
Isolation, Characterization, and Transcriptome Analysis of an ISKNV-Like Virus from Largemouth Bass
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Processing of Sick Fish Samples
2.2. Fish and Cell
2.3. Virus Detection
2.4. Virus Isolation
2.5. Transmission Electron Microscopy Assay
2.6. Complete Genomic Analysis of ISKNV-ZY in Cell Culture
2.7. Experimental Infection
2.8. Transcriptomic Analysis of Virus-Infected Largemouth Bass
2.9. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
2.10. Statistical Analysis
3. Results
3.1. Pathological Signs of Diseased Fish
3.2. Virus Isolation
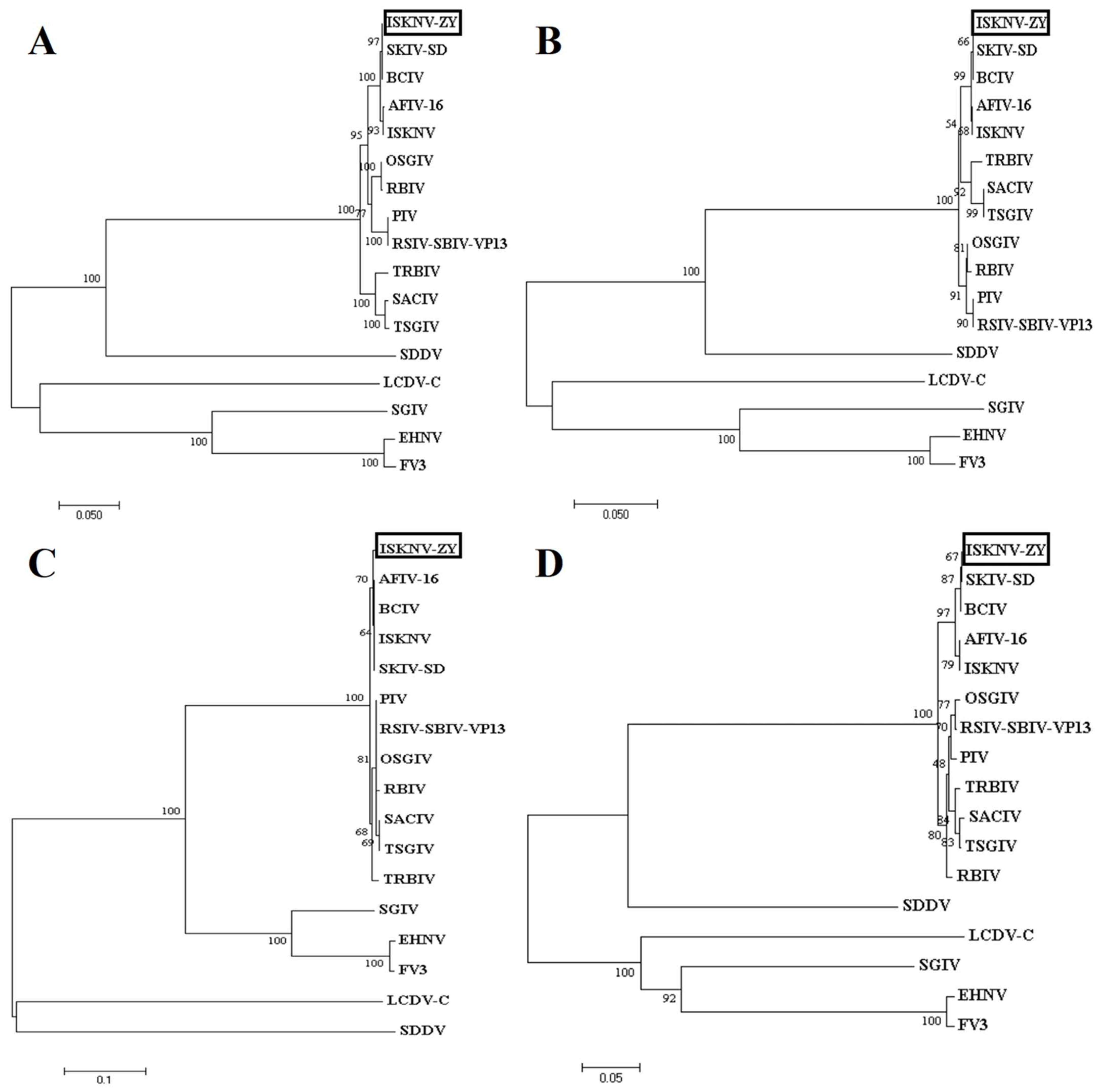
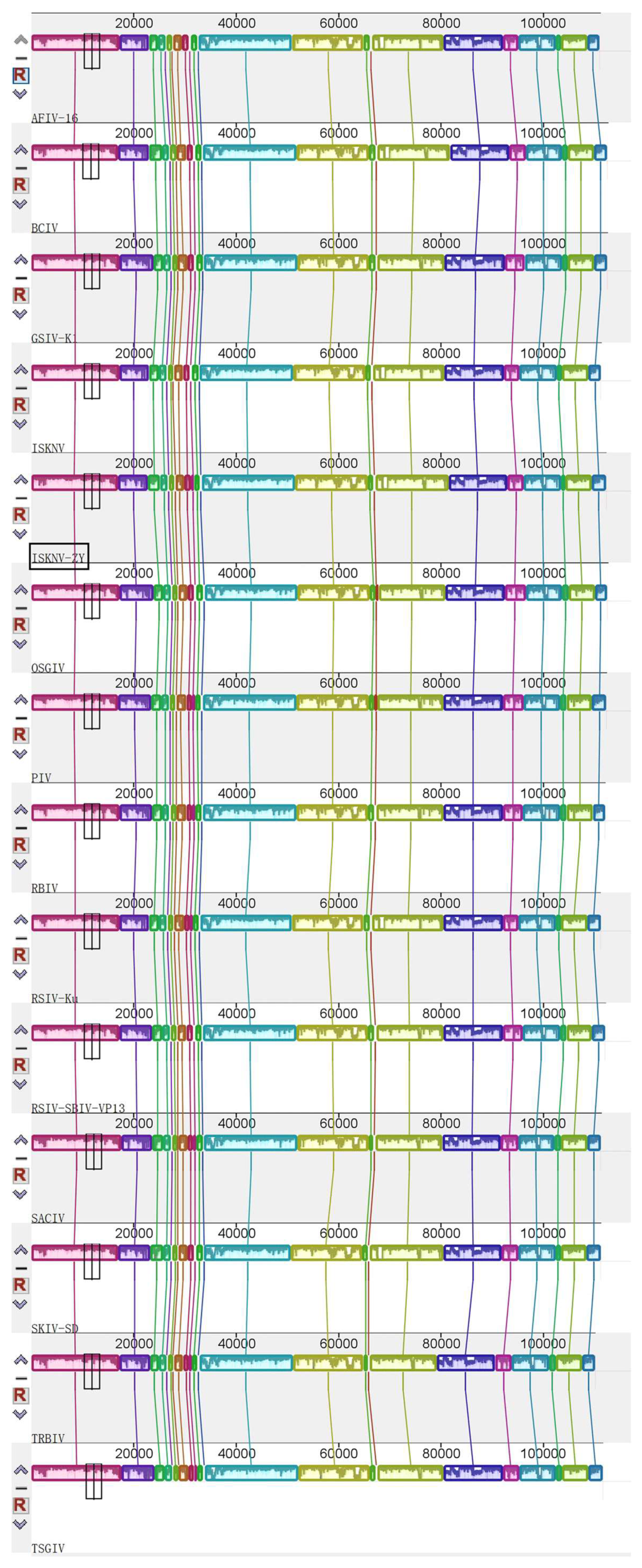
3.3. Genome Sequencing and Functional Annotation
3.4. Experimental Infection of Isolated Viruses in Largemouth Bass
3.5. Transcriptomic Analysis of Virus-Infected Largemouth Bass
3.6. DEG Validation Using qRT-PCR
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Williams, T.; Barbosa-Solomieu, V.; Chinchar, V.G. A decade of advances in iridovirus research. Adv. Virus Res. 2005, 65, 173–248. [Google Scholar]
- Chinchar, V.G.; Hyatt, A.; Miyazaki, T.; Williams, T. Family Iridoviridae: Poor viral relations no longer. Curr. Top. Microbiol. Immunol. 2009, 328, 123–170. [Google Scholar]
- Kurita, J.; Nakajima, K. Megalocytiviruses. Viruses 2012, 4, 521–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuang, F.; Luo, Y.; Xiong, X.P.; Weng, S.; Li, Y.; He, J.; Dong, C. Virions proteins of an RSIV-type megalocytivirus from spotted knifejaw Oplegnathus punctatus (SKIV-ZJ07). Virology 2013, 437, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinchar, V.; Essbauer, S.; He, J.; Hyatt, A.; Miyazaki, T.; Seligy, V.; Williams, T. Part II—The Double Stranded DNA Viruses, Family Iridoviridae. In LA Ball Virus Taxonomy, VIIIth Report of the International Committee on Taxonomy of Viruses; Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Eds.; Elsevier: London, UK; Academic Press: London, UK, 2005. [Google Scholar]
- Nakajima, K.; Maeno, Y.; Honda, A.; Yokoyama, K.; Tooriyama, T.; Manabe, S. Effectiveness of a vaccine against red sea bream iridoviral disease in a field trial test. Dis. Aquat. Org. 1999, 36, 73–75. [Google Scholar] [CrossRef]
