
Live Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Undergoes an Abortive Replication Cycle in the TG Neurons following Latency Reactivation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Cells and Medium
2.3. Viruses
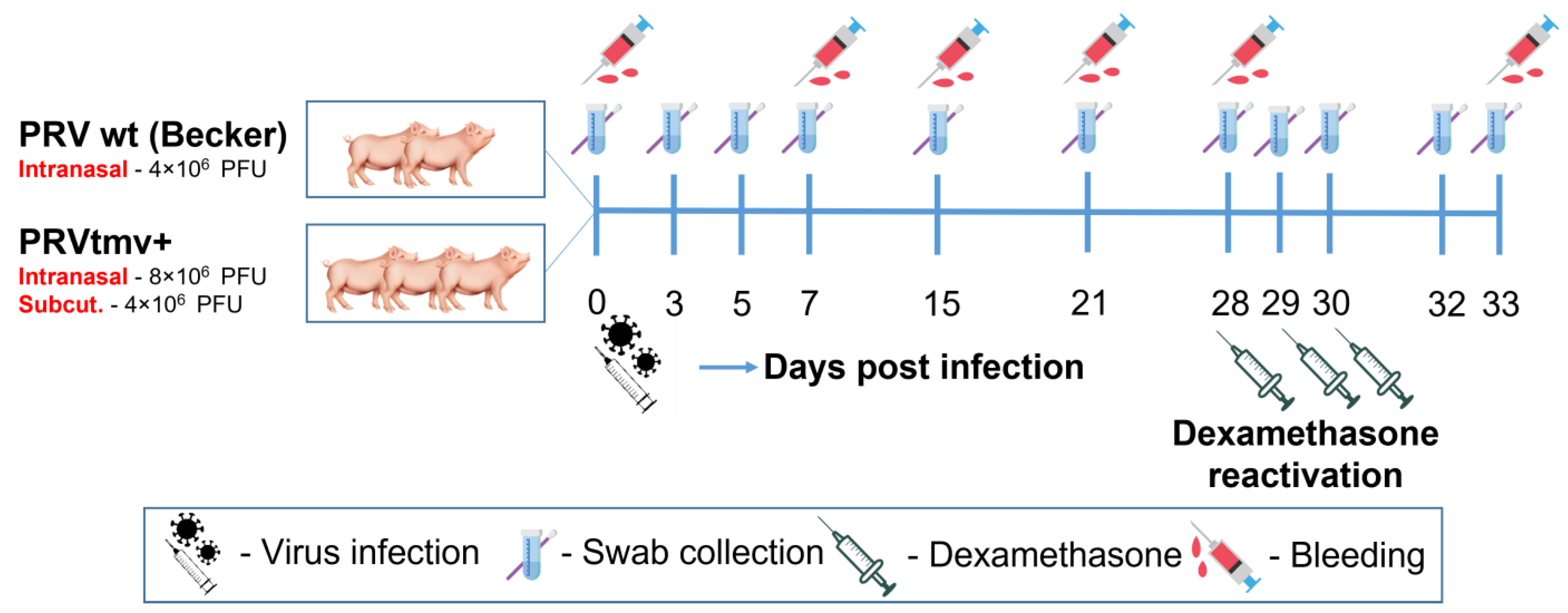
2.4. Animals and Experimental Design
2.5. Primary Virus Infection, Dexamethasone-Induced Latent Virus Reactivation, and Clinical Evaluation
2.6. Clinical Sample Collection and Processing
2.7. Euthanasia, Necropsy—Sample Collection and Processing
2.8. Serum Virus Neutralization Assay
2.9. DNA/RNA Isolation, cDNA Synthesis, and PRV-Specific Quantitative PCR (q-PCR)
2.10. Statistical Analysis
3. Results
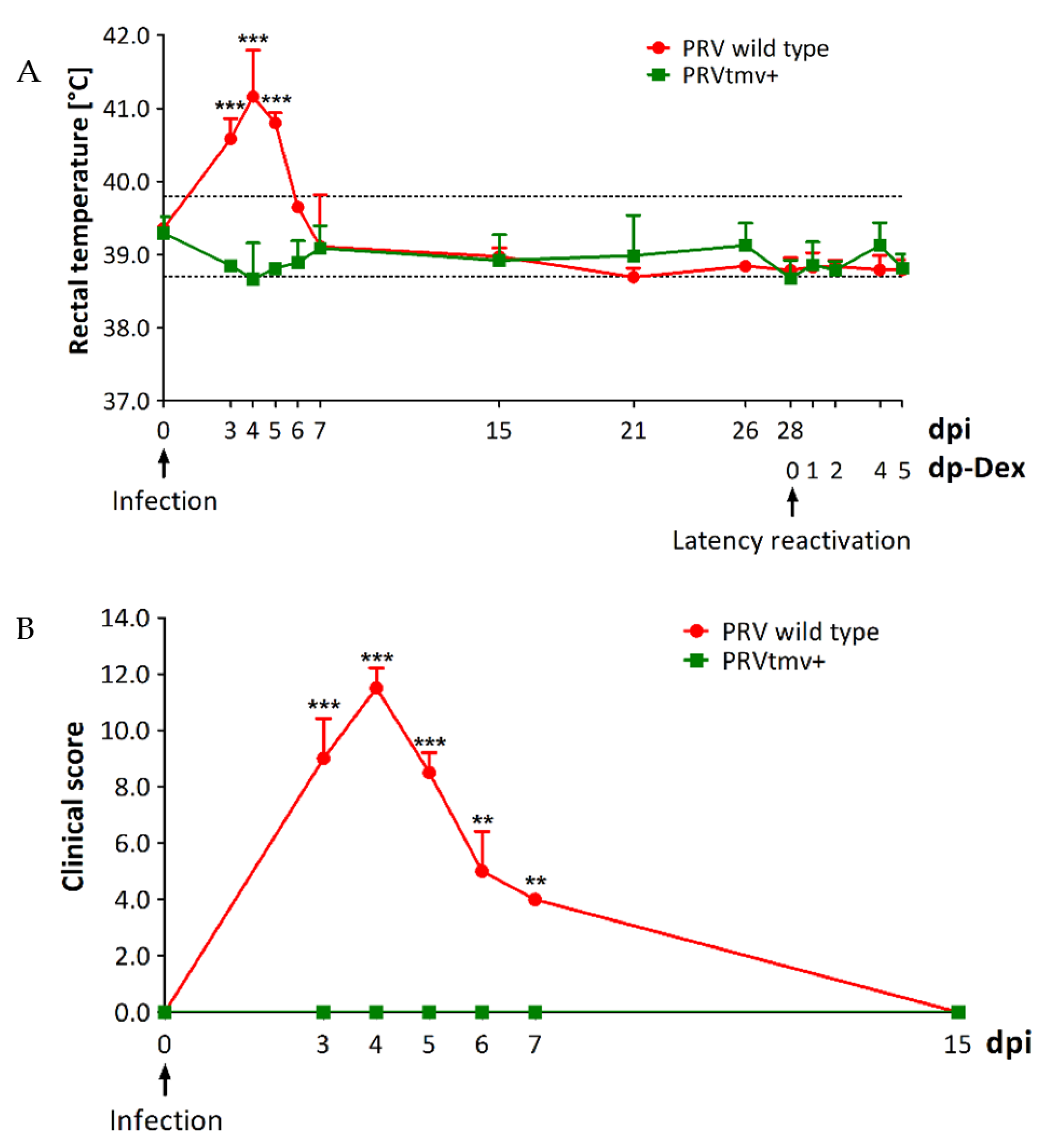
3.1. Clinical Evaluation
3.2. Nasal Virus Shedding following Intranasal (IN) Administration of PRV wt and PRVtmv+
3.3. Following Dex-Induced Reactivation, Only the PRV wt-Infected Pigs Shed the Virus in Their Nasal Secretions
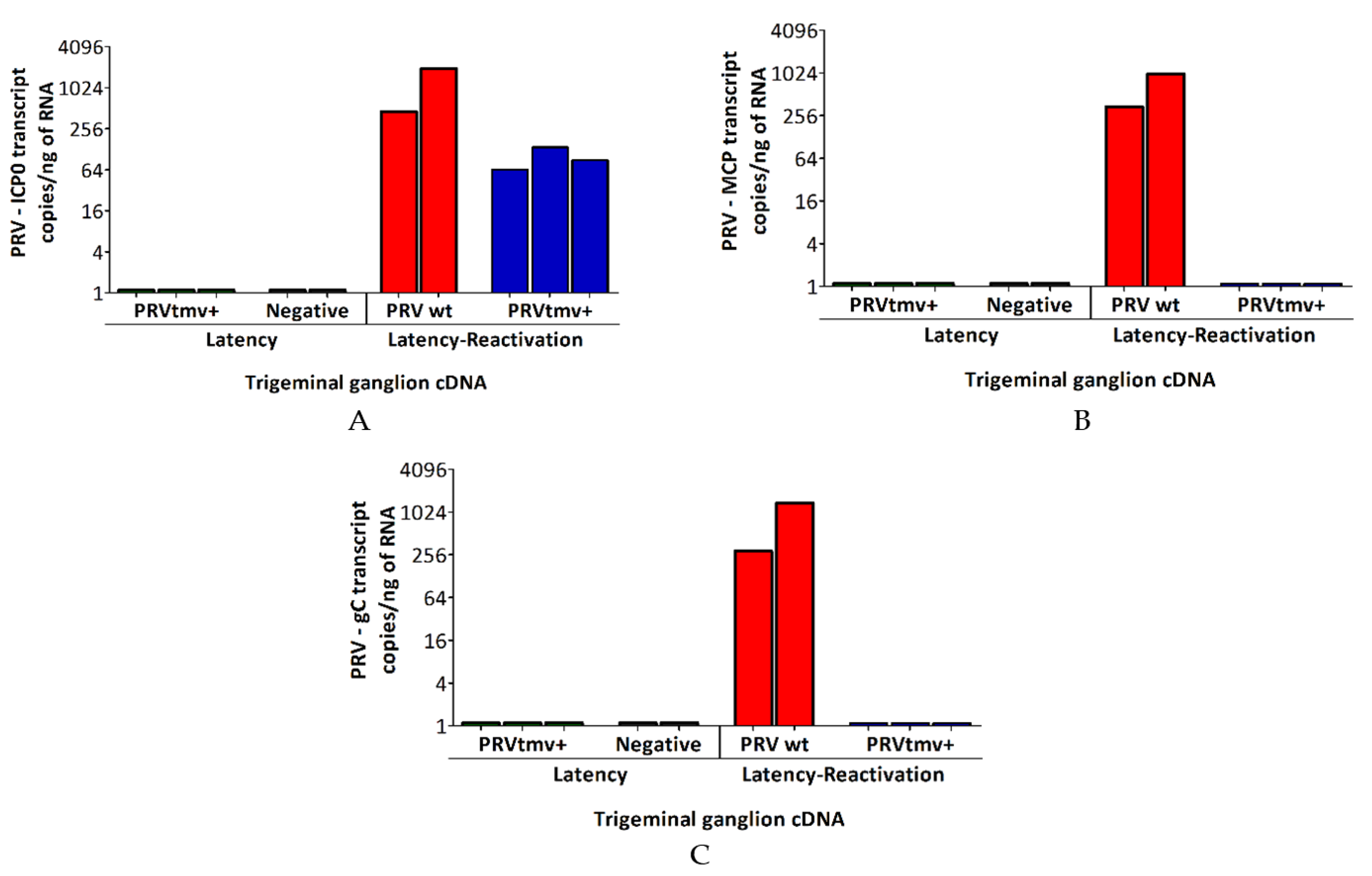
3.4. The PRV wt and PRVtmv+ Established Latency in the TG Neurons and Reactivated following Dex Injection, but Only PRV wt and Not PRVtmv+ Replicated in the TG Neurons
3.5. Following Dexamethasone (Dex)-Induced Latency Reactivation, Only PRV wt but Not PRVtmv+-Inoculated Pigs Had a Memory Serum Virus Neutralizing (SN) Antibody Response
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pavulraj, S.; Pannhorst, K.; Stout, R.W.; Paulsen, D.B.; Carossino, M.; Meyer, D.; Becher, P.; Chowdhury, S.I. A Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Protects Pigs against PCV2b Challenge and Induces Serum Neutralizing Antibody Response against CSFV. Vaccines 2022, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Bech-Nielsen, S.; Miller, G.Y.; Bowman, G.L.; Dodaro, S.J.; Orloski-Snider, K.A. Economic impact of an epizootic of pseudorabies in a commercial swine herd in Ohio, achieving test negative status and quarantine release by use of vaccination and test and removal. J. Am. Vet. Med. Assoc. 1992, 200, 1817–1823. [Google Scholar]
