
Pre-Clinical Evaluation of the Antiviral Activity of Epigalocatechin-3-Gallate, a Component of Green Tea, against Influenza A(H1N1)pdm Viruses
 , and
, and 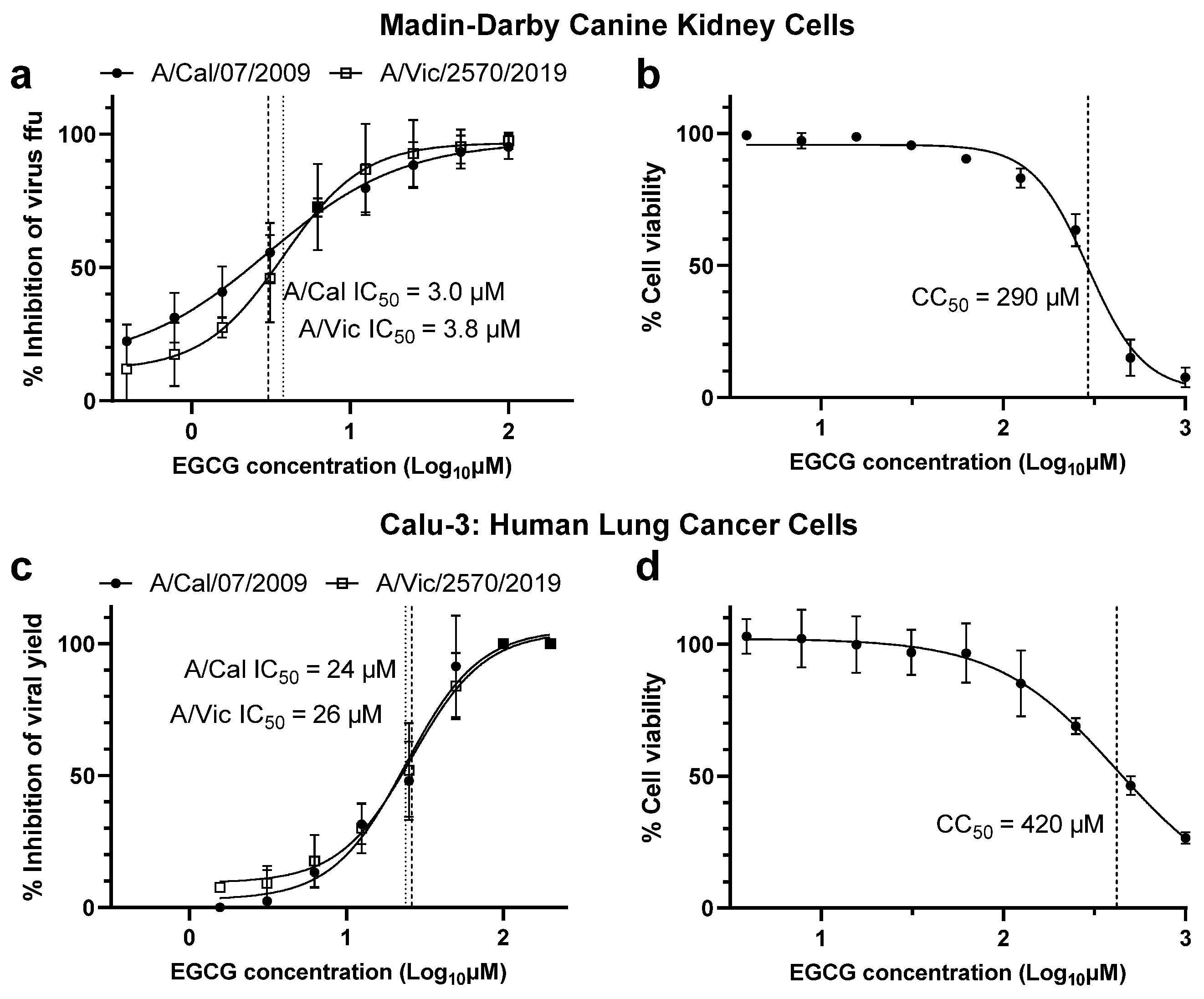{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Compounds
2.3. EGCG Cytotoxicity Assays in MDCK and Calu-3 Cells
2.4. Viruses
2.5. MDCK Cell-Based EGCG Viral Foci Reduction Assay
2.6. Calu-3 Cell-Based EGCG Virus Yield Reduction Assay
