
Comparing Control Intervention Scenarios for Raccoon Rabies in Southern Ontario between 2015 and 2025
 , ,
, ,
Abstract
:1. Introduction
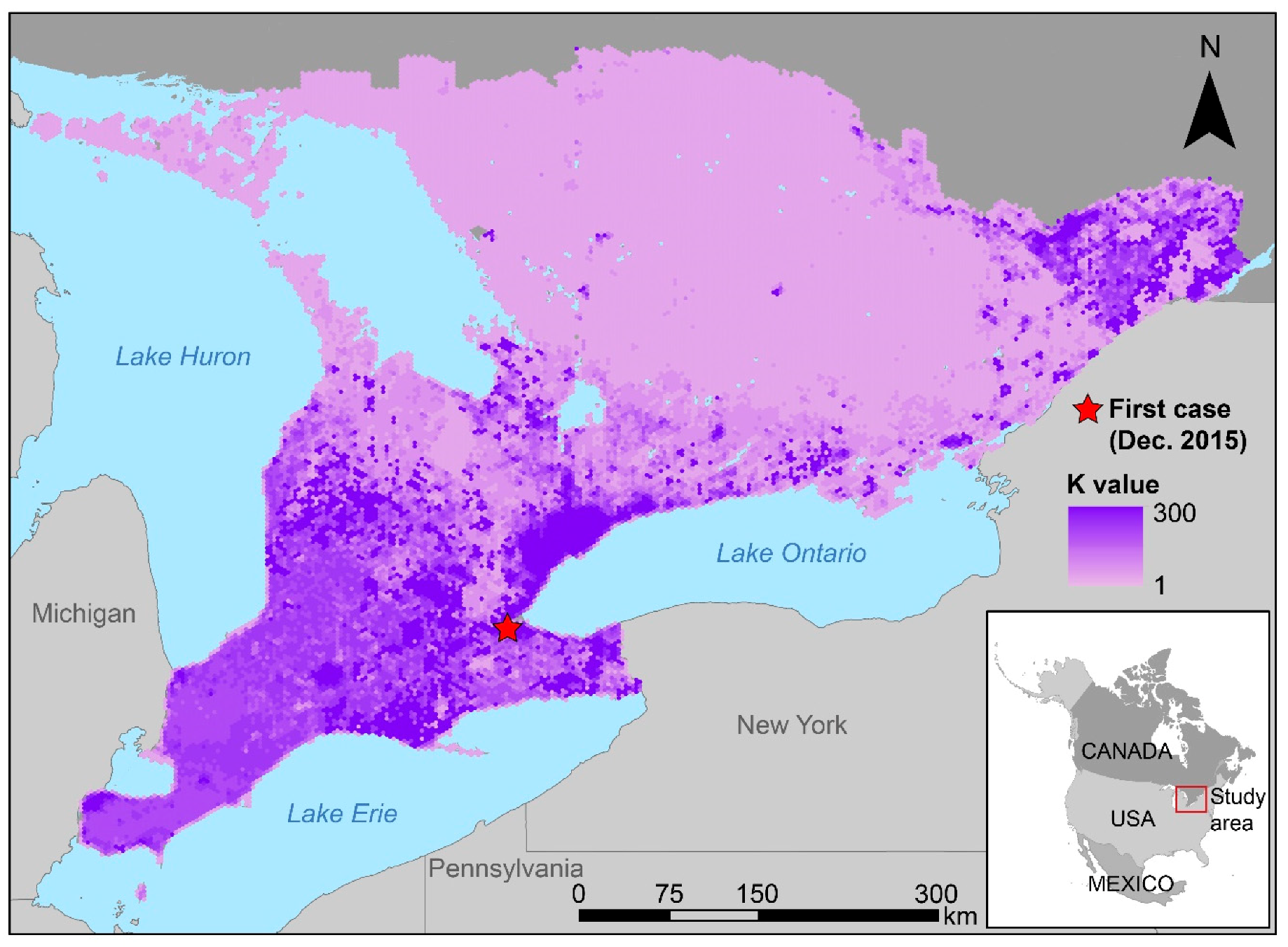
2. Materials and Methods
2.1. Agent-Based Model
2.2. Overview
2.2.1. Purpose
2.2.2. Entities, State Variables, and Scales
2.2.3. Process Overview and Scheduling
2.3. Design Concepts
2.3.1. Emergence
2.3.2. Interactions
2.3.3. Stochasticity
2.3.4. Collectives
2.3.5. Observation
2.4. Details
2.4.1. Initialization
2.4.2. Input
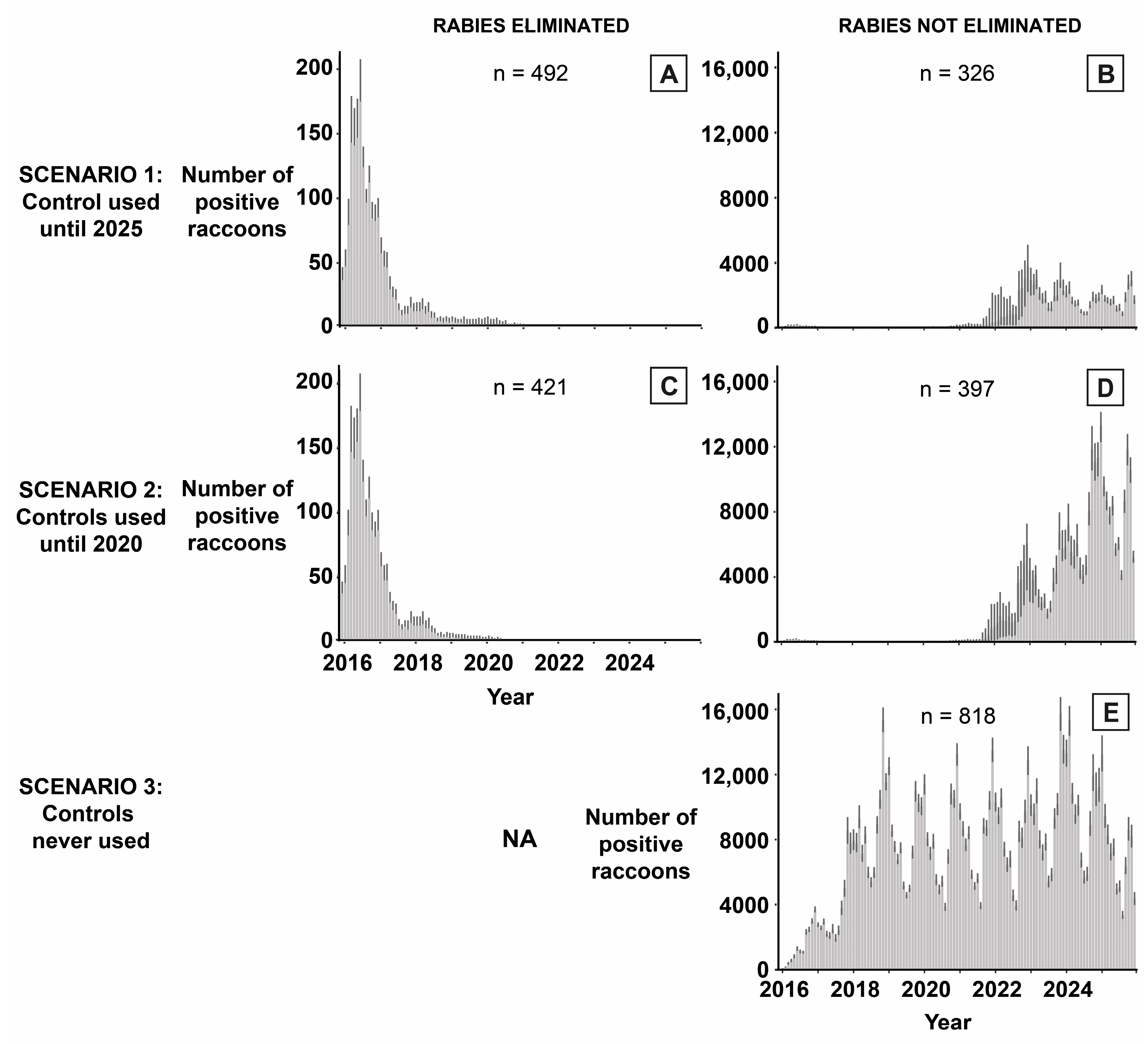
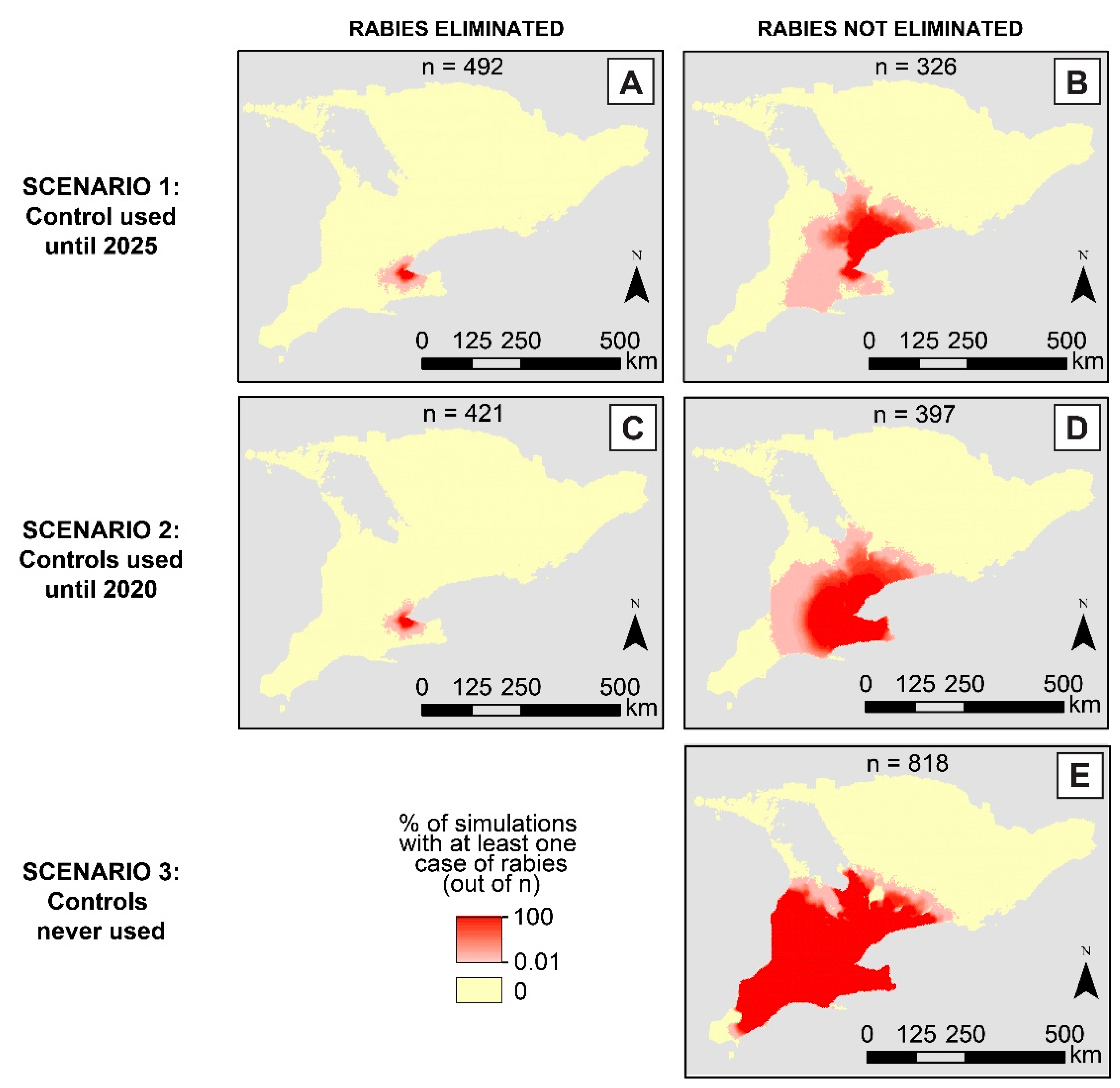
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fooks, A.R.; Banyard, A.C.; Horton, D.L.; Johnson, N.; McElhinney, L.M.; Jackson, A.C. Current status of rabies and prospects for elimination. Lancet 2014, 384, 1389–1399. [Google Scholar] [CrossRef] [PubMed]
- Shwiff, S.A.; Nunan, C.P.; Kirkpatrick, K.N.; Shwiff, S.S. A retrospective economic analysis of the Ontario red fox oral rabies vaccination programme. Zoonoses Public Health 2011, 58, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Nadin-Davis, S.; Buchanan, T.; Nituch, L.; Fehlner-Gardiner, C. A long-distance translocation initiated an outbreak of raccoon rabies in Hamilton, Ontario, Canada. PLOS Negl. Trop. Dis. 2020, 14, e0008113. [Google Scholar] [CrossRef] [PubMed]
- Rosatte, R.; Ryckman, M.; Ing, K.; Proceviat, S.; Allan, M.; Bruce, L.; Donovan, D.; Davies, C.D. Density, movements, and survival of raccoons in Ontario, Canada: Implications for disease spread and management. J. Mammal. 2010, 91, 122–135. [Google Scholar] [CrossRef]
- Trewby, H.; Nadin-Davis, S.A.; Real, L.A.; Biek, R. Processes underlying rabies virus incursions across US-Canada border as revealed by whole-genome phylogeography. Emerg. Infect. Dis. 2017, 23, 1454–1461. [Google Scholar] [CrossRef] [PubMed]
