
Antiviral Effect of Manganese against Foot-and-Mouth Disease Virus Both in PK15 Cells and Mice


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Cell Culture and Maintenance
2.3. Virus
2.4. Mn2+ Cytotoxicity Assay in PK15 Cells
2.5. Western Blotting
2.6. RNA Extraction and RT-qPCR
2.7. Animal Infection and Preparation of Samples
2.8. Detection of Cytokine Levels in the Serum
2.9. Statistical Analysis
3. Results
3.1. Antiviral Activity of Mn2+ against FMDV Infection in PK15 Cells
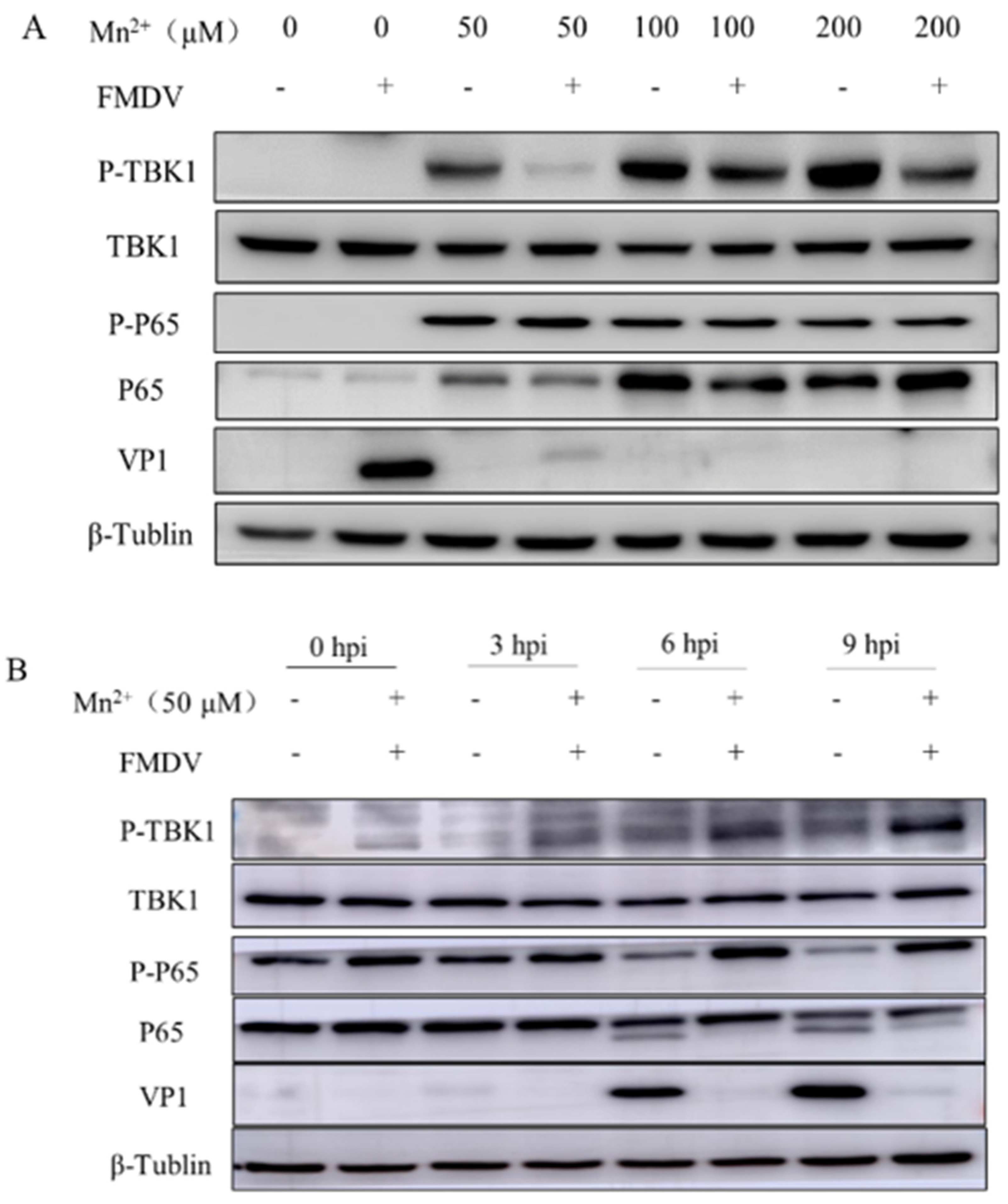
3.2. Mn2+ Activates Anti-Viral Innate Immunity via the NF-κB Signaling Pathway in PK15 Cells
3.3. Enhanced Survival Rate of Mice Inoculated with Mn2+ after Being Challenged by FMDV
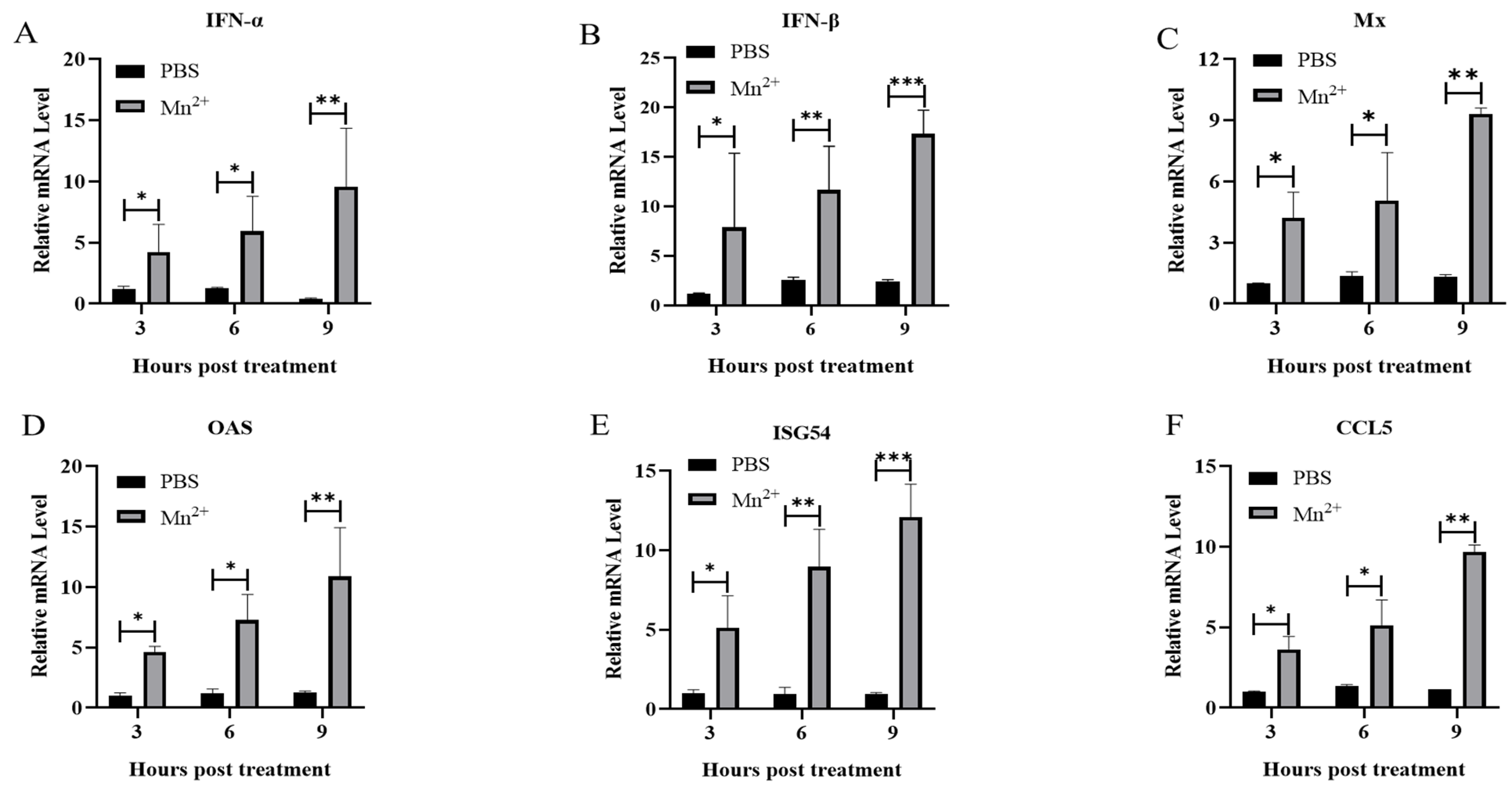
3.4. Cytokine Production in Mice Treated with Mn2+
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mahy, B.W. Introduction and history of foot-and-mouth disease virus. Curr. Top. Microbiol. Immunol. 2005, 288, 1–8. [Google Scholar]
- Rodriguez-Habibe, I.; Celis-Giraldo, C.; Patarroyo, M.E.; Avendano, C.; Patarroyo, M.A. A Comprehensive Review of the Immunological Response against Foot-and-Mouth Disease Virus Infection and Its Evasion Mechanisms. Vaccines 2020, 8, 764. [Google Scholar] [CrossRef]
- Jamal, S.M.; Belsham, G.J. Foot-and-mouth disease: Past, present and future. Vet. Res. 2013, 44, 116. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.; Nelson, N.; Gubbins, S.; Colenutt, C. Airborne Transmission of Foot-and-Mouth Disease Virus: A Review of Past and Present Perspectives. Viruses 2022, 14, 116. [Google Scholar] [CrossRef]
