1. Introduction
The African Swine Fever Virus (ASFV) is a large, cytoplasmically replicating DNA virus which causes a highly contagious and frequently fatal acute disease in domestic pigs [
1]. This disease, endemic in many African countries, entered Portugal and Spain in the past with disastrous economic consequences. More recently, in 2007, the virus entered Georgia, in the Caucasus, and has now spread essentially worldwide [
2]. There is no routinely used vaccine for ASFV and control relies on restricting animal movement and slaughter of infected pigs. This is particularly difficult in areas where both the wildlife reservoirs (warthog, bushpig and wild boar) and the tick vector of the virus are present, since these animal reservoirs may be persistently infected for many years. Development of an efficient and safe vaccine against ASFV would protect Europe from accidental entry of the virus, and remove a major pathogen of pigs from large endemic areas of Africa, where the increasing frequency of outbreaks has caused devastating losses to both rural and commercial farms.
Due to the acute nature of the infection and the complexity of the protective porcine anti-ASFV immune response [
3], the development of vaccine candidates has evolved slowly [
4]. So far, inactivated and subunit vaccines have shown poor results and a live, replicating virus vaccine attenuated by the deletion of virus host evasion gene(s) presents a practical solution with the advantage of stimulating both, innate and adaptive immunity.
As resistance to ASFV is known to correlate with a robust IFN response [
5], the virus is predicted to have evolved a number of counter-strategies to inhibit the IFN response, a prediction that has been verified [
6]. Therefore, deletion of virus genes inhibiting induction and/or impact of IFN is a logical choice for the construction of such an attenuated gene deletion virus vaccine. In our previous work we have shown that the protein encoded by the ASFV ORF I329L impairs the porcine, murine, and human TLR3-mediated cellular responses that lead to activation of both IFN-β promoter and NF-κB and thus induction of type I IFN and inflammatory cytokines [
7].
The ASFV I329L gene codes for a non-essential [
8] Type I membrane protein without any immediately obvious sequence homology, and which is highly conserved in both pathogenic and non-pathogenic virus strains. Functional assays, however, demonstrated that I329L not only inhibited the activation of the IFN-β promoter and NF-κB, but also inhibited the secretion of IFN [
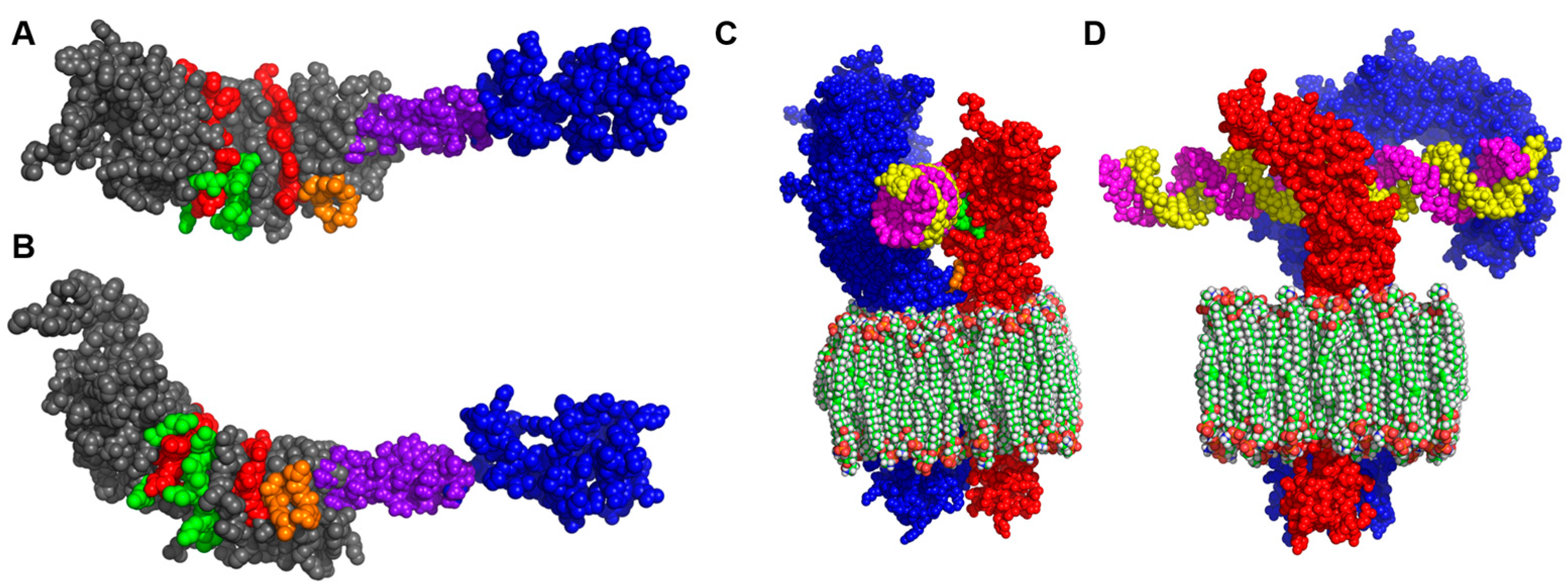
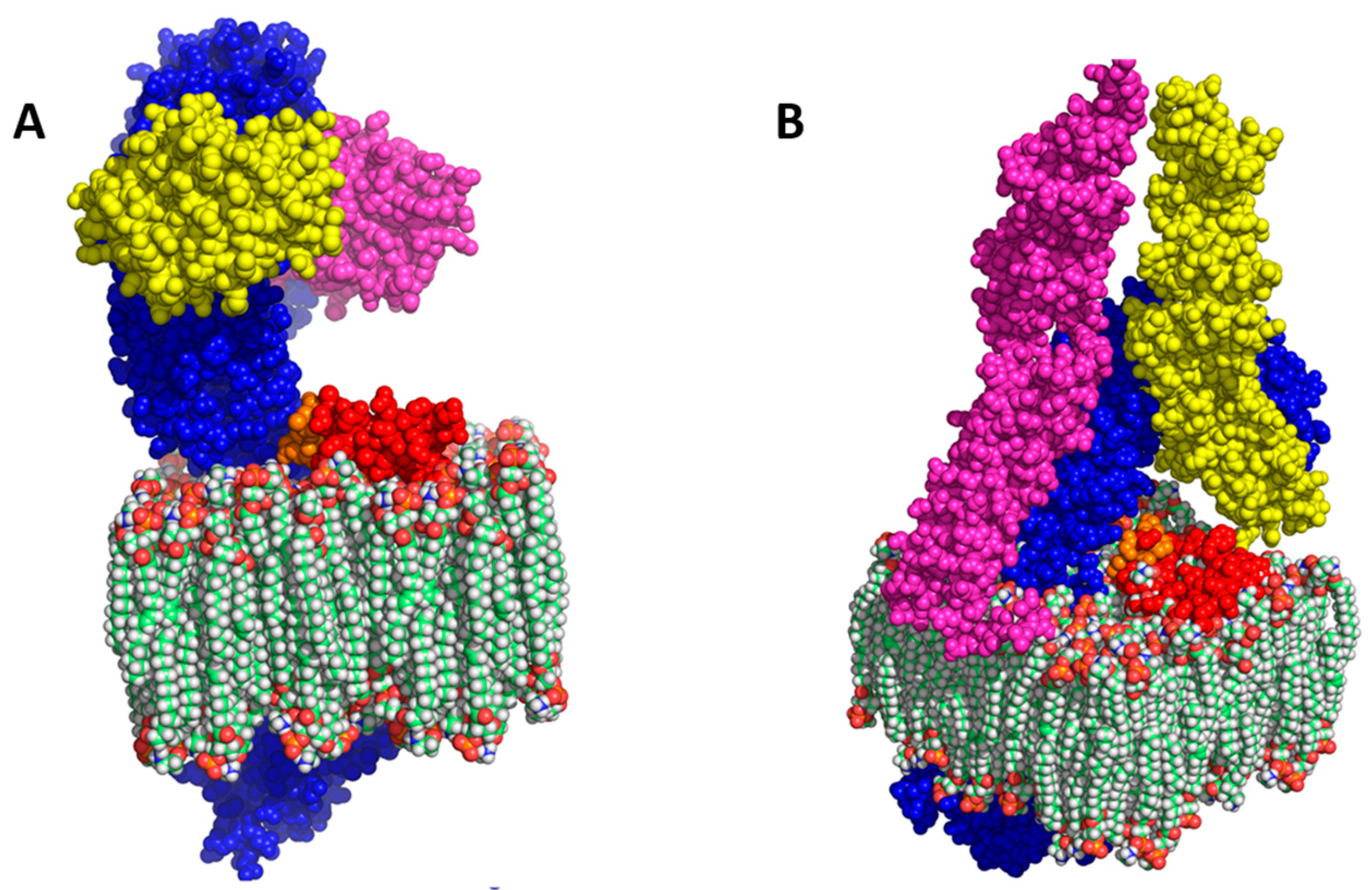
3]. More sophisticated bioinformatics structural modelling now suggests that (1) The extracellular domain (ECD) of I329L might serve as a dsRNA decoy or hinder TLR3 signalling, perhaps through formation of an I329L-TLR3 heterodimer, and (2) The intracellular domain (ICD) might interact with the downstream intracellular signalling intermediate TRIF [