- Lobo, D.; DeBenedet, C.; Fehlner-Gardiner, C.; Nadin-Davis, S.; Anderson, M.; Buchanan, T.; Middel, K.; Hopkins, J. Raccoon rabies outbreak in Hamilton, Ontario: A progress report. Can. Commun. Dis. Rep. 2018, 44, 116–121. [Google Scholar] [CrossRef]
- Rees, E.E.; Pond, B.A.; Tinline, R.R.; Belanger, D. Modelling the effect of landscape heterogeneity on the efficacy of vaccination for wildlife infectious disease control. J. Appl. Ecol. 2013, 50, 881–891. [Google Scholar] [CrossRef]
- Riley, S. Large-scale spatial-transmission models of infectious disease. Science 2007, 316, 1298–1301. [Google Scholar] [CrossRef]
- Viard, F.; Acheson, E.S.; Allibert, A.; Sauvé, C.C.; Leighton, P.A. SamPy: A New Python Library for Stochastic Spatial Agent-Based Modelling in the Epidemiology of Infectious Diseases. Preprint 2022. Available online: https://www.preprints.org/manuscript/202211.0556/v2 (accessed on 5 January 2023).
- Tinline, R.R. The Ontario Rabies Model Guide; Queen’s University: Kingston, ON, Canada, 2007. [Google Scholar]
- Rees, E.E.; Pond, B.A.; Cullingham, C.I.; Tinline, R.R.; Ball, D.; Kyle, C.J.; White, B.N. Landscape modelling spatial bottlenecks: Implications for raccoon rabies disease spread. Biol. Lett. 2009, 5, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Rees, E.E.; Pond, B.A.; Cullingham, C.I.; Tinline, R.; Ball, D.; Kyle, C.J.; White, B.N. Assessing a landscape barrier using genetic simulation modelling: Implications for raccoon rabies management. Prev. Vet. Med. 2008, 86, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Grimm, V.; Berger, U.; Bastiansen, F.; Eliassen, S.; Ginot, V.; Giske, J.; Goss-Custard, J.; Grand, T.; Heinz, S.K.; Huse, G. A standard protocol for describing individual-based and agent-based models. Ecol. Model. 2006, 198, 115–126. [Google Scholar] [CrossRef]