- Knight-Jones, T.J.; Rushton, J. The economic impacts of foot and mouth disease—What are they, how big are they and where do they occur? Prev. Vet. Med. 2013, 112, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, L.L.; Gay, C.G. Development of vaccines toward the global control and eradication of foot-and-mouth disease. Expert Rev. Vaccines 2011, 10, 377–387. [Google Scholar] [CrossRef]
- Mahapatra, M.; Parida, S. Foot and mouth disease vaccine strain selection: Current approaches and future perspectives. Expert Rev. Vaccines 2018, 17, 577–591. [Google Scholar] [CrossRef]
- Salt, J.S.; Barnett, P.V.; Dani, P.; Williams, L. Emergency vaccination of pigs against foot-and-mouth disease: Protection against disease and reduction in contact transmission. Vaccine 1998, 16, 746–754. [Google Scholar] [CrossRef]
- Golde, W.T.; Pacheco, J.M.; Duque, H.; Doel, T.; Penfold, B.; Ferman, G.S.; Gregg, D.R.; Rodriguez, L.L. Vaccination against foot-and-mouth disease virus confers complete clinical protection in 7 days and partial protection in 4 days: Use in emergency outbreak response. Vaccine 2005, 23, 5775–5782. [Google Scholar] [CrossRef]
- Horning, K.J.; Caito, S.W.; Tipps, K.G.; Bowman, A.B.; Aschner, M. Manganese Is Essential for Neuronal Health. Annu. Rev. Nutr. 2015, 35, 71–108. [Google Scholar] [CrossRef]
- Kwakye, G.F.; Paoliello, M.M.; Mukhopadhyay, S.; Bowman, A.B.; Aschner, M. Manganese-Induced Parkinsonism and Parkinson’s Disease: Shared and Distinguishable Features. Int. J. Environ. Res. Public Health 2015, 12, 7519–7540. [Google Scholar] [CrossRef] [PubMed]