2.7. Virus Titration Assay
2.8. Ferrets
2.9. Ferret Toxicity Study
2.10. Ferret Antiviral Treatment Study
2.11. Pyrosequencing
2.12. Statistics and Reproducibility
3. Results
3.1. EGCG Demonstrated Antiviral Efficacy in MDCK and Calu-3 Cells
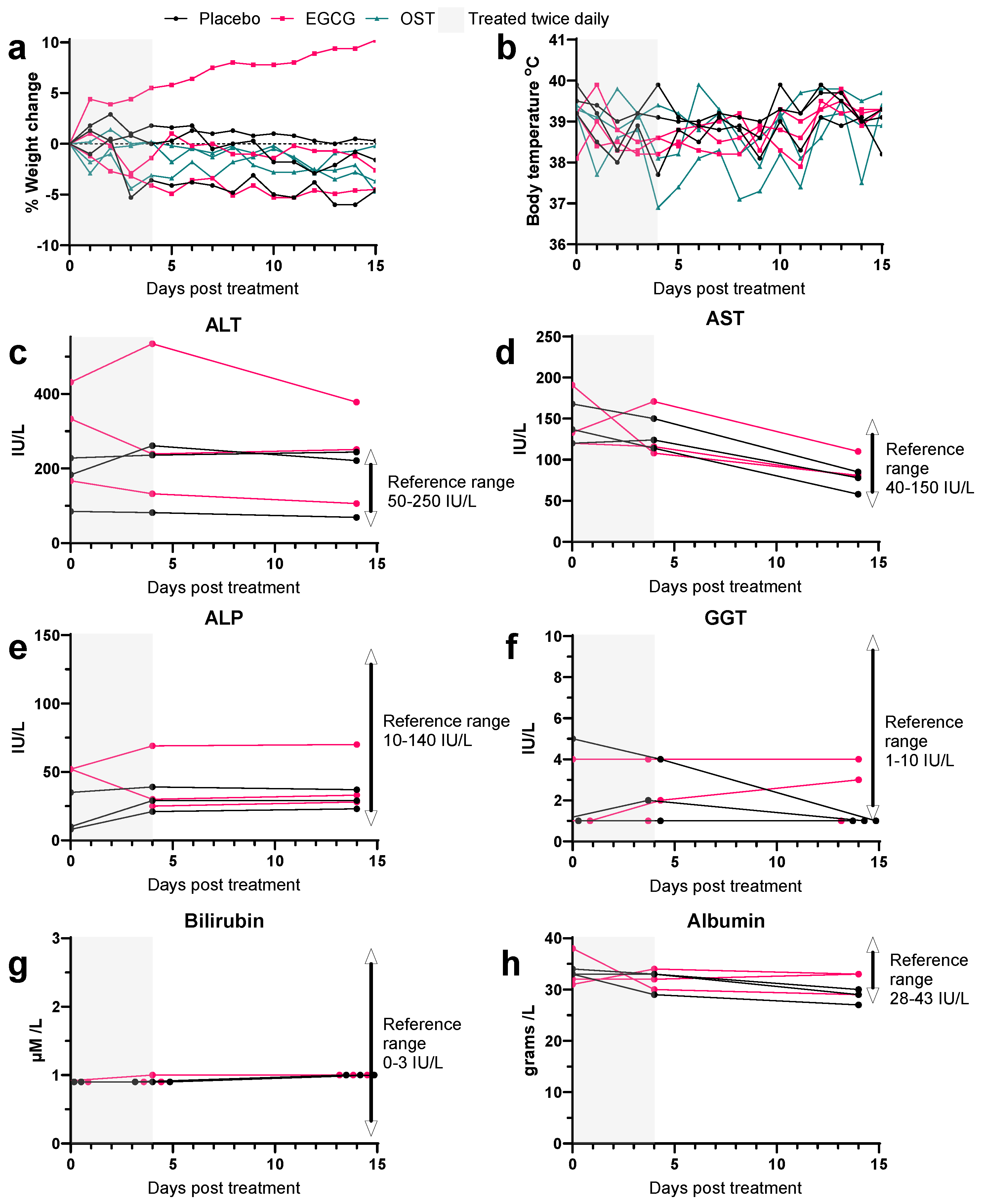
3.2. EGCG Treatment in Uninfected Ferrets Reveals No Signs of Toxicity
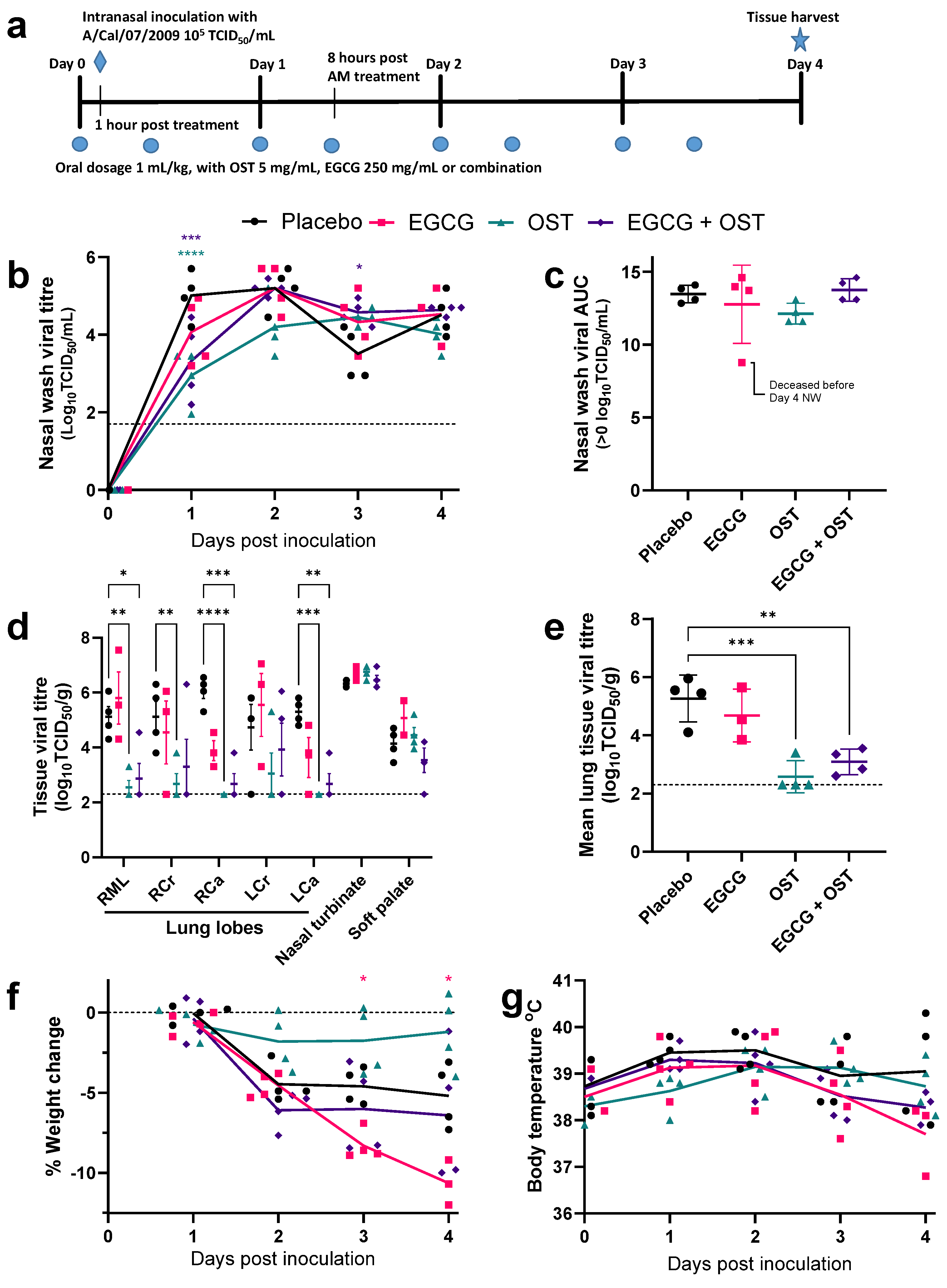
3.3. EGCG Shows No Reduction in Viral Titres of Influenza Infected Ferrets Compared to OST Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef] [PubMed]
- WHO. Up to 650,000 People Die of Respiratory Diseases Linked to Seasonal Flu Each Year. Available online: https://www.who.int/news/item/13-12-2017-up-to-650-000-people-die-of-respiratory-diseases-linked-to-seasonal-flu-each-year (accessed on 25 January 2023).