- Wheeler, J.G.; Osorio, F.A. Investigation of sites of pseudorabies virus latency, using polymerase chain reaction. Am. J. Vet. Res. 1991, 52, 1799–1803. [Google Scholar] [PubMed]
- Deng, J.; Wu, Z.; Liu, J.; Ji, Q.; Ju, C. The Role of Latency-Associated Transcripts in the Latent Infection of Pseudorabies Virus. Viruses 2022, 14, 1379. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, G.; Wan, C.; Li, Y.; Peng, L.; Fang, R.; Peng, Y.; Ye, C. A Comparison of Pseudorabies Virus Latency to Other alpha-Herpesvirinae Subfamily Members. Viruses 2022, 14, 1386. [Google Scholar] [CrossRef]
- Kit, S. Genetically engineered vaccines for control of Aujeszky’s disease (pseudorabies). Vaccine 1990, 8, 420–424. [Google Scholar] [CrossRef]
- Mettenleiter, T.C. Aujeszky’s Disease and the Development of the Marker/DIVA Vaccination Concept. Pathogens 2020, 9, 563. [Google Scholar] [CrossRef]
- van Oirschot, J.T. Diva vaccines that reduce virus transmission. J. Biotechnol. 1999, 73, 195–205. [Google Scholar] [CrossRef]
- Liu, Q.; Kuang, Y.; Li, Y.; Guo, H.; Zhou, C.; Guo, S.; Tan, C.; Wu, B.; Chen, H.; Wang, X. The Epidemiology and Variation in Pseudorabies Virus: A Continuing Challenge to Pigs and Humans. Viruses 2022, 14, 1463. [Google Scholar] [CrossRef]
- Maresch, C.; Lange, E.; Teifke, J.P.; Fuchs, W.; Klupp, B.; Muller, T.; Mettenleiter, T.C.; Vahlenkamp, T.W. Oral immunization of wild boar and domestic pigs with attenuated live vaccine protects against Pseudorabies virus infection. Vet. Microbiol. 2012, 161, 20–25. [Google Scholar] [CrossRef]
- Mettenleiter, T.C.; Schreurs, C.; Zuckermann, F.; Ben-Porat, T. Role of pseudorabies virus glycoprotein gI in virus release from infected cells. J. Virol. 1987, 61, 2764–2769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mettenleiter, T.C.; Zsak, L.; Kaplan, A.S.; Ben-Porat, T.; Lomniczi, B. Role of a structural glycoprotein of pseudorabies in virus virulence. J. Virol. 1987, 61, 4030–4032. [Google Scholar] [CrossRef] [PubMed]