- He, J.G.; Deng, M.; Weng, S.P.; Li, Z.; Zhou, S.Y.; Long, Q.X.; Wang, X.Z.; Chan, S.M. Complete genome analysis of the mandarin fish infectious spleen and kidney necrosis iridovirus. Virology 2001, 291, 126–139. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.C.; Zhang, J.; Sun, L. In-depth profiling and analysis of host and viral microRNAs in Japanese flounder (Paralichthys olivaceus) infected with megalocytivirus reveal involvement of microRNAs in host-virus interaction in teleost fish. BMC Genom. 2014, 15, 878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, C.Y.; Jia, K.T.; Yang, B.; Huang, J. Complete genome sequence of a Megalocytivirus (family Iridoviridae) associated with turbot mortality in China. Virol. J. 2010, 7, 159. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.L.; Xu, X.P.; He, B.L.; Weng, S.P.; Xiao, J.; Wang, L.; Lin, T.; Liu, X.; Wang, Q.; Yu, X.Q.; et al. Infectious spleen and kidney necrosis virus ORF48R functions as a new viral vascular endothelial growth factor. J. Virol. 2008, 82, 4371–4383. [Google Scholar] [CrossRef] [Green Version]
- Go, J.; Lancaster, M.; Deece, K.; Dhungyel, O.; Whittington, R. The molecular epidemiology of iridovirus in Murray cod (Maccullochella peelii peelii) and dwarf gourami (Colisa lalia) from distant biogeographical regions suggests a link between trade in ornamental fish and emerging iridoviral diseases. Mol. Cell. Probes 2006, 20, 212–222. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Lu, L.; Weng, S.P.; Huang, J.N.; Chan, S.M.; He, J.G. Molecular epidemiology and phylogenetic analysis of a marine fish infectious spleen and kidney necrosis virus-like (ISKNV-like) virus. Arch. Virol. 2007, 152, 763–773. [Google Scholar] [CrossRef]
- Jeong, J.B.; Kim, H.Y.; Jun, L.J.; Lyu, J.H.; Park, N.G.; Kim, J.K.; Jeong, H.D. Outbreaks and risks of infectious spleen and kidney necrosis virus disease in freshwater ornamental fishes. Dis. Aquat. Org. 2008, 78, 209–215. [Google Scholar] [CrossRef]
- Jin, Y.; Bergmann, S.M.; Mai, Q.; Yang, Y.; Liu, W.; Sun, D.; Chen, Y.; Yu, Y.; Liu, Y.; Cai, W.; et al. Simultaneous Isolation and Identification of Largemouth Bass Virus and Rhabdovirus from Moribund Largemouth Bass (Micropterus salmoides). Viruses 2022, 14, 1643. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Li, M.; Liu, M.; Huang, S.; Wang, G.; Wang, T.; Li, P. Selection and Characterization of ssDNA Aptamers Targeting Largemouth Bass Virus Infected Cells with Antiviral Activities. Front. Microbiol. 2021, 12, 785318. [Google Scholar] [CrossRef]