- Koszalka, P.; George, A.; Dhanasekaran, V.; Hurt, A.C.; Subbarao, K. Effect of Baloxavir and Oseltamivir in Combination on Infection with Influenza Viruses with PA/I38T or PA/E23K Substitutions in the Ferret Model. Mbio 2022, 13, e01056-22. [Google Scholar] [CrossRef] [PubMed]
- Sarker, A.; Gu, Z.; Mao, L.; Ge, Y.; Hou, D.; Fang, J.; Wei, Z.; Wang, Z. Influenza-existing drugs and treatment prospects. Eur. J. Med. Chem. 2022, 232, 114189. [Google Scholar] [CrossRef] [PubMed]
- Arts, I.C.; Hollman, P.C. Polyphenols and disease risk in epidemiologic studies. Am. J. Clin. Nutr. 2005, 81, 317S–325S. [Google Scholar] [CrossRef] [PubMed]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights Into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 283. [Google Scholar] [CrossRef] [PubMed]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [PubMed]
- Trisha, A.T.; Shakil, M.H.; Talukdar, S.; Rovina, K.; Huda, N.; Zzaman, W. Tea Polyphenols and Their Preventive Measures against Cancer: Current Trends and Directions. Foods 2022, 11, 3349. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Wang, H. Cancer Preventive Activities of Tea Catechins. Molecules 2016, 21, 1679. [Google Scholar] [CrossRef]
- Musial, C.; Kuban-Jankowska, A.; Gorska-Ponikowska, M. Beneficial Properties of Green Tea Catechins. Int. J. Mol. Sci. 2020, 21, 1744. [Google Scholar] [CrossRef]
- Kim, M.; Kim, S.Y.; Lee, H.W.; Shin, J.S.; Kim, P.; Jung, Y.S.; Jeong, H.S.; Hyun, J.K.; Lee, C.K. Inhibition of influenza virus internalization by (-)-epigallocatechin-3-gallate. Antivir. Res. 2013, 100, 460–472. [Google Scholar] [CrossRef]
- Song, J.-M.; Lee, K.-H.; Seong, B.-L. Antiviral effect of catechins in green tea on influenza virus. Antivir. Res. 2005, 68, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Onishi, S.; Mori, T.; Kanbara, H.; Habe, T.; Ota, N.; Kurebayashi, Y.; Suzuki, T. Green tea catechins adsorbed on the murine pharyngeal mucosa reduce influenza A virus infection. J. Funct. Foods 2020, 68, 103894. [Google Scholar] [CrossRef]
- Matsuura, R.; Kawamura, A.; Matsumoto, Y.; Iida, Y.; Kanayama, M.; Kurokawa, M.; Aida, Y. Epigallocatechin Gallate Stabilized by Cyclodextrin Inactivates Influenza Virus and Human Coronavirus 229E. Microorganisms 2022, 10, 1796. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.X.; Wei, F.; Li, N.; Li, J.L.; Chen, L.J.; Liu, Y.Y.; Luo, F.; Xiong, H.R.; Hou, W.; Yang, Z.Q. Amelioration of influenza virus-induced reactive oxygen species formation by epigallocatechin gallate derived from green tea. Acta Pharmacol. Sin. 2012, 33, 1533–1541. [Google Scholar] [CrossRef] [PubMed]
- Colpitts, C.C.; Schang, L.M. A small molecule inhibits virion attachment to heparan sulfate- or sialic acid-containing glycans. J. Virol. 2014, 88, 7806–7817. [Google Scholar] [CrossRef] [PubMed]
- Ohishi, T.; Hishiki, T.; Baig, M.S.; Rajpoot, S.; Saqib, U.; Takasaki, T.; Hara, Y. Epigallocatechin gallate (EGCG) attenuates severe acute respiratory coronavirus disease 2 (SARS-CoV-2) infection by blocking the interaction of SARS-CoV-2 spike protein receptor-binding domain to human angiotensin-converting enzyme 2. PLoS ONE 2022, 17, e0271112. [Google Scholar] [CrossRef]
