
Gene-Editing and RNA Interference in Treating Hepatitis B: A Review

 , , , , and
, , , , and 
Abstract
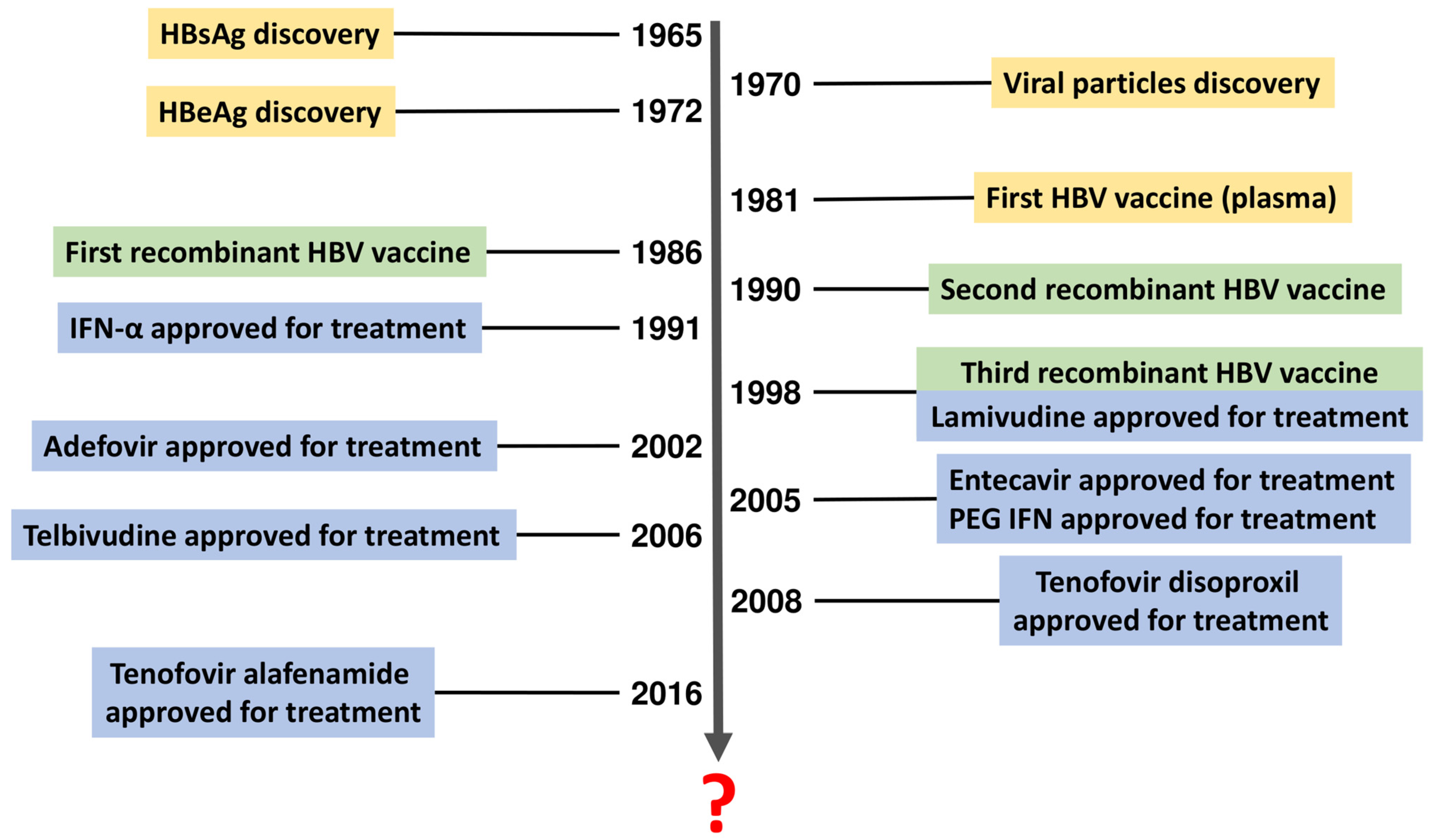
:1. Introduction
2. Current Options in HBV Treatment
2.1. Pegylated Interferon- α2a (PEG-IFN-α2a) in Chronic Hepatitis B Treatment
2.2. Nucleos(t)ide Analogs (NAs) in Chronic Hepatitis B Treatment
3. Biotechnological Approach to HBV Treatment
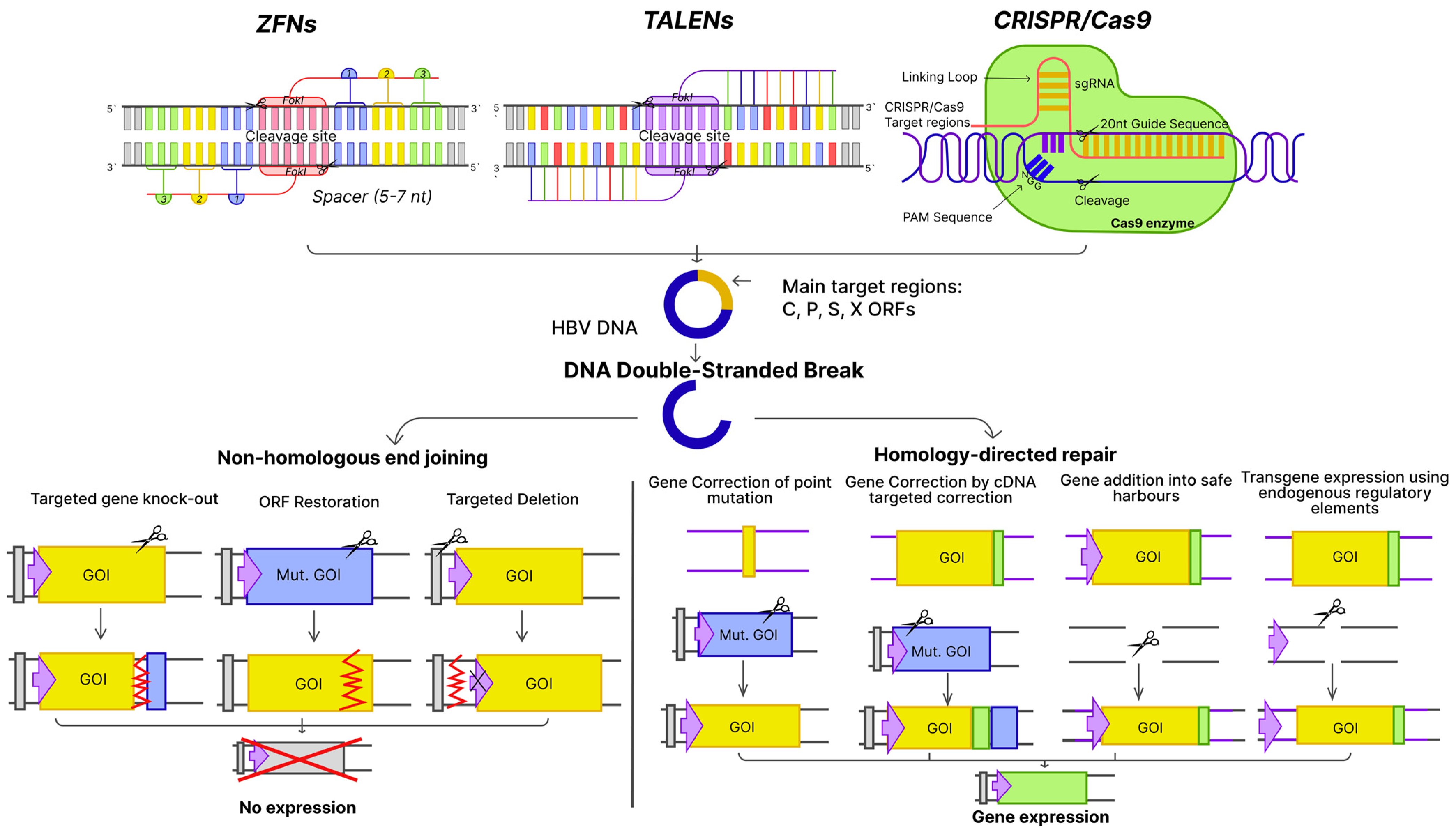
3.1. Gene-Editing Tools
3.1.1. Zinc-Finger Proteins (ZFPs)
3.1.2. Transcription Activator-like Effector Nucleases (TALENs)
3.1.3. The Clustered Regularly Interspaced Palindromic Repeat-Cas System (CRISPR/Cas)
3.2. RNA Interference Therapeutics

{kind=link}
{kind=link}
| Candidate | Phase | Targets | Key Findings | Disadvantages and Adverse Events | References |
|---|---|---|---|---|---|
| ALG-125755 | 1 | Single trigger within the ORF of HBV S | (1) Well tolerated with good safety profile of single dose in BeAg negative individuals (2) Dose-dependent reductions in HBsAg over 90 days after a single dose | (1) Discontinued due to inconclusive efficacy results in human trial | [166,167,168] |
| ARC-520 | 2 | Two triggers within the ORF of HBV X protein | (1) Prolonged HBsAg reduction (2) Targets all cccDNA-derived viral transcripts, preventing protein translation, reducing viral antigens and HBV DNA | (1) Limited effect on integrated HBV DNA (2) Differential response in patient populations (3) Terminated after mortality induced by the excipient in non-human primates was reported (4) Dosing in these studies was very high, but at normal dosing, no HBsAg response was observed | [81,138,164,169] |
| ARB-1467 | 2a | Three triggers targeting the ORF of HBV S and HBV X domains | (1) Single and multiple doses induced HBsAg decline independent of HBeAg status (2) HBcrAg and HBsAg reductions were observed following multiple doses. (3) No serious adverse events; no increase in ALT levels | [141,142,170] | |