- Waldron, K.J.; Rutherford, J.C.; Ford, D.; Robinson, N.J. Metalloproteins and metal sensing. Nature 2009, 460, 823–830. [Google Scholar] [CrossRef]
- Wang, C.; Guan, Y.; Lv, M.; Zhang, R.; Guo, Z.; Wei, X.; Du, X.; Yang, J.; Li, T.; Wan, Y.; et al. Manganese Increases the Sensitivity of the cGAS-STING Pathway for Double-Stranded DNA and Is Required for the Host Defense against DNA Viruses. Immunity 2018, 48, 675–687.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, H.L.; Jhingran, A.; Sun, Y.; Vareechon, C.; de Jesus Carrion, S.; Skaar, E.P.; Chazin, W.J.; Calera, J.A.; Hohl, T.M.; Pearlman, E. Zinc and Manganese Chelation by Neutrophil S100A8/A9 (Calprotectin) Limits Extracellular Aspergillus fumigatus Hyphal Growth and Corneal Infection. J. Immunol. 2016, 196, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Stacy, A.; Abraham, N.; Jorth, P.; Whiteley, M. Microbial Community Composition Impacts Pathogen Iron Availability during Polymicrobial Infection. PLoS Pathog. 2016, 12, e1006084. [Google Scholar] [CrossRef] [Green Version]
- Aschner, J.L.; Aschner, M. Nutritional aspects of manganese homeostasis. Mol. Aspects Med. 2005, 26, 353–362. [Google Scholar] [CrossRef]
- Bowman, A.B.; Aschner, M. Considerations on manganese (Mn) treatments for in vitro studies. Neurotoxicology 2014, 41, 141–142. [Google Scholar] [CrossRef] [Green Version]