- LeBlanc, E.V.; Colpitts, C.C. The green tea catechin EGCG provides proof-of-concept for a pan-coronavirus attachment inhibitor. Sci. Rep. 2022, 12, 12899. [Google Scholar] [CrossRef]
- Rabezanahary, H.; Badr, A.; Checkmahomed, L.; Pageau, K.; Desjardins, Y.; Baz, M. Epigallocatechin Gallate and Isoquercetin Synergize With Remdesivir to Reduce SARS-CoV-2 Replication In Vitro. Front. Virol. 2022, 2, 956113. [Google Scholar] [CrossRef]
- Liu, J.; Bodnar, B.H.; Meng, F.; Khan, A.I.; Wang, X.; Saribas, S.; Wang, T.; Lohani, S.C.; Wang, P.; Wei, Z.; et al. Epigallocatechin gallate from green tea effectively blocks infection of SARS-CoV-2 and new variants by inhibiting spike binding to ACE2 receptor. Cell Biosci. 2021, 11, 168. [Google Scholar] [CrossRef]
- Ciesek, S.; von Hahn, T.; Colpitts, C.C.; Schang, L.M.; Friesland, M.; Steinmann, J.; Manns, M.P.; Ott, M.; Wedemeyer, H.; Meuleman, P.; et al. The green tea polyphenol, epigallocatechin-3-gallate, inhibits hepatitis C virus entry. Hepatology 2011, 54, 1947–1955. [Google Scholar] [CrossRef]
- Calland, N.; Albecka, A.; Belouzard, S.; Wychowski, C.; Duverlie, G.; Descamps, V.; Hober, D.; Dubuisson, J.; Rouillé, Y.; Séron, K. (−)-Epigallocatechin-3-gallate is a new inhibitor of hepatitis C virus entry. Hepatology 2012, 55, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Jang, Y.H.; Byun, Y.H.; Cheong, Y.; Kim, P.; Lee, Y.J.; Lee, Y.J.; Sung, J.M.; Son, A.; Lee, H.M.; et al. Green Tea Catechin-Inactivated Viral Vaccine Platform. Front. Microbiol. 2017, 8, 2469. [Google Scholar] [CrossRef] [PubMed]
- Peltola, V.T.; Murti, K.G.; McCullers, J.A. Influenza Virus Neuraminidase Contributes to Secondary Bacterial Pneumonia. J. Infect. Dis. 2005, 192, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Rawangkan, A.; Kengkla, K.; Kanchanasurakit, S.; Duangjai, A.; Saokaew, S. Anti-Influenza with Green Tea Catechins: A Systematic Review and Meta-Analysis. Molecules 2021, 26, 4014. [Google Scholar] [CrossRef] [PubMed]
- Umeda, M.; Tominaga, T.; Kozuma, K.; Kitazawa, H.; Furushima, D.; Hibi, M.; Yamada, H. Preventive effects of tea and tea catechins against influenza and acute upper respiratory tract infections: A systematic review and meta-analysis. Eur. J. Nutr. 2021, 60, 4189–4202. [Google Scholar] [CrossRef] [PubMed]
- Furushima, D.; Nishimura, T.; Takuma, N.; Iketani, R.; Mizuno, T.; Matsui, Y.; Yamaguchi, T.; Nakashima, Y.; Yamamoto, S.; Hibi, M.; et al. Prevention of Acute Upper Respiratory Infections by Consumption of Catechins in Healthcare Workers: A Randomized, Placebo-Controlled Trial. Nutrients 2020, 12, 4. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Yamada, H.; Takuma, N.; Niino, H.; Sagesaka, Y.M. Effects of green tea catechins and theanine on preventing influenza infection among healthcare workers: A randomized controlled trial. BMC Complement. Altern. Med. 2011, 11, 15. [Google Scholar] [CrossRef]