- Jia, Y.J.; Guo, Z.R.; Ma, R.; Qiu, D.K.; Wang, G.X.; Zhu, B. Protective immunity of largemouth bass immunized with immersed DNA vaccine against largemouth bass ulcerative syndrome virus. Fish Shellfish Immunol. 2020, 107 Pt A, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Chu, X.; Han, M.; Yu, C.; Jiang, Y.; Wang, H.; Lu, L.; Xu, D. Rapid visual detection of Micropterus salmoides rhabdovirus using recombinase polymerase amplification combined with lateral flow dipsticks. J. Fish Dis. 2022, 45, 461–469. [Google Scholar] [CrossRef]
- Chao, C.B.; Chen, C.Y.; Lai, Y.Y.; Lin, C.S.; Huang, H.T. Histological, ultrastructural, and in situ hybridization study on enlarged cells in grouper Epinephelus hybrids infected by grouper iridovirus in Taiwan (TGIV). Dis. Aquat. Org. 2004, 58, 127–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, M.H.; Nikapitiya, C.; Vinay, T.N.; Lee, J.; Jung, S.J. Rock bream iridovirus (RBIV) replication in rock bream (Oplegnathus fasciatus) exposed for different time periods to susceptible water temperatures. Fish Shellfish Immunol. 2017, 70, 731–735. [Google Scholar] [CrossRef]
- Rimmer, A.E.; Becker, J.A.; Tweedie, A.; Lintermans, M.; Landos, M.; Stephens, F.; Whittington, R.J. Detection of dwarf gourami iridovirus (Infectious spleen and kidney necrosis virus) in populations of ornamental fish prior to and after importation into Australia, with the first evidence of infection in domestically farmed Platy (Xiphophorus maculatus). Prev. Vet. Med. 2015, 122, 181–194. [Google Scholar] [CrossRef]
- Chiew, I.K.M.; Salter, A.M.; Lim, Y.S. The significance of major viral and bacterial diseases in Malaysian aquaculture industry. Pertanika J. Trop. Agric. Sci. 2019, 42, 1023–1047. [Google Scholar]
- Nakajima, K.; Kunita, J. Red sea bream iridoviral disease. Uirusu 2005, 55, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Kurita, J.; Nakajima, K.; Hirono, I.; Aoki, T. Complete genome sequencing of red sea bream iridovirus (RSIV). Fish. Sci. 2002, 68 (Suppl. S2), 1113–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, J.W.; Moon, C.H.; Kim, H.J.; Ko, M.S.; Kim, S.B.; Son, J.H.; Kim, J.S.; An, E.J.; Kim, M.K.; Lee, S.K.; et al. Complete genomic DNA sequence of rock bream iridovirus. Virology 2004, 325, 351–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Zhou, S.Y.; Chen, C.; Weng, S.P.; Chan, S.M.; He, J.G. Complete genome sequence analysis of an iridovirus isolated from the orange-spotted grouper, Epinephelus coioides. Virology 2005, 339, 81–100. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.B.; Jun, L.J.; Yoo, M.H.; Kim, M.S.; Komisar, J.L.; Do Jeong, H. Characterization of the DNA nucleotide sequences in the genome of red sea bream iridoviruses isolated in Korea. Aquaculture 2003, 220, 119–133. [Google Scholar] [CrossRef]
- Do, J.W.; Cha, S.J.; Kim, J.S.; An, E.J.; Park, M.S.; Kim, J.W.; Kim, Y.C.; Park, M.A.; Park, J.W. Sequence variation in the gene encoding the major capsid protein of Korean fish iridoviruses. Arch. Virol. 2005, 150, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Weng, S.; Luo, Y.; Huang, M.; Ai, H.; Yin, Z.; He, J. A new marine megalocytivirus from spotted knifejaw, Oplegnathus punctatus, and its pathogenicity to freshwater mandarinfish, Siniperca chuatsi. Virus Res. 2010, 147, 98–106. [Google Scholar] [CrossRef]
- Shinmoto, H.; Taniguchi, K.; Ikawa, T.; Kawai, K.; Oshima, S. Phenotypic diversity of infectious red sea bream iridovirus isolates from cultured fish in Japan. Appl. Environ. Microbiol. 2009, 75, 3535–3541. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Li, N.; Liu, L.; Lin, Q.; Wang, F.; Lai, Y.; Jiang, H.; Pan, H.; Shi, C.; Wu, S. Genotype and host range analysis of infectious spleen and kidney necrosis virus (ISKNV). Virus Genes 2011, 42, 97–109. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Zhou, Y.; Fu, C.-H.; Huang, Z.-H.; Li, Z.; Jin, M.-R.; Fu, Z.-X.-; Li, Q.-N. Transcriptomic and proteomic analyses of the immune mechanism in pathogenetic and resistant mandarin fish (Siniperca chuatsi) infected with ISKNV. Aquaculture 2021, 545, 737198. [Google Scholar] [CrossRef]
- Byadgi, O.; Chen, W.-C.; Wang, C.-P.; Tsai, A.-M.; Chen, C.-S. De novo transcriptome analysis of differential functional gene expression in largemouth bass (Micropterus salmoides) after challenge with Nocardia seriolae. Int. J. Mol. Sci. 2016, 17, 1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geijtenbeek, T.B.; Gringhuis, S.I. Signalling through C-type lectin receptors: Shaping immune responses. Nat. Rev. Immunol. 2009, 9, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′–3′) | GenBank Accession Number |