- Bigler, W.J.; Hoff, G.L.; Smith, J.S.; McLean, R.G.; Trevino, H.A.; Ingwersen, J. Persistence of rabies antibody in free-ranging raccoons. J. Infect. Dis. 1983, 148, 610. [Google Scholar] [CrossRef] [PubMed]
- Gold, S.; Donnelly, C.A.; Nouvellet, P.; Woodroffe, R. Rabies virus-neutralising antibodies in healthy, unvaccinated individuals: What do they mean for rabies epidemiology? PLOS Negl Trop Dis. 2020, 14, e0007933. [Google Scholar] [CrossRef]
- Sobey, K.G.; Rosatte, R.; Bachmann, P.; Buchanan, T.; Bruce, L.; Donovan, D.; Brown, L.; Davies, J.C.; Fehlner-Gardiner, C.; Wandeler, A. Field evaluation of an inactivated vaccine to control raccoon rabies in Ontario, Canada. J. Wildl. Dis. 2010, 46, 818–831. [Google Scholar] [CrossRef]
- Tinline, R.R.; Ball, D.; Broadfoot, J.; Pond, B.A. The Ontario Rabies Model; Queen’s University: Kingston, ON, Canada, 2009. [Google Scholar]
- Hongoh, V.; Rees, E.; Massé, A.; Leighton, P.A. Modélisation du Risque de Réincursion de la Rage du Raton Laveur au Québec; Université de Montréal: Montréal, QC, Canada, 2020. [Google Scholar]
- Slate, D.; Saidy, B.D.; Simmons, A.; Nelson, K.M.; Davis, A.; Algeo, T.P.; Elmore, S.A.; Chipman, R.B. Rabies management implications based on raccoon population density indexes. J. Wildl. Manag. 2020, 84, 877–890. [Google Scholar] [CrossRef]
- Rees, E.E.; Belanger, D.; Lelievre, F.; Cote, N.; Lambert, L. Targeted surveillance of raccoon rabies in Quebec, Canada. J. Wildl. Manag. 2011, 75, 1406–1416. [Google Scholar] [CrossRef]
- Hirsch, B.T.; Reynolds, J.J.H.; Gehrt, S.D.; Craft, M.E. Which mechanisms drive seasonal rabies outbreaks in raccoons? A test using dynamic social network models. J. Appl. Ecol. 2016, 53, 804–813. [Google Scholar] [CrossRef]
