao-Lormeau, V.-M.; et al. Cell-Fusing Agent Virus Reduces Arbovirus Dissemination in Aedes aegypti Mosquitoes In Vivo. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Patterson, E.I.; Kautz, T.F.; Contreras-Gutierrez, M.A.; Guzman, H.; Tesh, R.B.; Hughes, G.L.; Forrester, N.L. Negeviruses Reduce Replication of Alphaviruses during Coinfection. J. Virol. 2021, 95, e0043321. [Google Scholar] [CrossRef]
- Nanfack-Minkeu, F.; Mitri, C.; Bischoff, E.; Belda, E.; Casademont, I.; Vernick, K.D. Interaction of RNA viruses of the natural virome with the African malaria vector, Anopheles coluzzii. Sci. Rep. 2019, 9, 6319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegde, S.; Rasgon, J.L.; Hughes, G.L. The microbiome modulates arbovirus transmission in mosquitoes. Curr. Opin. Virol. 2015, 15, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, M.; Nilsson, L.K.J.; Brunius, C.; Dabiré, R.K.; Hopkins, R.; Terenius, O. Bacterial associations reveal spatial population dynamics in Anopheles gambiae mosquitoes. Sci. Rep. 2016, 6, 22806. [Google Scholar] [CrossRef] [Green Version]
- Gimonneau, G.; Tchioffo, M.T.; Abate, L.; Boissière, A.; Awono-Ambéné, P.H.; Nsango, S.E.; Christen, R.; Morlais, I. Composition of Anopheles coluzzii and Anopheles gambiae microbiota from larval to adult stages. Infect. Genet. Evol. 2014, 28, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Coon, K.L.; Vogel, K.J.; Brown, M.R.; Strand, M.R. Mosquitoes rely on their gut microbiota for development. Mol. Ecol. 2014, 23, 2727–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuesta, E.B.; Coulibaly, B.; Bukhari, T.; Eiglmeier, K.; Kone, R.; Coulibaly, M.B.; Zongo, S.; Barry, M.; Gneme, A.; Guelbeogo, W.M.; et al. Comprehensive Ecological and Geographic Characterization of Eukaryotic and Prokaryotic Microbiomes in African Anopheles. Front. Microbiol. 2021, 12, 635772. [Google Scholar] [CrossRef]
- Boissière, A.; Tchioffo, M.T.; Bachar, D.; Abate, L.; Marie, A.; Nsango, S.E.; Shahbazkia, H.R.; Awono-Ambene, P.H.; Levashina, E.A.; Christen, R.; et al. Midgut Microbiota of the Malaria Mosquito Vector Anopheles gambiae and Interactions with Plasmodium falciparum Infection. PLoS Pathog. 2012, 8, e1002742. [Google Scholar] [CrossRef] [Green Version]
- Osei-Poku, J.; Mbogo, C.M.; Palmer, W.J.; Jiggins, F.M. Deep sequencing reveals extensive variation in the gut microbiota of wild mosquitoes from Kenya. Mol. Ecol. 2012, 21, 5138–5150. [Google Scholar] [CrossRef]
- Dong, Y.; Manfredini, F.; Dimopoulos, G. Implication of the Mosquito Midgut Microbiota in the Defense against Malaria Parasites. PLoS Pathog. 2009, 5, e1000423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pumpuni, C.B.; Beier, M.S.; Nataro, J.P.; Guers, L.D.; Davis, J.R. Plasmodium falciparum: Inhibition of Sporogonic Development in Anopheles stephensi by Gram-Negative Bacteria. Exp. Parasitol. 1993, 77, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Gendrin, M.; Rodgers, F.H.; Yerbanga, R.S.; Ouédraogo, J.B.; Basáñez, M.-G.; Cohuet, A.; Christophides, G.K. Antibiotics in ingested human blood affect the mosquito microbiota and capacity to transmit malaria. Nat. Commun. 2015, 6, 5921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitri, C.; Bischoff, E.; Cuesta, E.B.; Volant, S.; Ghozlane, A.; Eiglmeier, K.; Holm, I.; Dieme, C.; Brito-Fravallo, E.; Guelbeogo, W.M.; et al. Leucine-Rich Immune Factor APL1 Is Associated with Specific Modulation of Enteric Microbiome Taxa in the Asian Malaria Mosquito Anopheles stephensi. Front. Microbiol. 2020, 11, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia Symbiont in Aedes aegypti Limits Infection with Dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [Green Version]
- Agyekum, T.P.; Botwe, P.K.; Arko-Mensah, J.; Issah, I.; Acquah, A.A.; Hogarh, J.N.; Dwomoh, D.; Robins, T.G.; Fobil, J.N. A Systematic Review of the Effects of Temperature on Anopheles Mosquito Development and Survival: Implications for Malaria Control in a Future Warmer Climate. Int. J. Environ. Res. Public Health 2021, 18, 7255. [Google Scholar] [CrossRef] [PubMed]