9].
In this work, using a combination of biochemical and bioinformatics techniques, we confirm and extend the conclusion that the ASFV ORF I329L has evolved for the inhibition of innate immunity initiated through TLRs through two entirely different mechanisms. Protein structure simulation suggests that the ECD interferes with TLR signalling through a short sequence corresponding to the conserved TLR dimerization domain and termed the putative dimerization domain (PDD). Remarkably, both the full length and PDD constructs of I329L were demonstrated to inhibit activation not only of TLR3, but also TLR4, TLR5, TLR8 and TLR9. In addition to the inhibitory impact of the ECD, the ICD associates with the intracellular signalling intermediate TRIF, thereby impacting on both TLR3 and TLR4 signalling. Finally, bioinformatic analysis also suggested that both the ECD and ICD could inhibit TLR3 signalling. Thus, I329L offers potential as (1) An inhibitor of TLR responses and (2) A rational candidate for construction and testing of an I329L deletion mutant vaccine.
2. Materials and Methods
2.1. Homology Modelling of I329L
The sequences of I329L and its ECDTM and ICD domains were taken from the UniProt database (accession number A9JLD8) and investigated for possible homologies using Phyre2 (Protein Homology/analogY Recognition Engine V2.0 Webserver) [
10] to predict homologies with proteins of known structure. A preliminary set of models was obtained by use of the I-TASSER protein prediction server [
11] and used as a template for more accurate modelling. The resulting I329L structures were homology-modelled with Modeller 9.14 [
12] (
http://zhang.bioinformatics.ku.edu/I-TASSER, accessed on 31 December 2017) using the pl329L alignment with PDB templates obtained from Phyre2 and I-TASSER. The quality of the resulting models was assessed using PROCHECK v3.5.4 [
13], and the best quality models were used to determine possible protein complexes by superimposition with the structure of TLR3. The membrane was modelled using palmitoyl-oleoyl-phosphatidylethanolamine (POPE) molecules. Ray-traced images were obtained with PyMOL v1.7.2 RC2,
https://pymol.org/2/, version 1.7.2, accessed on 31 December 2017.
2.2. Cell Culture
African green monkey Vero and HEK-293T cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) (Gibco, Carlsbad, CA, USA) supplemented with 100 U/mL penicillin G sodium/100 μg/mL streptomycin sulfate (Gibco) and 10% (v/v) heat-inactivated fetal bovine serum (FBS) (Gibco).
2.3. Plasmids
The I329L ectodomain with the transmembrane region (ECDTM) (aminoacids 1 to 260), the putative dimerization domain with the transmembrane region (PDDTM) (aminoacids 149 to 260), the intracellular domain (ICD) (aminoacids 261 to 329), were amplified by PCR from the pcDNA3-I329L-HA construct [
7] and cloned into the pcDNA3-HA plasmid. All constructs contained a C-terminal HA “immunotag” sequence to allow confirmation of protein expression.