- Viejo-Borbolla, A.; Munoz, A.; Tabares, E.; Alcami, A. Glycoprotein G from pseudorabies virus binds to chemokines with high affinity and inhibits their function. J. Gen. Virol. 2010, 91, 23–31. [Google Scholar] [CrossRef]
- Tenser, R.B.; Ressel, S.J.; Fralish, F.A.; Jones, J.C. The role of pseudorabies virus thymidine kinase expression in trigeminal ganglion infection. J. Gen. Virol. 1983, 64 Pt 6, 1369–1373. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.I.; Silva, E.; Pavulraj, S.; Ray, A.; Medina-Ramirez, E.; Gladue, D.P.; Stout, R.W.; Paulsen, D.B.; Borca, M. A PRV Vectored PCV2/CSFV-Sub Vaccine Protects Pigs against Lethal CSFV Challenge; Department of Pathobiological Sciences and Louisiana Animal Disease Diagnostic Laboratory, School of Veterinary Medicine, Louisiana State University: Baton Rouge, LA, USA, 2023; in preparation. [Google Scholar]
- Platt, K.B.; Mare, C.J.; Hinz, P.N. Differentiation of vaccine strains and field isolates of pseudorabies (Aujeszky’s disease) virus: Thermal sensitivity and rabbit virulence markers. Arch. Virol. 1979, 60, 13–23. [Google Scholar] [CrossRef]
- Chowdhury, S.I.; Pannhorst, K.; Sangewar, N.; Pavulraj, S.; Wen, X.; Stout, R.W.; Mwangi, W.; Paulsen, D.B. BoHV-1-Vectored BVDV-2 Subunit Vaccine Induces BVDV Cross-Reactive Cellular Immune Responses and Protects against BVDV-2 Challenge. Vaccines 2021, 9, 46. [Google Scholar] [CrossRef]
- Tham, K.M.; Motha, M.X.; Horner, G.W.; Ralston, J.C. Polymerase chain reaction amplification of latent Aujeszky’s disease virus in dexamethasone treated pigs. Arch. Virol. 1994, 136, 197–205. [Google Scholar] [CrossRef]
- Balasch, M.; Pujols, J.; Segales, J.; Plana-Duran, J.; Pumarola, M. Study of the persistence of Aujeszky’s disease (pseudorabies) virus in peripheral blood mononuclear cells and tissues of experimentally infected pigs. Vet. Microbiol. 1998, 62, 171–183. [Google Scholar] [CrossRef]