- Kirby, J.D.; Chipman, R.B.; Nelson, K.M.; Rupprecht, C.E.; Blanton, J.D.; Algeo, T.P.; Slate, D. Enhanced rabies surveillance to support effective oral rabies vaccination of raccoons in the eastern United States. Trop. Med. Inf. Dis. 2017, 2, 34. [Google Scholar] [CrossRef]
- Nadin-Davis, S.A.; Fu, Q.; Trewby, H.; Biek, R.; Johnson, R.H.; Real, L. Geography but not alternative host species explain the spread of raccoon rabies virus in Vermont. Epidemiol. Infect. 2018, 146, 1977–1986. [Google Scholar] [CrossRef]
- Johnson, S.R.; Slate, D.; Nelson, K.M.; Davis, A.J.; Mills, S.A.; Forbes, J.T.; VerCauteren, K.C.; Gilbert, A.T.; Chipman, R.B. Serological responses of raccoons and striped skunks to Ontario rabies vaccine bait in West Virginia during 2012-2016. Viruses 2021, 13, 157. [Google Scholar] [CrossRef]
- Dodds, W.J.; Larson, L.J.; Christine, K.L.; Schultz, R.D. Duration of immunity after rabies vaccination in dogs: The Rabies Challenge Fund research study. Can. J. Vet. Res. 2020, 84, 153–158. [Google Scholar]
- Rosatte, R.C.; Power, M.J.; MacInnes, C.D.; Lawson, K.F. Rabies control for urban foxes, skunks, and raccoons. Proc. Vertebr. Pest Conf. 1990, 14, 72. [Google Scholar]
- Brown, L.J.; Rosatte, R.C.; Fehlner-Gardiner, C.; Knowles, M.K.; Bachmann, P.; Davies, J.C.; Wandeler, A.; Sobey, K.; Donovan, D. Immunogenicity and efficacy of two rabies vaccines in wild-caught, captive raccoons. J. Wildl. Dis. 2011, 47, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.J.; Rosatte, R.C.; Fehlner-Gardiner, C.; Taylor, J.S.; Davies, J.C.; Donovan, D. Immune response and protection in raccoons (Procyon lotor) following consumption of baits containing ONRAB®, a human adenovirus rabies glycoprotein recombinant vaccine. J. Wildl. Dis. 2012, 48, 1010–1020. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, A.; Johnson, S.; Walker, N.; Wickham, C.; Beath, A.; VerCauteren, K. Efficacy of Ontario Rabies Vaccine Baits (ONRAB) against rabies infection in raccoons. Vaccine 2018, 36, 4919–4926. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Raccoon Population Growth Parameters † | |
|---|---|
| Definition of Parameter | Input Settings |
| Number of initial breeding pairs | 5 (5 males, 5 females) |
| Age of breeding raccoon agents | 52 weeks |
| Number of simulation years for population building | 150 years |
| Age of independence (i.e., age when raccoon agent becomes a juvenile) | 20 weeks |
| Age adulthood is reached | 75 weeks |
| Gestation period | 9 weeks |
| Week of the year in which females give birth | Week 18 |
| Annual mortality rates | Probability by age (0; 1; 2; 3; 4; 5; 6; 7) For both sexes (0.6; 0.4; 0.3; 0.3; 0.3; 0.6; 0.6; 0.6) |
| Mean litter size | 4 |
| Litter size variance | 1 |
| Probability density values for number of offspring produced in a litter | Number of offspring possible (0, 1, 2, 3, 4, 5, 6, 7, 8, 9) Probabilities (0.0001, 0.0044, 0.0539, 0.2416, 0.4000, 0.2416, 0.0539, 0.0044, 0.0001) |
| Litter sex ratio | 50 |
| Period of dispersion (week(s) of the year, where juveniles are defined as 20 weeks or older but under 75 weeks of age) | Juvenile males: 38–43 Juvenile females: 38–43 Adult males: 8–43 Adult females: 8–15 and 38–43 |