| AB-729 | 2 | HBV X | (1) Targets both sources of HBsAg (cccDNA-derived and integrated HBV DNA) (2) Significant decline of HBsAg (3) Well tolerated alone and in combination with IFN | (1) The decrease in total HBsAg clearly correlated with declines over time in M-HBsAg and L-HBsAg and not with S-HBsAg | [171,172] |
| VIR-2218 | 2 | One trigger targeting the ORF of X protein | (1) HBsAg decline persisting for 28 weeks after treatment (2) Reductions in HBsAg in HBeAg− and HBeAg+ patients across all dose levels (3) Well tolerated, no significant increase in ALT level | (1) Differential patterns of decline suggest that early responses (<8 weeks) in HBsAg may not predict the final magnitude of decline. | [148,173] |
| JNJ-3989 | 2 | HBV X and HBV S domains | (1) Targets both sources of HBsAg (cccDNA and integrated HBV DNA) (2) Regardless of HBe serostatus, three monthly doses combined with daily NUC treatment resulted in reduced HBsAg, HBV DNA, HBV RNA, and HBcAg levels (3) 56% of patients achieved a sustained HBsAg response 9 months after treatment (4) Well tolerated, with mild-to-moderate adverse events | [149,174,175,176] | |
| ARB-1740 | 1a/b | 3 triggers that target three highly conserved regions of the HBV genome | (1) HBV RNA reduction, leading to inhibition of HBsAg, HBeAg, and HBcAg (2) Synergistic activity against HBV relaxed circular DNA when used in combination with a capsid inhibitor and/or pegylated interferon-alpha (3) May have therapeutic benefits for hepatitis delta patients | (1) Suspended due to transferring the liver-targeted delivery system from liposomes to GalNAc–siRNA conjugates. | [144,177] |
| ARC-521 | 1 | A conserved sequence in HBV X and S ORFs | (1) Targets cccDNA and integrated DNA (2) Potent reductions in HBsAg and HBV DNA (3) Silenced all HBV gene products (4) Suppression of HBeAg and HBsAg suppression (5) Well tolerated in human participants | (1) Deaths in a non-clinical toxicology study in non-human primates (related to drug EX1 delivery vehicle) | [139,178] |