The human cytomegalovirus (CMV)–vesicular stomatitis virus G envelope protein, the packaging plasmid pCMVR8.9 and the vector pHR-CMV-eGFP constructs have been described previously [
14]. For construction of a recombinant lentivirus vector (pHR-CMV-I329LeGFP), I329L was excised from pCDNA3, together with the HA tag and cloned into the vector pHR-CMV-eGFP upstream of an internal ribosome entry site-driven enhanced green fluorescent protein gene (eGFP).
The luciferase reporter plasmid containing the sequence of the IFN-β promoter [pIFΔ(−125/+72)lucter], [p(PRDII)5tkA(-39)lucter] were gifts of Dr. S. Goodbourn (St George’s Biomedical Sciences Research Centre, University of London, UK). The pCMVβ plasmid contains a β-galactosidase gene that serves as an internal control for transfection efficiency.
The pCMV-1Flag fused to TRIF, hTLR3, hTLR5, hTLR8 were gifts from Dr A. Bowie (Trinity Biomedical Sciences Institute, Trinity College Dublin, Ireland).
The EEA1 Ct-tomato plasmid was a gift from Professor H. Stenmark (Department of Biochemistry, The Norwegian Radium Hospital, Norway).
2.4. Production and Measurement of Lentivirus
Lentivirus was produced by transient transfection of HEK-293T cells with a weight ratio of 3:1:1 of vector to packaging to envelope plasmids using Fugene 6 (Roche, Basel, Switzerland) according to the manufacturer’s instructions. Control lentivirus was produced by co-transfection of the packaging and envelope plasmid together with the empty pHR-CMV-eGFP plasmid. For production of recombinant lentivirus expressing I329L, the plasmid pHR-CMV-I329L- eGFP was used. Supernatants containing the lentivirus were collected at 48, 72 and 96 h post-transfection and clarified by low-speed centrifugation, and the lentivirus was collected by ultracentrifugation (25,000 r.p.m. in an SW28 rotor in a Beckman centrifuge). Virus pellets were resuspended in fresh culture medium and frozen at −80 °C. Lentivirus titres were measured by infection of HEK-293T cells with a dilution factor of 4. Analysis of lentivirus-infected cells was performed by detecting eGFP-positive cells by flow cytometry at 48 h post-infection (p.i.).
2.5. Immunofluorescence
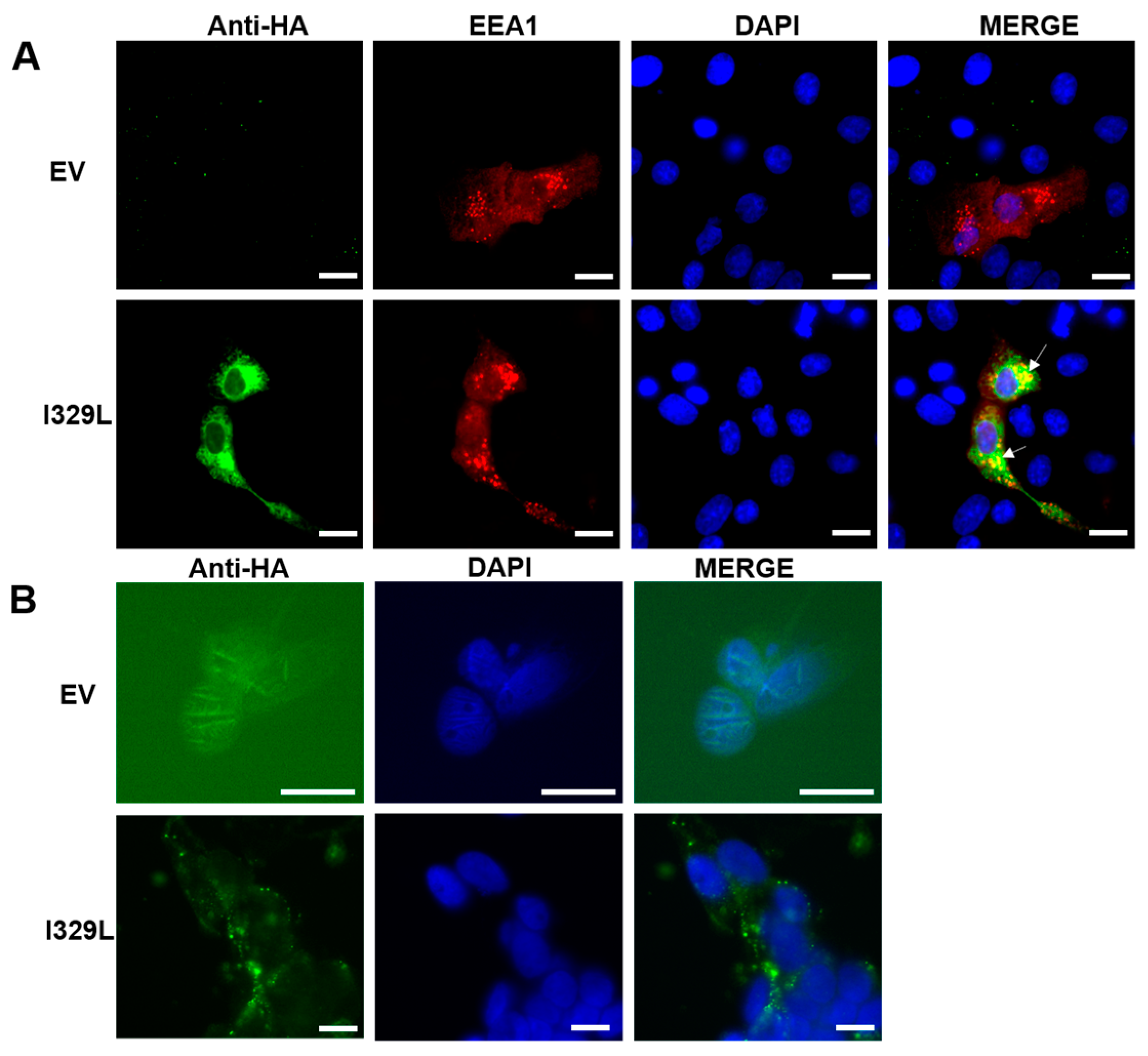
Vero cells grown on coverslips were transfected using X-treme9 (Roche) with pcDNA3-HA empty vector or pcDNA3-I329L-HA and EEA1 Ct-tomato plasmid (to visualize early endosome marker). At 48 h post-transfection, the cells were fixed with 4% paraformaldehyde in PBS for 15 min at room temperature. The cells were permeabilized with PBS containing 0.1% Triton X-100 for 20 min, washed and blocked with PBS containing 5% normal goat serum and 0.05% Tween 20 for 30 min. In an alternative protocol, the fixation and permeabilization step were omitted in order to stain cell surface proteins. The cells were then incubated with a rat monoclonal anti-HA (SIGMA, St. Louis, MO, USA), and then anti-mouse cy5 (Jackson, Chicago, IL, USA) to visualize the HA tagged I329L, and the cell nuclei were stained with DAPI.