|---|---|---|
| β-actin-RT-F | CCACCACAGCCGAGAGGGAA | MH018565.1 |
| β-actin-RT-R | TCATGGTGGATGGGGCCAGG | |
| TRAF2B-RT-F | AACAACAGGGAGCACATT | XM_038734170.1 |
| TRAF2B-RT-R | GCCAGCCAGTTTAGCC | |
| STAT1A-RT-F | AACTGCTCGGAGCCAAAG | XM_038715521.1 |
| STAT1A-RT-R | CAGCCAGAAGGGAAACG | |
| STAT2-RT-F | GGCCGACCTAAATAAGC | XM_038696682.1 |
| STAT2-RT-R | AAGAAACTCCCGCACC | |
| MYD88-RT-F | TGCCTTCATCTGCTACTGC | XM_038728827.1 |
| MYD88-RT-R | CACCATCCGCTTACACCT | |
| TRAF6-RT-F | AATAAGGGCAGTTTGGAG | XM_038700569.1 |
| TRAF6-RT-R | GAAGGGCTGAGTAGGGA | |
| DHX58-RT-F | GGGCTGGCAATGGTA | XM_038704577.1 |
| DHX58-RT-R | GCTGCTGGGCAATCTC | |
| SOCS3A-RT-F | CCTCACCGTTACAAGACC | XM_038701469.1 |
| SOCS3A-RT-R | CCTCGGAAGCCAGCAT | |
| SOCS1A-RT-F | ATGGTAGCCGAAAGCACA | XM_038696259.1 |
| SOCS1A-RT-R | AGGGACGGACACCAATCT | |
| CXCL10-RT-F | GCTATTGCCTGAACCC | XM_038699228.1 |
| CXCL10-RT-R | TGCTAGTGCTGCGTGT | |
| IRF3-RT-F | CTGCGAGCACAGATTGAC | XM_038735464.1 |
| IRF3-RT-R | GCCCACGCCTTAAAGAT | |
| IRF7-RT-F | TGTAGCACCCGAGAACC | XM_038706685.1 |
| IRF7-RT-R | GGAATGTGCCCTTTAGC | |
| ATG9B-RT-F | AAAAGGTGCAACACGATG | XM_038717820.1 |
| ATG9B-RT-R | GTCCTGACGCAGTTAGAAT | |
| PIAS1B-RT-F | CCCCATCAACATAACCTC | XM_038706189.1 |
| PIAS1B-RT-R | AATTCCTTTAGCCCGTAG | |
| IRF9-RT-F | GAAGCTGACGGATGGGAGT | XM_038732190.1 |
| IRF9-RT-R | CGAGGCGGTAGACCTTGTAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Z.; Liao, J.; Zhang, D.; Liu, S.; Zhang, L.; Kang, S.; Xu, L.; Chen, H.; Peng, W.; Zhou, S.; et al. Isolation, Characterization, and Transcriptome Analysis of an ISKNV-Like Virus from Largemouth Bass. Viruses 2023, 15, 398. https://doi.org/10.3390/v15020398
Xu Z, Liao J, Zhang D, Liu S, Zhang L, Kang S, Xu L, Chen H, Peng W, Zhou S, et al. Isolation, Characterization, and Transcriptome Analysis of an ISKNV-Like Virus from Largemouth Bass. Viruses. 2023; 15(2):398. https://doi.org/10.3390/v15020398
Chicago/Turabian StyleXu, Zhuqing, Jiaming Liao, Dongzhuo Zhang, Shaoli Liu, Luhao Zhang, Shaozhu Kang, Linting Xu, Hong Chen, Wenquan Peng, Sheng Zhou, and et al. 2023. "Isolation, Characterization, and Transcriptome Analysis of an ISKNV-Like Virus from Largemouth Bass" Viruses 15, no. 2: 398. https://doi.org/10.3390/v15020398