| GSK3228836 (bepirovirsen) | 2b | All HBV mRNAs | (1) Sustained HBsAg and HBV DNA loss in 9 to 10% of participants after 300 mg injections weekly for 24 weeks | (1) HBsAg reduction only in patients with low HBsAg (<1000 IU/mL) at baseline (2) Increased levels of ALT (3) Injection-site reactions, pyrexia, and fatigue were common | [179,180] |
| GSK3389404 (bepirovirsen conjugated to GalNAc) | 2a | Pregenomic and mRNA transcripts | (1) Dose-dependent reduction in HBsAg after 12 weeks of treatment (2) Reduction in HBsAg in HBeAg+ and HBeAg- patients (3) Good safety profile | (1) Only 5% had a >1.5-log reduction in HBsAg with no HBsAg seroclearance achieved | [181,182] |
| RO7062931 | 1 | A highly conserved sequence in the shared 3′ region | (1) Dose-dependent declines in HBsAg demonstrated target engagement (2) Good safety profile and tolerance | (1) Suboptimal magnitude of HBsAg reduction (2) Further development halted | [183] |
| RG6346 | 1 | HBV S domain | (1) Four monthly doses resulted in substantial and durable reductions in HBsAg levels lasting up to 1 year following the last dose (2) Well-tolerated, mild, local adverse reactions | [184] |
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsai, E. Review of Current and Potential Treatments for Chronic Hepatitis B Virus Infection. Gastroenterol. Hepatol. 2021, 17, 367–376. [Google Scholar]
- Zhao, K.; Liu, A.; Xia, Y. Insights into Hepatitis B Virus DNA Integration-55 Years after Virus Discovery. Innovation 2020, 1, 100034. [Google Scholar] [CrossRef]
- Huang, Q.; Zhou, B.; Cai, D.; Zong, Y.; Wu, Y.; Liu, S.; Mercier, A.; Guo, H.; Hou, J.; Colonno, R.; et al. Rapid Turnover of Hepatitis B Virus Covalently Closed Circular DNA Indicated by Monitoring Emergence and Reversion of Signature-Mutation in Treated Chronic Hepatitis B Patients. Hepatology 2021, 73, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Razavi-Shearer, D.; Gamkrelidze, I.; Pan, C.; Jia, J.; Berg, T.; Gray, R.; Lim, Y.