2.6. Luciferase Reporter Gene Assay
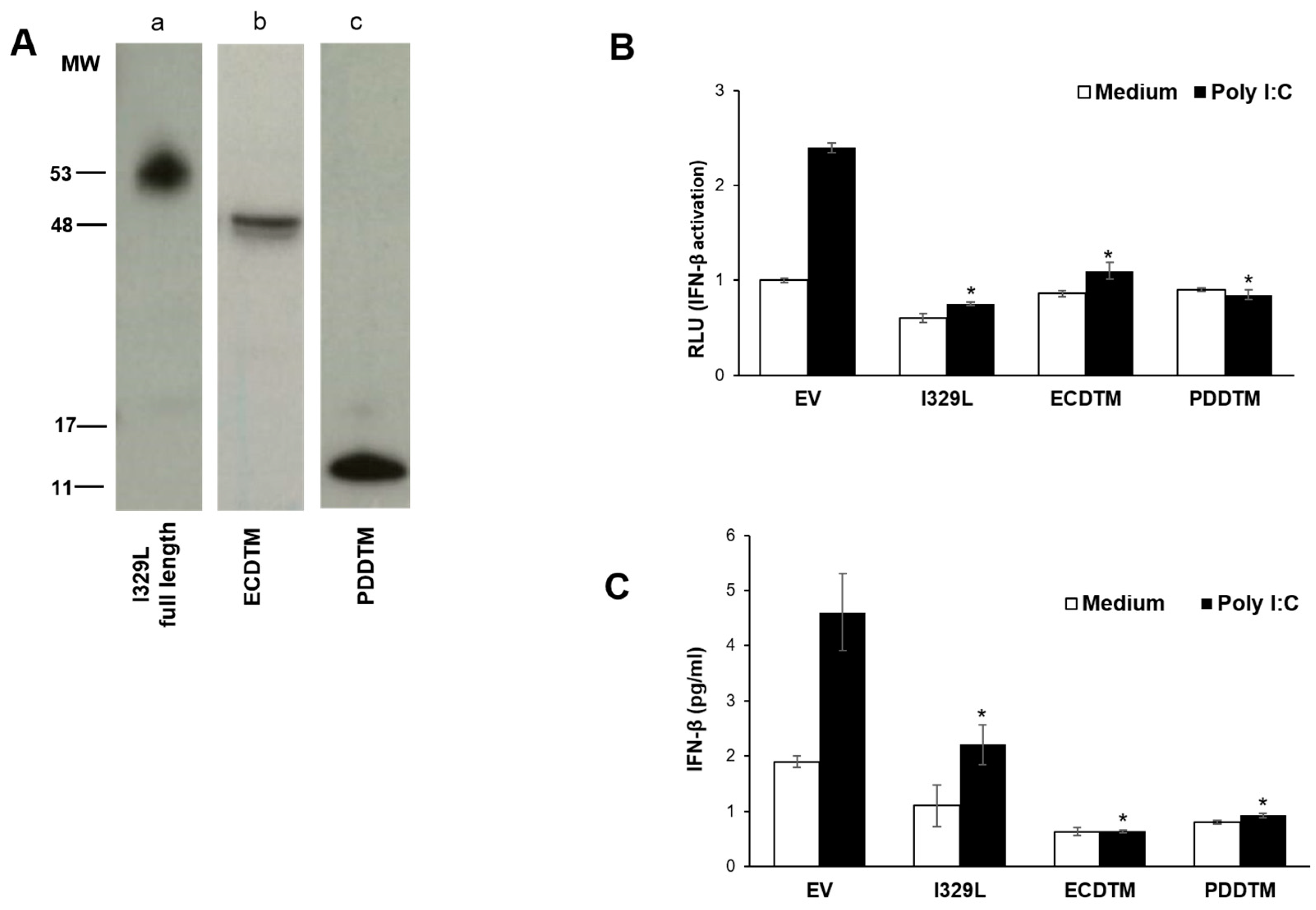
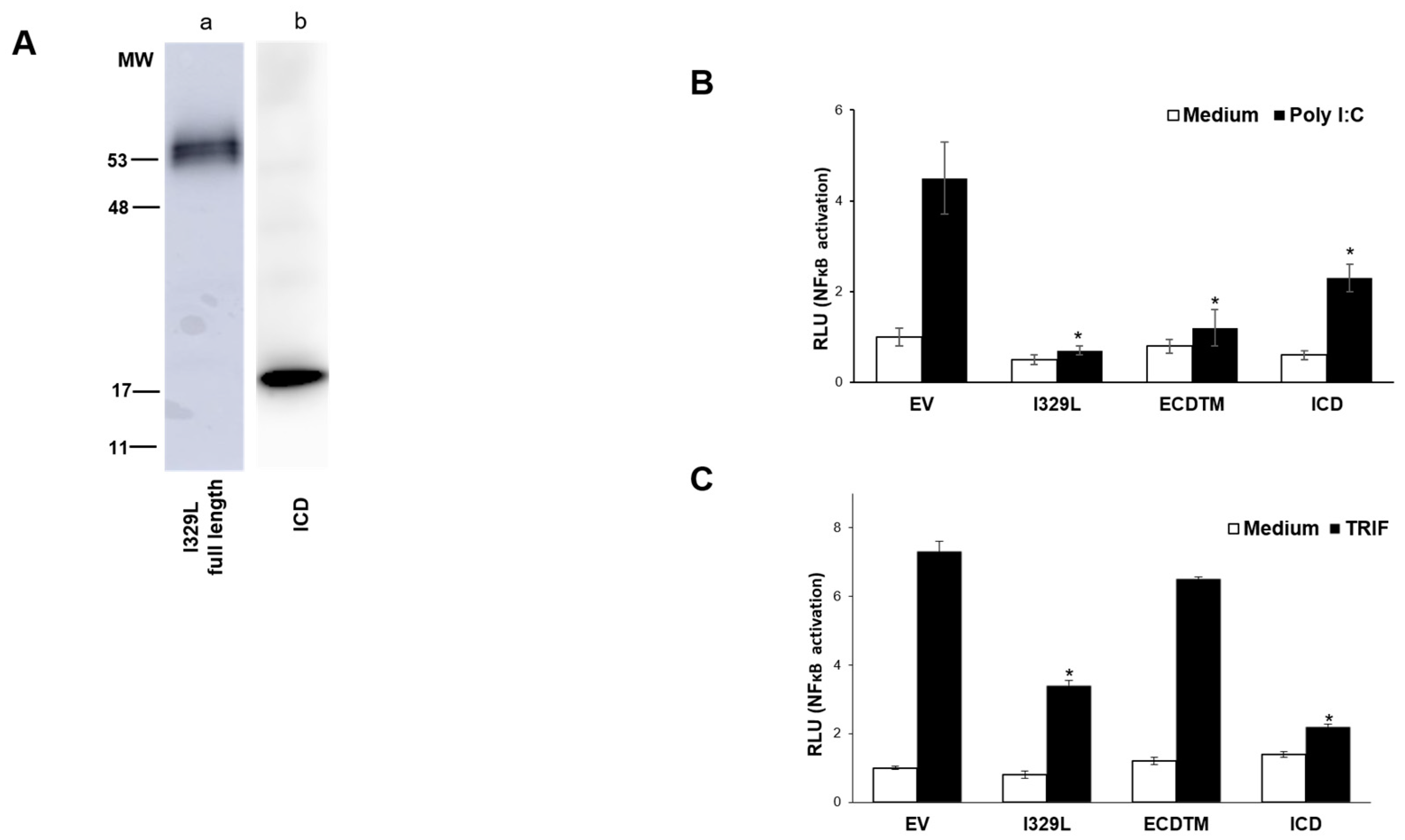
To define the impact of I329L domains in type I IFN induction via TLR3/TRIF, HEK-293T cells were co-transfected with the IFN-β luciferase reporter plasmid, a TLR3 expression plasmid, the β-galactosidase internal control plasmid and either the empty vector pcDNA3HA (EV), pcDNA3-I329L-HA, pcDNA3-ECDTM-HA, pcDNA3-ICD-HA or pcDNA3-PDDTM-HA plasmids, as indicated, according to the Lipofectamine 2000 (Invitrogen) protocol. Cells were either induced with Poly(I:C) (Amersham, Stafford, UK) (25 μg/mL), ectopic TRIF (pCMV-1Flag-TRIF) expression or left untreated (medium).
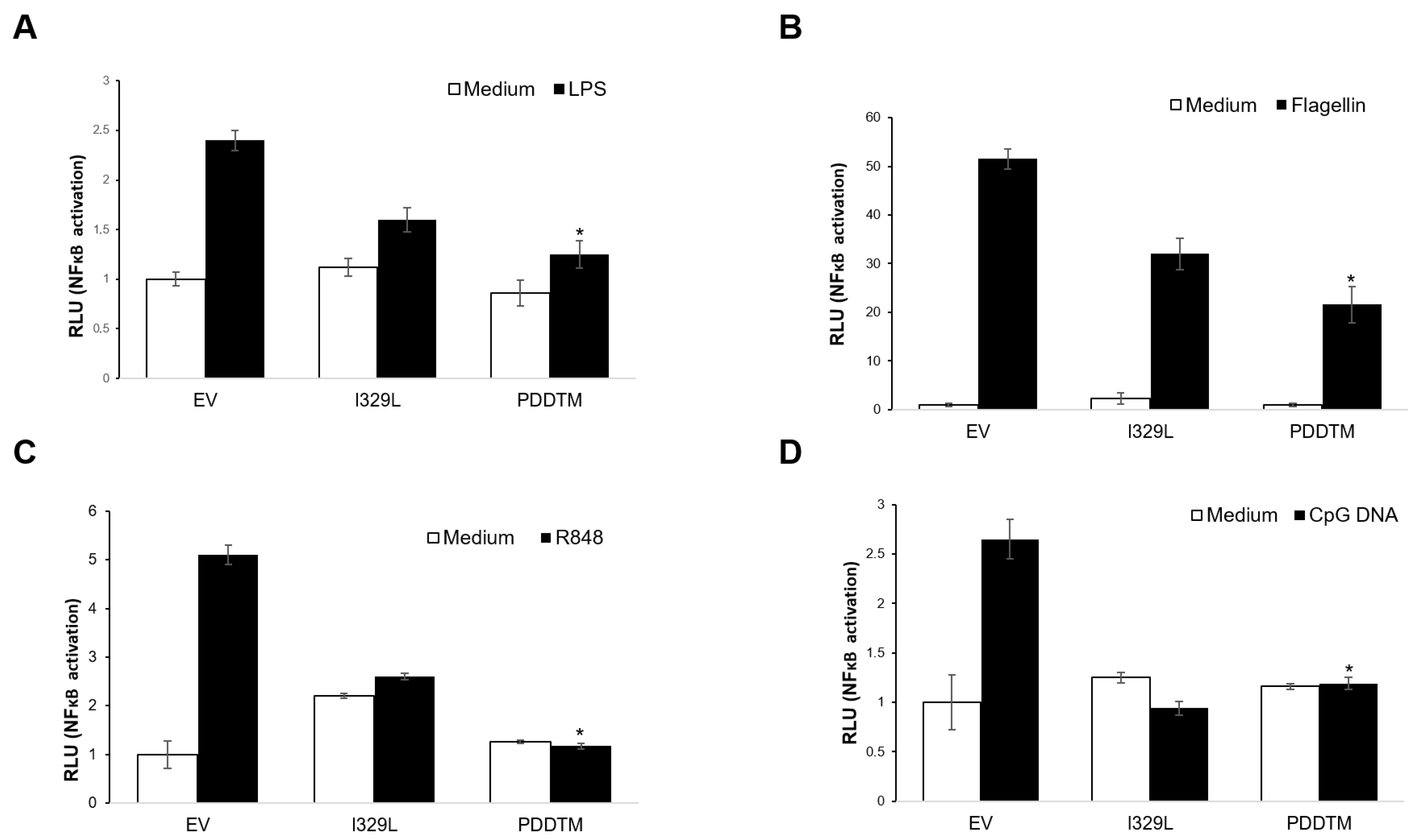
To investigate the impact of I329L and its domains on signalling through TLR5 and TLR8, HEK-293T cells were co-transfected with the NF-κB (PRDII element)-luciferase reporter plasmid, the β-galactosidase internal control plasmid, and either TLR5 or TLR8 plasmids, together with either empty vector pcDNA3HA (EV), pcDNA3-I329L-HA, pcDNA3-ECDTM-HA or pcDNA3-PDDTM-HA plasmids, as