- Arzt, J.; Fish, I.H.; Bertram, M.R.; Smoliga, G.R.; Hartwig, E.J.; Pauszek, S.J.; Holinka-Patterson, L.; Diaz-San Segundo, F.C.; Sitt, T.; Rieder, E.; et al. Simultaneous and Staggered Foot-and-Mouth Disease Virus Coinfection of Cattle. J. Virol. 2021, 95, e0165021. [Google Scholar] [CrossRef]
- Paton, D.J.; Reeve, R.; Capozzo, A.V.; Ludi, A. Estimating the protection afforded by foot-and-mouth disease vaccines in the laboratory. Vaccine 2019, 37, 5515–5524. [Google Scholar] [CrossRef]
- Stenfeldt, C.; Arzt, J. The Carrier Conundrum; A Review of Recent Advances and Persistent Gaps Regarding the Carrier State of Foot-and-Mouth Disease Virus. Pathogens 2020, 9, 167. [Google Scholar] [CrossRef] [PubMed]
- Bekerman, E.; Einav, S. Infectious disease. Combating emerging viral threats. Science 2015, 348, 282–283. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Zhang, R.; Yin, J.; Zhao, S.; Qin, X.; Chen, F.; Yang, Y.; Bai, L.; Guo, Z.; Wu, Y.; et al. Antiviral Effect of Manganese against Foot-and-Mouth Disease Virus Both in PK15 Cells and Mice. Viruses 2023, 15, 390. https://doi.org/10.3390/v15020390
Zhang Z, Zhang R, Yin J, Zhao S, Qin X, Chen F, Yang Y, Bai L, Guo Z, Wu Y, et al. Antiviral Effect of Manganese against Foot-and-Mouth Disease Virus Both in PK15 Cells and Mice. Viruses. 2023; 15(2):390. https://doi.org/10.3390/v15020390
Chicago/Turabian StyleZhang, Zhixiong, Rui Zhang, Juanbin Yin, Shuaiyang Zhao, Xiaodong Qin, Fei Chen, Yang Yang, Ling Bai, Zijing Guo, Yongshu Wu, and et al. 2023. "Antiviral Effect of Manganese against Foot-and-Mouth Disease Virus Both in PK15 Cells and Mice" Viruses 15, no. 2: 390. https://doi.org/10.3390/v15020390