-S.; Chen, C.-J.; Ocama, P.; Desalegn, H.; et al. Global Prevalence, Cascade of Care, and Prophylaxis Coverage of Hepatitis B in 2022: A Modelling Study. Lancet Gastroenterol. Hepatol. 2023, 8, 879–907. [Google Scholar] [CrossRef] [PubMed]
- WHO. Hepatitis B. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-b (accessed on 14 May 2023).
- Yuen, M.-F.; Chen, D.-S.; Dusheiko, G.M.; Janssen, H.L.A.; Lau, D.T.Y.; Locarnini, S.A.; Peters, M.G.; Lai, C.-L. Hepatitis B Virus Infection. Nat. Rev. Dis. Primers 2018, 4, 18035. [Google Scholar] [CrossRef]
- Bower, W.A.; Finelli, L.; Goldstein, S.T. Hepatitis B and Hepatitis D Viruses. In Principles and Practice of Pediatric Infectious Disease; Long, S.S., Ed.; Elsevier: Toronto, ON, Canada, 2008; pp. 1062–1072. ISBN 9780702034688. [Google Scholar]
- Kappus, M.R.; Sterling, R.K. Extrahepatic Manifestations of Acute Hepatitis B Virus Infection. Gastroenterol. Hepatol. 2013, 9, 123–126. [Google Scholar]
- Rzymski, P.; Zarębska-Michaluk, D.; Flisiak, R. Could Chronic HBV Infection Explain Beethoven’s Hearing Loss? Implications for Patients Currently Living with Hepatitis B. J. Infect. 2023, 87, 171–176. [Google Scholar] [CrossRef]
- Mårdh, O.; Quinten, C.; Amato-Gauci, A.J.; Duffell, E. Mortality from Liver Diseases Attributable to Hepatitis B and C in the EU/EEA—Descriptive Analysis and Estimation of 2015 Baseline. Infect. Dis. 2020, 52, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Guan, L.; Tian, H.; Zeng, Z.; Chen, J.; Huang, D.; Sun, J.; Guo, J.; Cui, H.; Li, Y. Risk Factors and Prevention of Viral Hepatitis-Related Hepatocellular Carcinoma. Front. Oncol. 2021, 11, 686962. [Google Scholar] [CrossRef]
- Total Burden Related to Hepatitis B—Cause. Available online: https://www.healthdata.org/results/gbd_summaries/2019/total-burden-related-hepatitis-b-cause (accessed on 5 September 2023).