- Xu, M.-J.; Liu, B.-J.; Wang, C.-L.; Wang, G.-H.; Tian, Y.; Wang, S.-H.; Li, J.; Li, P.-Y.; Zhang, R.-H.; Wei, D.; et al. Epigallocatechin-3-gallate inhibits TLR4 signaling through the 67-kDa laminin receptor and effectively alleviates acute lung injury induced by H9N2 swine influenza virus. Int. Immunopharmacol. 2017, 52, 24–33. [Google Scholar] [CrossRef]
- Belser, J.A.; Pulit-Penaloza, J.A.; Maines, T.R. Ferreting Out Influenza Virus Pathogenicity and Transmissibility: Past and Future Risk Assessments in the Ferret Model. Cold Spring Harb. Perspect. Med. 2020, 10, a038323. [Google Scholar] [CrossRef]
- Stannard, H.L.; Mifsud, E.J.; Wildum, S.; Brown, S.K.; Koszalka, P.; Shishido, T.; Kojima, S.; Omoto, S.; Baba, K.; Kuhlbusch, K.; et al. Assessing the fitness of a dual-antiviral drug resistant human influenza virus in the ferret model. Commun. Biol. 2022, 5, 1026. [Google Scholar] [CrossRef]
- Koszalka, P.; Tilmanis, D.; Roe, M.; Vijaykrishna, D.; Hurt, A.C. Baloxavir marboxil susceptibility of influenza viruses from the Asia-Pacific, 2012–2018. Antivir. Res. 2019, 164, 91–96. [Google Scholar] [CrossRef]
- Oh, D.Y.; Barr, I.G.; Hurt, A.C. A Novel Video Tracking Method to Evaluate the Effect of Influenza Infection and Antiviral Treatment on Ferret Activity. PLoS ONE 2015, 10, e0118780. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, Y.N.; Youn, H.N.; Lee, D.H.; Kwak, J.H.; Seong, B.L.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Anti-influenza virus activity of green tea by-products in vitro and efficacy against influenza virus infection in chickens. Poult. Sci. 2012, 91, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, U.; Haller, J.; Decourt, J.P.; Girault, N.; Girault, J.; Richard-Caudron, A.S.; Pineau, B.; Weber, P. A single ascending dose study of epigallocatechin gallate in healthy volunteers. J. Int. Med. Res. 2003, 31, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Naumovski, N.; Blades, B.L.; Roach, P.D. Food Inhibits the Oral Bioavailability of the Major Green Tea Antioxidant Epigallocatechin Gallate in Humans. Antioxidants 2015, 4, 373–393. [Google Scholar] [CrossRef] [PubMed]
- Chow, H.H.; Hakim, I.A. Pharmacokinetic and chemoprevention studies on tea in humans. Pharmacol. Res. 2011, 64, 105–112. [Google Scholar] [CrossRef]
- Janle, E.M.; Morré, D.M.; Morré, D.J.; Zhou, Q.; Zhu, Y. Pharmacokinetics of green tea catechins in extract and sustained-release preparations. J. Diet. Suppl. 2008, 5, 248–263. [Google Scholar] [CrossRef]
- Deng, Y.-M.; Caldwell, N.; Hurt, A.; Shaw, T.; Kelso, A.; Chidlow, G.; Williams, S.; Smith, D.; Barr, I. A comparison of pyrosequencing and neuraminidase inhibition assays for the detection of oseltamivir-resistant pandemic influenza A(H1N1) 2009 viruses. Antivir. Res. 2011, 90, 87–91. [Google Scholar] [CrossRef]
- Hein, J.; Spreyer, F.; Sauter-Louis, C.; Hartmann, K. Reference ranges for laboratory parameters in ferrets. Vet. Rec. 2012, 171, 218. [Google Scholar] [CrossRef]