- Rziha, H.J.; Mettenleiter, T.C.; Ohlinger, V.; Wittmann, G. Herpesvirus (pseudorabies virus) latency in swine: Occurrence and physical state of viral DNA in neural tissues. Virology 1986, 155, 600–613. [Google Scholar] [CrossRef]
- Romero, C.H.; Meade, P.N.; Homer, B.L.; Shultz, J.E.; Lollis, G. Potential sites of virus latency associated with indigenous pseudorabies viruses in feral swine. J. Wildl. Dis. 2003, 39, 567–575. [Google Scholar] [CrossRef]
- Enquist, L.W. Exploiting circuit-specific spread of pseudorabies virus in the central nervous system: Insights to pathogenesis and circuit tracers. J. Infect. Dis. 2002, 186 (Suppl. 2), S209–S214. [Google Scholar] [CrossRef] [PubMed]
- Brockmeier, S.L.; Lager, K.M.; Mengeling, W.L. Comparison of in vivo reactivation, in vitro reactivation, and polymerase chain reaction for detection of latent pseudorabies virus infection in swine. J. Vet. Diagn. Invest. 1993, 5, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Mengeling, W.L. Virus reactivation in pigs latently infected with a thymidine kinase negative vaccine strain of pseudorabies virus. Arch. Virol. 1991, 120, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Daniel, G.R.; Sollars, P.J.; Pickard, G.E.; Smith, G.A. Pseudorabies Virus Fast Axonal Transport Occurs by a pUS9-Independent Mechanism. J. Virol. 2015, 89, 8088–8091. [Google Scholar] [CrossRef] [PubMed]
- Antinone, S.E.; Smith, G.A. Retrograde axon transport of herpes simplex virus and pseudorabies virus: A live-cell comparative analysis. J. Virol. 2010, 84, 1504–1512. [Google Scholar] [CrossRef] [PubMed]
- Shapira, M.; Homa, F.L.; Glorioso, J.C.; Levine, M. Regulation of the herpes simplex virus type 1 late (gamma 2) glycoprotein C gene: Sequences between base pairs −34 to +29 control transient expression and responsiveness to transactivation by the products of the immediate early (alpha) 4 and 0 genes. Nucleic Acids Res. 1987, 15, 3097–3111. [Google Scholar] [CrossRef]