- Zhang, C.; Liu, Y.; Zhao, H.; Wang, G. Global Patterns and Trends in Total Burden of Hepatitis B from 1990 to 2019 and Predictions to 2030. Clin. Epidemiol. 2022, 14, 1519–1533. [Google Scholar] [CrossRef]
- Trickey, A.; Bivegete, S.; Duffell, E.; McNaughton, A.L.; Nerlander, L.; Walker, J.G.; Fraser, H.; Hickman, M.; Vickerman, P.; Brooks-Pollock, E.; et al. Estimating Hepatitis B Virus Prevalence among Key Population Groups for European Union and European Economic Area Countries and the United Kingdom: A Modelling Study. BMC Infect. Dis. 2023, 23, 457. [Google Scholar] [CrossRef]
- Ott, J.J.; Horn, J.; Krause, G.; Mikolajczyk, R.T. Time Trends of Chronic HBV Infection over Prior Decades—A Global Analysis. J. Hepatol. 2017, 66, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Velkov, S.; Ott, J.; Protzer, U.; Michler, T. The Global Hepatitis B Virus Genotype Distribution Approximated from Available Genotyping Data. Genes 2018, 9, 495. [Google Scholar] [CrossRef]
- Reuter, T.Q.; Gomes-Gouvea, M.; Chuffi, S.; Duque, U.H.; Carvalho, J.A.; Perini, W.; Queiroz, M.M.; Segal, I.M.; Azevedo, R.S.; Pinho, J.R.R. Hepatitis B Virus Genotypes and Subgenotypes and the Natural History and Epidemiology of Hepatitis B. Ann. Hepatol. 2022, 27, 100574. [Google Scholar] [CrossRef] [PubMed]
- Świderska, M.; Pawłowska, M.; Mazur, W.; Tomasiewicz, K.; Simon, K.; Piekarska, A.; Wawrzynowicz-Syczewska, M.; Jaroszewicz, J.; Rajewski, P.; Zasik, E.; et al. Original Article Distribution of HBV Genotypes in Poland. Clin. Exp. Hepatol. 2015, 1, 1–4. [Google Scholar] [CrossRef]
- Genowska, A.; Zarębska-Michaluk, D.; Tyszko, P.; Strukcinskiene, B.; Moniuszko-Malinowska, A.; Rzymski, P.; Flisiak, R. Trends of Infections and Mortality Due to Hepatitis B Virus (2005–2022) and the Potential Impact of the COVID-19 Pandemic: A Population-Based Study in Poland. Clin. Exp. Hepatol. 2023, 9, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Sunbul, M. Hepatitis B Virus Genotypes: Global Distribution and Clinical Importance. World J. Gastroenterol. 2014, 20, 5427. [Google Scholar] [CrossRef] [PubMed]
- Farci, P.; Smedile, A.; Lavarini, C.; Piantino, P.; Crivelli, O.; Caporaso, N.; Toti, M.; Bonino, F.; Rizzetto, M. Delta Hepatitis in Inapparent Carriers of Hepatitis B Surface Antigen. Gastroenterology 1983, 85, 669–673. [Google Scholar] [CrossRef]
- Sureau, C.; Negro, F. The Hepatitis Delta Virus: Replication and Pathogenesis. J. Hepatol. 2016, 64, S102–S116. [Google Scholar] [CrossRef]
- Sagnelli, C.; Sagnelli, E.; Russo, A.; Pisaturo, M.; Occhiello, L.; Coppola, N. HBV/HDV Co-Infection: Epidemiological and Clinical Changes, Recent Knowledge and Future Challenges. Life 2021, 11, 169. [Google Scholar] [CrossRef]
- Li, X.; Mukandavire, C.; Cucunubá, Z.M.; Echeverria Londono, S.; Abbas, K.; Clapham, H.E.; Jit, M.; Johnson, H.L.; Papadopoulos, T.; Vynnycky, E.; et al. Estimating the Health Impact of Vaccination against Ten Pathogens in 98 Low-Income and Middle-Income Countries from 2000 to 2030: A Modelling Study. Lancet 2021, 397, 398–408. [Google Scholar] [CrossRef]
- Ozawa, S.; Clark, S.; Portnoy, A.; Grewal, S.; Stack, M.L.; Sinha, A.; Mirelman, A.; Franklin, H.; Friberg, I.K.; Tam, Y.; et al. Estimated Economic Impact of Vaccinations in 73 Low- and Middle-Income Countries, 2001–2020. Bull. World Health Organ. 2017, 95, 629–638. [Google Scholar] [CrossRef]