indicated, according to the Lipofectamine 2000 (Invitrogen, Waltham, MA, USA) protocol. Cells were either induced with 100 ng/mL Flagellin (Alexis Biochemicals, Lausen, Switzerland) for the TLR5 transfected cells or 1 μg/mL R848 (Invivogen) for the TLR8 transfected cells. Control cells were left untreated.
In an alternative protocol, to test the impact of I329L and its domains on TLR4 and TLR9 signalling pathways, Vero cells were co-transfected with the NF-κB (PRDII element)-luciferase reporter plasmid, the β-galactosidase internal control plasmid, and either the empty vector pcDNA3HA (EV), pcDNA3-I329L-HA, pcDNA3-ECDTM-HA or pcDNA3-PDDTM-HA plasmids, as indicated, according to the Lipofectamine 2000 (Invitrogen) protocol. Cells were either induced with 100 ng/mL LPS (Invivogen) to stimulate TLR4, or 10 μg/mL ODN 2006 (Class B CpG) (Invivogen) to stimulate TLR9. Control cells were left untreated.
Luciferase activity was normalized to β-galactosidase activity as a control for transfection efficiency. Data are expressed as means of relative luciferase units (RLU) ± SD of triplicate culture wells, and are representative of three independent experiments. Statistically significant difference was assessed by Student’s t-test and is represented as p ≤ 0.05 (*).