- Faiman, R.; Solon-Biet, S.; Sullivan, M.; Huestis, D.L.; Lehmann, T. The contribution of dietary restriction to extended longevity in the malaria vector Anopheles coluzzii. Parasites Vectors 2017, 10, 156. [Google Scholar] [CrossRef] [Green Version]
- Goindin, D.; Delannay, C.; Ramdini, C.; Gustave, J.; Fouque, F. Parity and Longevity of Aedes aegypti According to Temperatures in Controlled Conditions and Consequences on Dengue Transmission Risks. PLoS ONE 2015, 10, e0135489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mordecai, E.A.; Cohen, J.M.; Evans, M.V.; Gudapati, P.; Johnson, L.R.; Lippi, C.A.; Miazgowicz, K.; Murdock, C.C.; Rohr, J.R.; Ryan, S.J.; et al. Detecting the impact of temperature on transmission of Zika, dengue, and chikungunya using mechanistic models. PLoS Negl. Trop. Dis. 2017, 11, e0005568. [Google Scholar] [CrossRef] [Green Version]
- Heitmann, A.; Jansen, S.; Lühken, R.; Helms, M.; Pluskota, B.; Becker, N.; Kuhn, C.; Schmidt-Chanasit, J.; Tannich, E. Experimental risk assessment for chikungunya virus transmission based on vector competence, distribution and temperature suitability in Europe. Eurosurveillance 2018, 23, 1800033. [Google Scholar] [CrossRef] [Green Version]
- Bellone, R.; Failloux, A.-B. The Role of Temperature in Shaping Mosquito-Borne Viruses Transmission. Front. Microbiol. 2020, 11, 584846. [Google Scholar] [CrossRef]
- Mercier, A.; Obadia, T.; Carraretto, D.; Velo, E.; Gabiane, G.; Bino, S.; Vazeille, M.; Gasperi, G.; Dauga, C.; Malacrida, A.R.; et al. Impact of temperature on dengue and chikungunya transmission by the mosquito Aedes albopictus. Sci. Rep. 2022, 12, 6973. [Google Scholar] [CrossRef]
- Olayemi, I.; Danlami, G.; Isah, B.; Odeyemi, O.; Ukubuiwe, A.; Mustapha, O. Indoor Behaviour Responses of the Principal Malaria Vector, Anopheles gambiae (Diptera: Culicidae), in Relation to Micro-climatic Conditions in Minna, North Central Nigeria. Res. J. Parasitol. 2011, 6, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.C.; Ho, B.C.; Chan, K.L. Aedes aegypti (L.) and Aedes albopictus (Skuse) in Singapore City. Bull World Health Organ 1971, 44, 651–657. [Google Scholar]
- Schmidt, C.A.; Comeau, G.; Monaghan, A.J.; Williamson, D.J.; Ernst, K.C. Effects of desiccation stress on adult female longevity in Aedes aegypti and Ae. albopictus (Diptera: Culicidae): Results of a systematic review and pooled survival analysis. Parasites Vectors 2018, 11, 267. [Google Scholar] [CrossRef] [PubMed]
- Shil, P.; Kothawale, D.R.; Sudeep, A.B. Rainfall and Chikungunya incidences in India during 2010–2014. Virusdisease 2018, 29, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Hery, L.; Guidez, A.; Durand, A.-A.; Delannay, C.; Normandeau-Guimond, J.; Reynaud, Y.; Issaly, J.; Goindin, D.; Legrave, G.; Gustave, J.; et al. Natural Variation in Physicochemical Profiles and Bacterial Communities Associated with Aedes aegypti Breeding Sites and Larvae on Guadeloupe and French Guiana. Microb. Ecol. 2021, 81, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, R.P.; Dessie, Z.G.; Noreddin, A.; El Zowalaty, M.E. Systematic Review of Important Viral Diseases in Africa in Light of the ‘One Health’ Concept. Pathogens 2020, 9, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngono, A.E.; Shresta, S. Immune Response to Dengue and Zika. Annu. Rev. Immunol. 2018, 36, 279–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poddar, S.; Hyde, J.L.; Gorman, M.J.; Farzan, M.; Diamond, M.S. The Interferon-Stimulated Gene IFITM3 Restricts Infection and Pathogenesis of Arthritogenic and Encephalitic Alphaviruses. J. Virol. 2016, 90, 8780–8794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellington, D.; Laurenson-Schafer, H.; Abdel-Haq, A.; Dong, T. IFITM3: How genetics influence influenza infection demographically. Biomed. J. 2019, 42, 19–26. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| A. Viral comparisons | |||
| ONNV | CHIKV | ||
| Viral classification | Family | Togaviridae | |
| Genus | Alphavirus | ||
| Antigenic complex group | Semliki forest complex | ||
| Viral genome | Group | Baltimore group IV | |
| Type | ssRNA (+) | ||
| Nucleic sequences | 76.48% identity with 93% of coverage | ||