| Distance of dispersion | The probability of displacement by a particular distance (number of grid cells: probability) Juvenile males (0: 75.1, 1: 12.9, 2: 6.5, 3: 1.8, 4: 0.9, 5: 0.9, 6: 0.92, 7: 0, 8: 1, >=9: 0) Juvenile females (0: 90.8; 1: 4.07; 2: 1.4, 3: 0.3, 4: 0.7, 5: 0.3, 6: 1.02, 7: 0, 8: 1, >=9: 0) Adult males (0: 88.9; 1: 4.25; 2: 1.4; 3: 0.9; 4: 0.7, 5: 0.9, 6: 0, 7: 1, 8: 1, >=9: 0) Adult females (0: 92.3, 1: 3.06, 2: 1, 3: 0.9, 4: 0.4, 5: 0.7, 6: 0.73, >=7: 0) |
| Probability of leaving current grid cell to interact with raccoons in any of the six neighbouring grid cells | 22.2%, divided equally between neighbouring grid cells |
| Rabies dynamics parameters | |
| Chance of death from raccoon rabies | 95% |
| Transmission probability (the probability that pathogen transmission will occur given contact) | 3.5% |
| Level of initial infection | 5% |
| Week when rabies was introduced | Week 47 of 2015 |
| Incubation period | Number of weeks of infection: Probability of incubation period lasting x number of weeks (1: 1, 2: 5, 3: 5, 4: 10, 5: 15, 6: 20, 7: 15, 8: 10, 9: 5, 10: 5, 11: 5, 12: 2, 13: 1, 14: 1) |
| Infection period | 100% chance of infectious period lasting 1 week |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acheson, E.S.; Viard, F.; Buchanan, T.; Nituch, L.; Leighton, P.A. Comparing Control Intervention Scenarios for Raccoon Rabies in Southern Ontario between 2015 and 2025. Viruses 2023, 15, 528. https://doi.org/10.3390/v15020528
Acheson ES, Viard F, Buchanan T, Nituch L, Leighton PA. Comparing Control Intervention Scenarios for Raccoon Rabies in Southern Ontario between 2015 and 2025. Viruses. 2023; 15(2):528. https://doi.org/10.3390/v15020528
Chicago/Turabian StyleAcheson, Emily Sohanna, François Viard, Tore Buchanan, Larissa Nituch, and Patrick A. Leighton. 2023. "Comparing Control Intervention Scenarios for Raccoon Rabies in Southern Ontario between 2015 and 2025" Viruses 15, no. 2: 528. https://doi.org/10.3390/v15020528