2.7. Immunoprecipitation to Demonstrate Association of I329L and TLR3
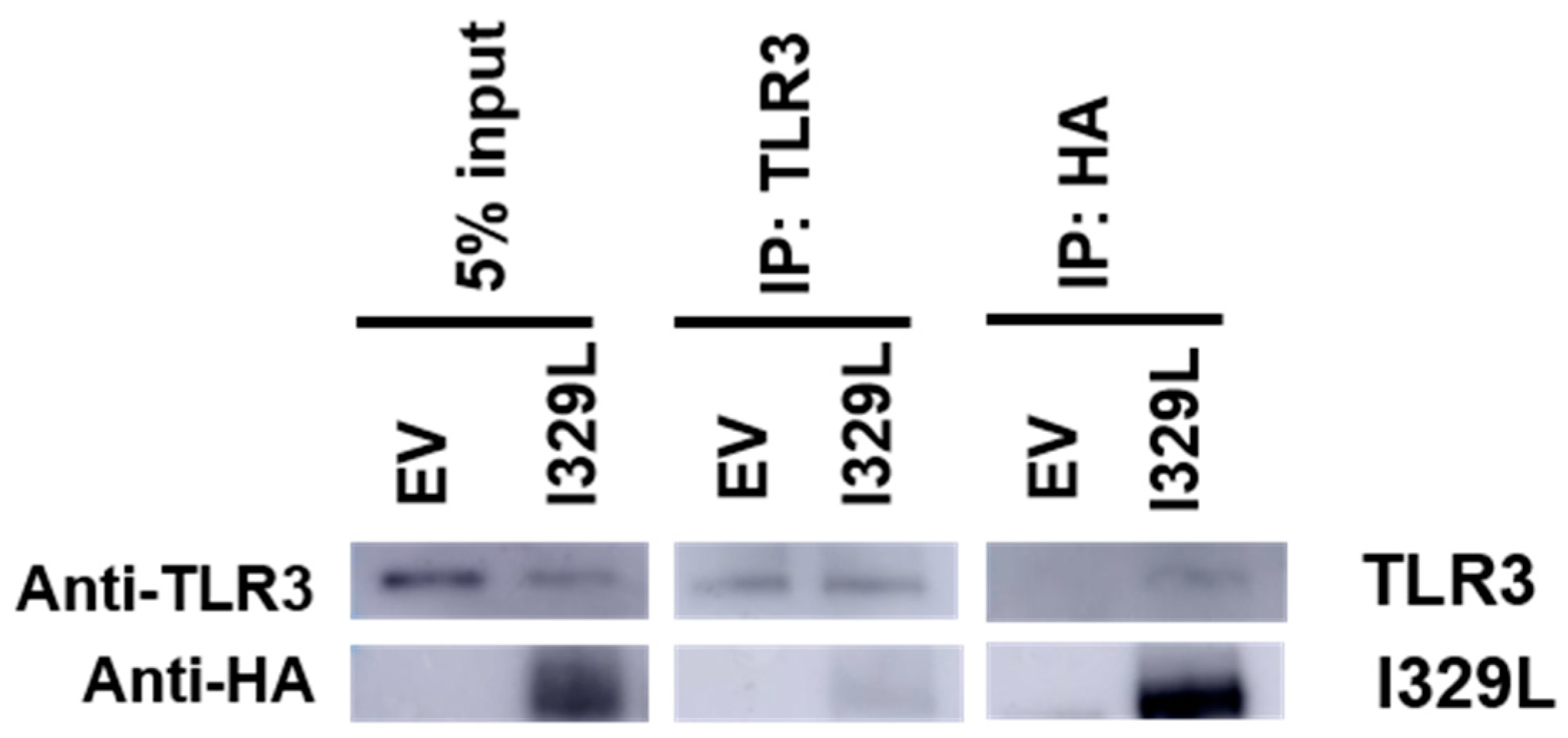
HEK-293T cells were stably infected with HA-I329L recombinant lentivirus (pHR-CMV-I329LeGFP), and as a negative control, with the control “empty” recombinant lentivirus (pHR-CMV-eGFP), and then transfected with TLR3-Flag expression plasmid, according to the Fugene 6 (Promega, Madison, WI, USA) protocol. At 48 h post-transfection, the cells were harvested and lysed in 1% digitonin lysis buffer (1% digitonin, 50 mM Iodoacetamide and 20 mM Tris-HCl (pH 8.0)) containing a protease inhibitor cocktail (SIGMA). Immunoprecipitations were performed with Dynabeads protein G (Millipore, Burlington, MA, USA) and 0.5% digitonin washing buffer (0.5% digitonin and 35 mM Tris-HCl (pH 8.0)) containing a protease inhibitor cocktail (SIGMA), using mouse anti-human TLR3 (Thermofisher, Waltham, MA, USA) and mouse anti-HA (SIGMA). Immunoprecipitates were resolved