| Mutation rate | CHIKV: average estimation of 4.33 × 10−4 nucleotide substitutions per site per year [9] ONNV: no estimation | ||
| Viral cycle in cells | similar; same viral proteins harbor similar functions | ||
| Geographic distribution | Sub-Saharan region (Africa) | ||
| Protein sequence | Opal-to-Arg codon between nsP3 and nsP4 | Equilibrium of Opal-to-Arg codon [10,11] | Minority of Arg codon [12,13] |
| Geographic distribution | Restricted to Africa | Found in four continents (America, Africa, Europe and Asia | |
| Mortality rate in patients | not reported | 0,1% of cases [14] | |
| Vectors used for transmission | Anopheles funestus Anopheles gambiae Aedes aegypti [15] Mansonia uniformis [11] | Aedes aegypt Aedes albopictus | |
| B. Mosquito vector comparisons | |||
| Anopheles gambiae | Aedes aegypti | ||
| Behavioral traits | Blood feeding preferences | Anthropophilic | |
| Blood feeding time | Crepuscular or nocturnal | ||
| Blood feeding places | Endophilic | ||
| Developmental stages | Laying sites | Clear, unpolluted, fresh or salt water | Walls of water containers |
| Larval habitats | Rice fields or flooded areas | Tires, bowls, cups, natural basins | |
| Climate preference | Predominant during dry season [16] | Predominant beginning of the rainy season [17] | |
| Genome | Genome size | 278 Mb [18] | 1380 Mb [19] |
| Chromosome length | Shorter | 2.3 times longer [20] | |
| Transposable element composition | 16% of the genome [18] | 50% of the genome [21] | |
| Transposable element localisation | Pericentromeric heterochromatin | Euchromatin | |
| Protein orthology | 67% [19] | ||
| Number of orthologs | 2000 [19] | ||
| Geographic distribution | Sahara, Northern Europe, Northern Asia | Northern Africa, Australia | |
| America, South and Sub-Saharan Africa, West and East Asia Western Europe | |||
| Pathogens transmission | Parasites | Plasmodium spp. | / |
| Wuchereria bancrofti [22,23] | |||
| Viruses | ONNV | Flaviviruses Alphaviruses Phleboviruses Orthobuynyaviruses | |
| C. Vectorial system comparison | |||
| ONNV/Anopheles gambiae | CHIKV/Aedes aegypti | ||
| Extrinsic incubation period | Passage of the Midgut barrier | 3 days post infection [24] | Before 2 days post infection [25,26,27,28] |
| In salivary glands | 7 days post infection [24] | 2 to 3 days post infection [25,26,27,28] | |
| Receptors for viral entry | Putative receptors | 230 kDa Cadherin and Rab5 ortholog could be involved in ONNV entry [29] | 38 kDa and 60 kDa protein at the membrane brush border of the midgut [30] |
| Lectins and prohibitins [29,30,31] | |||
| Attachment factors | Glycoaminoglycans [31] | ||
| Antiviral immunity | RNA interference | siRNA, piRNA and miRNA | |
| Toll pathway | Inhibited by virus Repress viral dissemination [24] | Inhibited by virus and no antiviral response [32] | |
| IMD pathway | Inhibited by virus Protective against ONNV midgut infection (with Rel2-F) [24] | Inhibited by virus and no antiviral response [32] | |
| JAK/STAT pathway | Inhibited by virus Protective against ONNV midgut infection [24] | Inhibited by virus and no antiviral response [32] | |
| JNK pathway | No evidence | Antiviral response [33] | |
| Complement-like pathway | APL1A, APL1C and LRIM4 [24] | TEP20 [33] | |
| AMP | / | Cecropin-like peptide [34] | |
| Microbiota | These viruses require enteric microbiome [24,35,36] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cottis, S.; Blisnick, A.A.; Failloux, A.-B.; Vernick, K.D. Determinants of Chikungunya and O’nyong-Nyong Virus Specificity for Infection of Aedes and Anopheles Mosquito Vectors. Viruses 2023, 15, 589. https://doi.org/10.3390/v15030589
Cottis S, Blisnick AA, Failloux A-B, Vernick KD. Determinants of Chikungunya and O’nyong-Nyong Virus Specificity for Infection of Aedes and Anopheles Mosquito Vectors. Viruses. 2023; 15(3):589. https://doi.org/10.3390/v15030589
Chicago/Turabian StyleCottis, Solène, Adrien A. Blisnick, Anna-Bella Failloux, and Kenneth D. Vernick. 2023. "Determinants of Chikungunya and O’nyong-Nyong Virus Specificity for Infection of Aedes and Anopheles Mosquito Vectors" Viruses 15, no. 3: 589. https://doi.org/10.3390/v15030589