- Yang, Z.-F.; Bai, L.-P.; Huang, W.-b.; Li, X.-Z.; Zhao, S.-S.; Zhong, N.-S.; Jiang, Z.-H. Comparison of in vitro antiviral activity of tea polyphenols against influenza A and B viruses and structure–activity relationship analysis. Fitoterapia 2014, 93, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Furubayashi, T.; Inoue, D.; Nishiyama, N.; Tanaka, A.; Yutani, R.; Kimura, S.; Katsumi, H.; Yamamoto, A.; Sakane, T. Comparison of Various Cell Lines and Three-Dimensional Mucociliary Tissue Model Systems to Estimate Drug Permeability Using an In Vitro Transport Study to Predict Nasal Drug Absorption in Rats. Pharmaceutics 2020, 12, 79. [Google Scholar] [CrossRef] [PubMed]
- Hsu, A.C.; Barr, I.; Hansbro, P.M.; Wark, P.A. Human influenza is more effective than avian influenza at antiviral suppression in airway cells. Am. J. Respir. Cell Mol. Biol. 2011, 44, 906–913. [Google Scholar] [CrossRef] [PubMed]
- Tréhin, R.; Krauss, U.; Beck-Sickinger, A.G.; Merkle, H.P.; Nielsen, H.M. Cellular Uptake But Low Permeation of Human Calcitonin-Derived Cell Penetrating Peptides and Tat(47-57) Through Well-Differentiated Epithelial Models. Pharm. Res. 2004, 21, 1248–1256. [Google Scholar] [CrossRef] [PubMed]
- Marriott, A.C.; Dove, B.K.; Whittaker, C.J.; Bruce, C.; Ryan, K.A.; Bean, T.J.; Rayner, E.; Pearson, G.; Taylor, I.; Dowall, S.; et al. Low Dose Influenza Virus Challenge in the Ferret Leads to Increased Virus Shedding and Greater Sensitivity to Oseltamivir. PLoS ONE 2014, 9, e94090. [Google Scholar] [CrossRef] [PubMed]
- Govorkova, E.A.; Marathe, B.M.; Prevost, A.; Rehg, J.E.; Webster, R.G. Assessment of the efficacy of the neuraminidase inhibitor oseltamivir against 2009 pandemic H1N1 influenza virus in ferrets. Antivir. Res. 2011, 91, 81–88. [Google Scholar] [CrossRef]
- Mifsud, E.J.; Tilmanis, D.; Oh, D.Y.; Ming-Kay Tai, C.; Rossignol, J.F.; Hurt, A.C. Prophylaxis of ferrets with nitazoxanide and oseltamivir combinations is more effective at reducing the impact of influenza a virus infection compared to oseltamivir monotherapy. Antivir. Res. 2020, 176, 104751. [Google Scholar] [CrossRef]
- Mok, C.-K.; Ng, Y.L.; Ahidjo, B.A.; Aw, Z.Q.; Chen, H.; Wong, Y.H.; Lee, R.C.H.; Loe, M.W.C.; Liu, J.; Tan, K.S.; et al. Evaluation of In Vitro and In Vivo Antiviral Activities of Vitamin D for SARS-CoV-2 and Variants. Pharmaceutics 2023, 15, 925. [Google Scholar] [CrossRef]
- Mereles, D.; Hunstein, W. Epigallocatechin-3-gallate (EGCG) for clinical trials: More pitfalls than promises? Int. J. Mol. Sci. 2011, 12, 5592–5603. [Google Scholar] [CrossRef]
- Shammas, M.A.; Neri, P.; Koley, H.; Batchu, R.B.; Bertheau, R.C.; Munshi, V.; Prabhala, R.; Fulciniti, M.; Tai, Y.T.; Treon, S.P.; et al. Specific killing of multiple myeloma cells by (-)-epigallocatechin-3-gallate extracted from green tea: Biologic activity and therapeutic implications. Blood 2006, 108, 2804–2810. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Moreno-Rojas, J.M.; Brindani, N.; Del Rio, D.; Lean, M.E.J.; Hara, Y.; Crozier, A. Bioavailability of Black Tea Theaflavins: Absorption, Metabolism, and Colonic Catabolism. J. Agric. Food Chem. 2017, 65, 5365–5374. [Google Scholar] [CrossRef] [PubMed]