on a 10% SDS-PAGE gel and Western blot was performed using: mouse anti-human TLR3 (Thermofisher) followed by horseradish peroxidase goat anti-mouse secondary antibodies (Invitrogen), or rat anti-HA-HRP conjugated (Roche) to detect I329L.
2.8. Immunoprecipitation to Demonstrate Association of ICD-I329L and TRIF
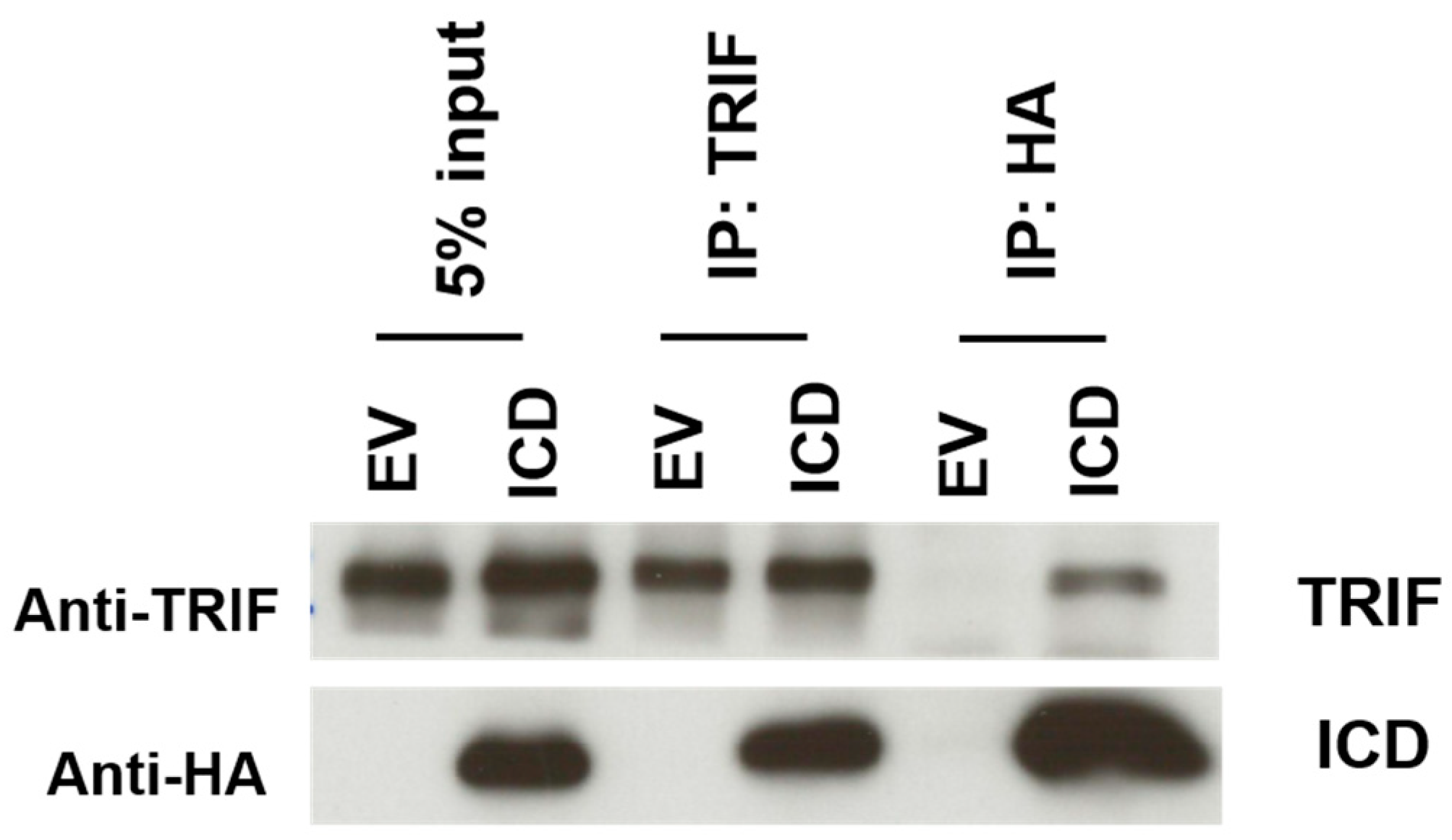
HEK-293T cells were transfected with pcDNA3-HA (EV) or pcDNA3-ICD-HA (ICD), according to the Fugene 6 (Promega) protocol. 48 h post-transfection, the cells were harvested and lysed in 1× lysis buffer (Cell Signaling, Danvers, MA, USA) containing Triton X-100 and a protease inhibitor cocktail (SIGMA). Immunoprecipitations were performed with Dynabeads protein G (Millipore) using rabbit anti-human TRIF antibody (Cell Signaling), and rabbit anti-HA (SIGMA). Immunoprecipitates were resolved on a 12% SDS-PAGE gel and detection on Western blot was performed using: rabbit anti-human TRIF (Cell Signaling) followed by horseradish peroxidase goat anti-rabbit secondary antibodies (Invitrogen), or rat anti-HA-HRP conjugated (Roche) to detect the ICD of I329L.