- Andreau, K.; Lemaire, C.; Souvannavong, V.; Adam, A. Induction of apoptosis by dexamethasone in the B cell lineage. Immunopharmacology 1998, 40, 67–76. [Google Scholar] [CrossRef]
- Schang, L.M.; Kutish, G.F.; Osorio, F.A. Correlation between precolonization of trigeminal ganglia by attenuated strains of pseudorabies virus and resistance to wild-type virus latency. J. Virol. 1994, 68, 8470–8476. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Scoring Criteria | Rectal Temperature (°C) | Nasal Discharge | Lethargy | Dyspnea | Anorexia | Cough |
|---|---|---|---|---|---|---|
| Normal (0) | <39.7 | None | Normal in attitude | Normal breathing | Normal appetite | None |
| Mild (1) | 39.8–40.3 | Serous | Moves slowly, head down | Slight difficulty breathing | Slightly off feed | <3 occasional cough |
| Moderate (2) | 40.4–40.8 | Mucopurulent | Tends to lie down, moves reluctantly | Labored breathing | Moderately off feed | >3 repeated cough |
| Severe (3) | 40.9–41.1 | Copious mucopurulent | Difficult to stand, little response to stimuli | Very labored breathing | Not eating | NA |
| (4) | >41.2 | NA | NA | NA | NA | NA |
| Primer/Probe/ds-Gblock | Name | Sequence |
|---|---|---|
| PRV-ICP0 | Forward | 5′-atcccgtgctcctggataatctcg-3′ |
| Reverse | 5′-tccccgtcttcaactggctttatg-3′ | |
| Probe | 5′Fam-atgttgtccacgacggcctcgcgga-3′ Tamra | |
| ds-gblock | 5′-ggcctcggtcacgcgctggcggttcatcccgtgctcctggataatctcgacg Atgttgtccacgacggcctcgcggatggggtcgctctcgatgaccgtcgagacct gcccataaagccagttgaagacggggactctggggcgggcgcgagacccaga-3′ | |
| PRV–Major Capsid Protein | Forward | 5′-ccatccagtttgaggtgcag-3′ |
| Reverse | 5′-cgaggcgcttgatcatgtag-3′ | |
| Probe | 5′Fam-cccgtcgcgcgcgatcatcg-3′ Tamra | |
| ds-gblock | 5′-ctcagctacgtggccgagggcaccatccagtttgaggtgcagcagccgatg atcgcgcgcgacgggccgcacccggccgaccagcccgtgcacaactacatgatca agc gcctcgatcgccgctccctcaacgccgc-3′ | |
| PRV–Glycoprotein C | Forward | 5′-gtcgtccgcgactactacc-3′ |
| Reverse | 5′-tcacgttcaccacggagac-3′ | |
| Probe | 5′Fam-cgtccgcgaaccagcgcagg-3′ Tamra | |