- Sima, M.; Lv, C.; Qi, J.; Guo, J.; Luo, R.; Deng, X.; Li, Y.; Wang, T.; Yue, D.; Gao, Y. Anti-inflammatory effects of theaflavin-3′-gallate during influenza virus infection through regulating the TLR4/MAPK/p38 pathway. Eur. J. Pharmacol. 2022, 938, 175332. [Google Scholar] [CrossRef] [PubMed]
- Aree, T.; Jongrungruangchok, S. Enhancement of antioxidant activity of green tea epicatechins in β-cyclodextrin cavity: Single-crystal X-ray analysis, DFT calculation and DPPH assay. Carbohydr. Polym. 2016, 151, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Meng, X.; Yang, C.S. Enzymology of methylation of tea catechins and inhibition of catechol-O-methyltransferase by (−)-epigallocatechin gallate. Drug Metab. Dispos. Biol. Fate Chem. 2003, 31, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Isbrucker, R.A.; Edwards, J.A.; Wolz, E.; Davidovich, A.; Bausch, J. Safety studies on epigallocatechin gallate (EGCG) preparations. Part 2: Dermal, acute and short-term toxicity studies. Food Chem. Toxicol. 2006, 44, 636–650. [Google Scholar] [CrossRef]
- Chan, P.C.; Ramot, Y.; Malarkey, D.E.; Blackshear, P.; Kissling, G.E.; Travlos, G.; Nyska, A. Fourteen-week toxicity study of green tea extract in rats and mice. Toxicol. Pathol. 2010, 38, 1070–1084. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Katz, J.M.; Tumpey, T.M. The ferret as a model organism to study influenza A virus infection. Dis. Models Mech. 2011, 4, 575–579. [Google Scholar] [CrossRef]
- Belser, J.A.; Eckert, A.M.; Tumpey, T.M.; Maines, T.R. Complexities in Ferret Influenza Virus Pathogenesis and Transmission Models. Microbiol. Mol. Biol. Rev. 2016, 80, 733–744. [Google Scholar] [CrossRef]
- Cheong, Y.; Kim, M.; Ahn, J.; Oh, H.; Lim, J.; Chae, W.; Yang, S.W.; Kim, M.S.; Yu, J.E.; Byun, S.; et al. Epigallocatechin-3-Gallate as a Novel Vaccine Adjuvant. Front. Immunol. 2021, 12, 769088. [Google Scholar] [CrossRef]
- Kumazoe, M.; Takamatsu, K.; Horie, F.; Yoshitomi, R.; Hamagami, H.; Tanaka, H.; Fujimura, Y.; Tachibana, H. Methylated (−)-epigallocatechin 3-O-gallate potentiates the effect of split vaccine accompanied with upregulation of Toll-like receptor 5. Sci. Rep. 2021, 11, 23101. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stannard, H.; Koszalka, P.; Deshpande, N.; Desjardins, Y.; Baz, M. Pre-Clinical Evaluation of the Antiviral Activity of Epigalocatechin-3-Gallate, a Component of Green Tea, against Influenza A(H1N1)pdm Viruses. Viruses 2023, 15, 2447. https://doi.org/10.3390/v15122447
Stannard H, Koszalka P, Deshpande N, Desjardins Y, Baz M. Pre-Clinical Evaluation of the Antiviral Activity of Epigalocatechin-3-Gallate, a Component of Green Tea, against Influenza A(H1N1)pdm Viruses. Viruses. 2023; 15(12):2447. https://doi.org/10.3390/v15122447
Chicago/Turabian StyleStannard, Harry, Paulina Koszalka, Nikita Deshpande, Yves Desjardins, and Mariana Baz. 2023. "Pre-Clinical Evaluation of the Antiviral Activity of Epigalocatechin-3-Gallate, a Component of Green Tea, against Influenza A(H1N1)pdm Viruses" Viruses 15, no. 12: 2447. https://doi.org/10.3390/v15122447