2.9. ELISA
HEK-293T cells were co-transfected with empty vector pcDNA3HA (EV), pcDNA3-I329L-HA, pcDNA3-ECDTM-HA or pcDNA3-PDDTM-HA and TLR3 plasmid, according to Lipofectamine 2000 (Invitrogen) protocol. At 48 h post-transfection the cells were either stimulated with 25 μg/mL of Poly(I:C) for 5 h, or left untreated. Cell supernatants were collected and ELISA for IFN-β was performed according to the manufacturer’s protocol (Human IFN-β bioluminescent ELISA kit from Invivogen). Data are expressed as means ± SD of triplicate wells and are representative of three independent experiments. Statistically significant difference was assessed by Student’s t-test and is represented as p ≤ 0.05 (*).
4. Discussion
In this work, we define the mechanism of I329L, a virus inhibitor of TLR responses. Remarkably, I329L inhibits TLR responses by two entirely different mechanisms, one through its extracellular domain (ECD) and the other through its intracellular domain (ICD). The ASFV I329L gene codes for a Type I membrane protein which is highly conserved in the pathogenic and non-pathogenic virus, and which consists of a putative signal peptide (aminoacids 1–17), an extracellular N terminal domain (ECD), (aminoacids 18–239), a cytoplasmic domain (ICD) (aminoacids 261–329), and a transmembrane region (aminoacids 240–260). Subsequent work confirmed the inhibition of secreted IFN by I329L [
3], and bioinformatics modelling [
9] suggested that (1) The ECD of I329L might serve as a dsRNA decoy or hinder TLR3 signalling through formation of a I329L-TLR3 heterodimer, and (2) The ICD might inhibit through an impact on the downstream signalling intermediate TRIF.
We now demonstrate that I329L is a general inhibitor of TLR activation, with appropriate cellular localization on the surface membrane and in the early endosome. Furthermore, we present evidence supporting the claim that the ECDTM and ICD regions inhibit activation of IFN-β promoter and NF-κB by entirely different mechanisms, effectively preventing the induction of type I IFN and pro-inflammatory cytokines. Specifically, the ECDTM region inhibits activation of TLR3, TLR4, TLR5, TLR8 and TLR9 at the receptor level, whereas the ICD, but not the ECDTM, interferes with the adaptor TRIF, blocking intracellular activation of TLR3 and TLR4 signalling pathways.
The bioinformatics analysis of I329L identified a short sequence marginally homologous to the TLR dimerization domain, named the putative dimerization domain (PDD). Importantly, we show that this domain not only inhibits the activation of IFN-β promoter via TLR3, but also the activation of NF-κB by TLR4, TLR5, TLR8 and TLR9. On the basis of this broad specificity, we therefore hypothesized that the ECDTM of I329L might generally inhibit TLR activation through forming a non-signalling association with the TLR molecule via interaction of the PDD. The demonstration of a weak TLR3-I329L association by immunoprecipitation (in digitonin but not in Triton X-100) was consistent with this hypothesis.
We also reveal a previously unknown association between the ICD of I329L and TRIF, suggesting a role for I329L as a sequester of TRIF in the cytoplasm, thus preventing its interaction with signalling TLR3 or TLR4 homodimers. This hypothesis is supported by our experimental results and consistent with the previously observed inhibition of both TLR3 and TLR4 signalling by I329L [
3], and the fact that both TLR3 and TLR4 signal via TRIF.
One area left unexplored is the precise function of the I329L DID region. Its structural homology to the ligand-binding region of TLR3 suggests that I329L could function as a competitor for binding to RNA or dsRNA. Further studies are required to explore this possibility, which could extend our understanding of I329L function and perhaps add another mechanism by which this viral protein prevents activation of TLR responses.
From the point of view of vaccine construction, we have further characterized the mechanism of I329L, a novel TLR antagonist encoded by the African Swine Fever Virus (ASFV) [
3,
7,
9], as a prelude to its practical exploitation, for example, for the development of a possible attenuated virus vaccine for ASFV. Significantly, deletion of the I329L gene from an already attenuated ASFV strain, OURT88/3, led to a reduction on its capacity to induce protection against a highly virulent virus challenge [
8]. This excessive attenuation with loss of capacity to induce protection has been described as a consequence of serial passage in cell cultures [
18] or of successive gene deletion [
19,
20]. In addition, the deletion of I329L from a highly virulent strain, Georgia/2007, was not enough for attenuation [
8]. Therefore, I329L is a player in the equilibrium between attenuation and protection and its characterization is certainly a valid contribution for the future development of an effective and safe vaccine against ASF.
The Toll-like receptor (TLR) family plays a crucial regulatory role in both the early innate and the later adaptive immune responses, driving pathogenic mechanisms in various immune-mediated and autoimmune diseases such as infection-associated sepsis, systemic lupus erythematosus, rheumatoid arthritis, and atherosclerosis. Manipulation of TLR responses offers an extraordinary opportunity for clinical intervention in health and disease [
21,
22,
23,
24,
25]. For example, the development of structural analogues of TLR ligands or sequesters of adaptor proteins involved in TLR signalling is a considerable area of pharmaceutical research. Pathogens, such as ASFV, are known to develop strategies to suppress TLR signalling [
26,
27,
28,
29]. Our work clearly presents I329L as a viral immunomodulatory protein with at least two distinct mechanisms of action, and thus its potential as a therapeutic option for the control of excessive TLR activation provides an exciting avenue for future research.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