| ds-gblock | 5′-agcccttccgggcggtgtgcgtcgtccgcgactactacccgcggcgcagcgtg cgcctgcgctggttcgcggacgagcacccggtggacgccgccttcgtgaccaa cagcaccgtggccgacgagctcgggcgccgcacgcgcgtctccgtggtgaac gtgacgcgcgcggacgtcccgggc-3′ | |
| Swine Glyceraldehyde 3-Phosphate Dehydrogenase (GAPDH) | Forward | 5′-atgacaacttcggcatcgtg-3′ |
| Reverse | 5′-ccatccacagtcttctgggt-3′ | |
| Probe | 5′Fam-accacagtccatgccatcactgcc-3′ Tamra | |
| ds-gblock | 5′-gcacccctggccaaggtcatccatgacaacttcggcatcgtggaaggactca tgaccacagtccatgccatcactgccacccagaagactgtggatggc ccctctgggaaacgtggcgt-3′ |
| Group | Animal # | Virus Isolation (PFU/mL) | PRV Genome Copies per 100 ng of DNA (Days Post-Infection) | |||||
|---|---|---|---|---|---|---|---|---|
| Day 3 | Day 5 | Day 3 | Day 5 | Day 15 | Day 21 | Day 28 | ||
| PRV wild-type | 2303 | 4.0 × 104 | 0 | 3.1 × 106 | 1.9 × 104 | 5.7 × 102 | 2.0 × 102 | 0 |
| 2306 | 7.6 × 104 | 0 | 6.7 × 106 | 4.1 × 104 | 3.6 × 102 | 3.8 × 102 | 0 | |
| PRVtmv+ | 2309 | 0 | 0 | 2.9 × 101 | 5.2 × 101 | 0 | 0 | 0 |
| 2312 | 0 | 0 | 2.4 × 102 | 1.4 × 102 | 0 | 0 | 0 | |
| 2315 | 0 | 0 | 8.1 × 101 | 1.8 × 102 | 0 | 0 | 0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavulraj, S.; Stout, R.W.; Paulsen, D.B.; Chowdhury, S.I. Live Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Undergoes an Abortive Replication Cycle in the TG Neurons following Latency Reactivation. Viruses 2023, 15, 473. https://doi.org/10.3390/v15020473
Pavulraj S, Stout RW, Paulsen DB, Chowdhury SI. Live Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Undergoes an Abortive Replication Cycle in the TG Neurons following Latency Reactivation. Viruses. 2023; 15(2):473. https://doi.org/10.3390/v15020473
Chicago/Turabian StylePavulraj, Selvaraj, Rhett W. Stout, Daniel B. Paulsen, and Shafiqul I. Chowdhury. 2023. "Live Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Undergoes an Abortive Replication Cycle in the TG Neurons following Latency Reactivation" Viruses 15, no. 2: 473. https://doi.org/10.3390/v15020473