- Flores, J.E.; Thompson, A.J.; Ryan, M.; Howell, J. The Global Impact of Hepatitis B Vaccination on Hepatocellular Carcinoma. Vaccines 2022, 10, 793. [Google Scholar] [CrossRef] [PubMed]
- Nayagam, S.; Thursz, M.; Sicuri, E.; Conteh, L.; Wiktor, S.; Low-Beer, D.; Hallett, T.B. Requirements for Global Elimination of Hepatitis B: A Modelling Study. Lancet Infect. Dis. 2016, 16, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Lancet. The Lancet Viral Hepatitis Elimination: A Challenge, but within Reach. Lancet 2022, 400, 251. [Google Scholar] [CrossRef]
- Lampertico, P.; Agarwal, K.; Berg, T.; Buti, M.; Janssen, H.L.; Papatheodoridis, G.; Zoulim, F.; Tacke, F. EASL 2017 Clinical Practice Guidelines on the Management of Hepatitis B Virus Infection. J. Hepatol. 2017, 67, 370–398. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y. Obstacles and Opportunities in the Prevention and Treatment of HBV-Related Hepatocellular Carcinoma. Genes Dis. 2020, 7, 291–298. [Google Scholar] [CrossRef] [PubMed]
- EMA. Hepcludex. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/hepcludex (accessed on 26 October 2023).
- Wedemeyer, H.; Aleman, S.; Brunetto, M.R.; Blank, A.; Andreone, P.; Bogomolov, P.; Chulanov, V.; Mamonova, N.; Geyvandova, N.; Morozov, V.; et al. A Phase 3, Randomized Trial of Bulevirtide in Chronic Hepatitis D. N. Engl. J. Med. 2023, 389, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Gavidia, M. FDA Rejects Bulevirtide for Hepatitis D, Compensated Liver Disease. Available online: https://www.ajmc.com/view/fda-rejects-bulevirtide-for-hepatitis-d-compensated-liver-disease (accessed on 26 October 2023).
- Liao, C.; Xiao, S.; Wang, X. Bench-to-Bedside: Translational Development Landscape of Biotechnology in Healthcare. Health Sci. Rev. 2023, 7, 100097. [Google Scholar] [CrossRef]
- Rzymski, P.; Szuster-Ciesielska, A.; Dzieciątkowski, T.; Gwenzi, W.; Fal, A. MRNA Vaccines: The Future of Prevention of Viral Infections? J. Med. Virol. 2023, 95, e28572. [Google Scholar] [CrossRef]
- Barbier, A.J.; Jiang, A.Y.; Zhang, P.; Wooster, R.; Anderson, D.G. The Clinical Progress of MRNA Vaccines and Immunotherapies. Nat. Biotechnol. 2022, 40, 840–854. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, K.; Rzymski, P.; Islam, M.S.; Makuku, R.; Mushtaq, A.; Khan, A.; Ivanovska, M.; Makka, S.A.; Hashem, F.; Marquez, L.; et al. COVID-19 Vaccinations: The Unknowns, Challenges, and Hopes. J. Med. Virol. 2021, 94, 1336–1349. [Google Scholar] [CrossRef] [PubMed]
- Watson, O.J.; Barnsley, G.; Toor, J.; Hogan, A.B.; Winskill, P.; Ghani, A.C. Global Impact of the First Year of COVID-19 Vaccination: A Mathematical Modelling Study. Lancet Infect. Dis. 2022, 22, 1293–1302. [Google Scholar] [CrossRef]
- COVID-19 Vaccine Doses Administered by Manufacturer. Available online: https://ourworldindata.org/grapher/covid-vaccine-doses-by-manufacturer (accessed on 25 November 2023).
- Zheng, C.; Shao, W.; Chen, X.; Zhang, B.; Wang, G.; Zhang, W. Real-World Effectiveness of COVID-19 Vaccines: A Literature Review and Meta-Analysis. Int. J. Infect. Dis. 2022, 114, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Study of EBT-101 in Aviremic HIV-1 Infected Adults on Stable ART. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT05144386 (accessed on 6 September 2023).
- Yang, Z.-T.; Huang, S.-Y.; Chen, L.; Liu, F.; Cai, X.-H.; Guo, Y.-F.; Wang, M.-J.; Han, Y.; Yu, D.-M.; Jiang, J.-H.; et al. Characterization of Full-Length Genomes of Hepatitis B Virus Quasispecies in Sera of Patients at Different Phases of Infection. J. Clin. Microbiol. 2015, 53, 2203–2214. [Google Scholar] [CrossRef]
- Zhou, T.-C.; Li, X.; Li, L.; Li, X.-F.; Zhang, L.; Wei, J. Evolution of Full-Length Genomes of HBV Quasispecies in Sera of Patients with a Coexistence of HBsAg and Anti-HBs Antibodies. Sci. Rep. 2017, 7, 661. [Google Scholar] [CrossRef]
- Gencay, M.; Hübner, K.; Gohl, P.; Seffner, A.; Weizenegger, M.; Neofytos, D.; Batrla, R.; Woeste, A.; Kim, H.-S.; Westergaard, G.; et al. Ultra-Deep Sequencing Reveals High Prevalence and Broad Structural Diversity of Hepatitis B Surface Antigen Mutations in a Global Population. PLoS ONE 2017, 12, e0172101. [Google Scholar] [CrossRef]
- Lin, S.-R.; Yang, T.-Y.; Peng, C.-Y.; Lin, Y.-Y.; Dai, C.-Y.; Wang, H.-Y.; Su, T.-H.; Tseng, T.-C.; Liu, I.-J.; Cheng, H.-R.; et al. Whole Genome Deep Sequencing Analysis of Viral Quasispecies Diversity and Evolution in HBeAg Seroconverters. JHEP Rep. 2021, 3, 100254. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, Y.; Li, L.; Song, Y.; Zhai, X.; Liu, J.; Duan, Z.; Yan, L.; Ding, F.; Liu, J.; et al. The Dynamic Changes of HBV Quasispecies Diversity in Infancy after Immunoprophylaxis Failure: A Prospective Cohort Study. Virol. J. 2021, 18, 236. [Google Scholar] [CrossRef]
- Chen, Q.-Y.; Jia, H.-H.; Wang, X.-Y.; Shi, Y.-L.; Zhang, L.-J.; Hu, L.-P.; Wang, C.; He, X.; Harrison, T.J.; Jackson, J.B.; et al. Analysis of Entire Hepatitis B Virus Genomes Reveals Reversion of Mutations to Wild Type in Natural Infection, a 15 Year Follow-up Study. Infect. Genet. Evol. 2022, 97, 105184. [Google Scholar] [CrossRef]
- Audisio, K.; Lia, H.; Robinson, N.; Rahouma, M.; Soletti, G.; Cancelli, G.; Perezgrovas Olaria, R.; Chadow, D.; Tam, D.; Vervoort, D.; et al. Impact of the COVID-19 Pandemic on Non-COVID-19 Clinical Trials. J. Cardiovasc. Dev. Dis. 2022, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.; Chan, A.Y.; Diaz Peralta, P.; Jin, L.; Pereira Nunes, C.R.; Bell, M.L. Impacts of the COVID-19 Pandemic on Scientists’ Productivity in Science, Technology, Engineering, Mathematics (STEM), and Medicine Fields. Humanit. Soc. Sci. Commun. 2022, 9, 434. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Pokorska-Śpiewak, M.; Jackowska, T.; Kuchar, E.; Nitsch-Osuch, A.; Pawłowska, M.; Babicki, M.; Jaroszewicz, J.; Szenborn, L.; Wysocki, J.; et al. Key Essentials during the Transition from the Acute Phase of the COVID-19 Pandemic. Vaccines 2023, 11, 1502. [Google Scholar] [CrossRef] [PubMed]
- Flisiak, R.; Halota, W.; Jaroszewicz, J.; Juszczyk, J.; Małkowski, P.; Pawłowska, M.; Piekarska, A.; Simon, K.; Tomasiewicz, K.; Wawrzynowicz-Syczewska, M. Recommendations for the Treatment of Hepatitis B in 2017. Clin. Exp. Hepatol. 2017, 2, 35–46. [Google Scholar] [CrossRef]
- Terrault, N.A.; Lok, A.S.F.; McMahon, B.J.; Chang, K.-M.; Hwang, J.P.; Jonas, M.M.; Brown, R.S.; Bzowej, N.H.; Wong, J.B. Update on Prevention, Diagnosis, and Treatment of Chronic Hepatitis B: AASLD 2018 Hepatitis B Guidance. Hepatology 2018, 67, 1560–1599. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Wang, F.; Li, T.; Xu, X.; Sun, Y.; Nan, Y.; Wang, G.; Hou, J.; Duan, Z.; Wei, L.; et al. Guidelines for the Prevention and Treatment of Chronic Hepatitis B (Version 2022). J. Clin. Transl. Hepatol. 2023, 3, 145–162. [Google Scholar] [CrossRef]
- Viganò, M.; Grossi, G.; Loglio, A.; Lampertico, P. Treatment of Hepatitis B: Is There Still a Role for Interferon? Liver Int. 2018, 38, 79–83. [Google Scholar] [CrossRef]
- Belloni, L.; Allweiss, L.; Guerrieri, F.; Pediconi, N.; Volz, T.; Pollicino, T.; Petersen, J.; Raimondo, G.; Dandri, M.; Levrero, M. IFN-α Inhibits HBV Transcription and Replication in Cell Culture and in Humanized Mice by Targeting the Epigenetic Regulation of the Nuclear CccDNA Minichromosome. J. Clin. Investig. 2012, 122, 529–537. [Google Scholar] [CrossRef]
- Woo, A.S.J.; Kwok, R.; Ahmed, T. Alpha-Interferon Treatment in Hepatitis B. Ann. Transl. Med. 2017, 5, 159. [Google Scholar] [CrossRef]
- Kim, S.S.; Cheong, J.Y.; Cho, S.W. Current Nucleos(t)Ide Analogue Therapy for Chronic Hepatitis B. Gut Liver 2011, 5, 278–287. [Google Scholar] [CrossRef]
- Jaroszewicz, J.; Ho, H.; Markova, A.; Deterding, K.; Wursthorn, K.; Schulz, S.; Bock, C.-T.; Tillmann, H.L.; Manns, M.P.; Wedemeyer, H.; et al. Hepatitis B Surface Antigen (HBsAg) Decrease and Serum Interferon-Inducible Protein-10 Levels as Predictive Markers for HBsAg Loss during Treatment with Nucleoside/Nucleotide Analogues. Antivir. Ther. 2011, 16, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Han, Z.; Liu, J.; Xue, J.; Zhang, S.; Zhu, Z.; Xia, J.; Huang, M. Comparing Efficacy of Lamivudine, Adefovir Dipivoxil, Telbivudine, and Entecavir in Treating Nucleoside Analogues Naïve for HBeAg-Negative Hepatitis B with Medium Hepatitis B Virus (HBV) DNA Levels. Med. Sci. Monit. 2017, 23, 5230–5236. [Google Scholar] [CrossRef] [PubMed]
- Seto, W.-K.; Lam, Y.-F.; Fung, J.; Wong, D.K.-H.; Huang, F.-Y.; Hung, I.F.-N.; Lai, C.-L.; Yuen, M.-F. Changes of HBsAg and HBV DNA Levels in Chinese Chronic Hepatitis B Patients after 5 Years of Entecavir Treatment. J. Gastroenterol. Hepatol. 2014, 29, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Sun, Q.; Gu, J.; Cen, S.; Zhang, Q. Characterization of the Tenofovir Resistance-Associated Mutations in the Hepatitis B Virus Isolates across Genotypes A to D. Antivir. Res. 2022, 203, 105348. [Google Scholar] [CrossRef] [PubMed]
- Buti, M.; Tsai, N.; Petersen, J.; Flisiak, R.; Gurel, S.; Krastev, Z.; Aguilar Schall, R.; Flaherty, J.F.; Martins, E.B.; Charuworn, P.; et al. Seven-Year Efficacy and Safety of Treatment with Tenofovir Disoproxil Fumarate for Chronic Hepatitis B Virus Infection. Dig. Dis. Sci. 2015, 60, 1457–1464. [Google Scholar] [CrossRef]
- Buti, M.; Gane, E.; Seto, W.K.; Chan, H.L.Y.; Chuang, W.-L.; Stepanova, T.; Hui, A.-J.; Lim, Y.-S.; Mehta, R.; Janssen, H.L.A.; et al. Tenofovir Alafenamide versus Tenofovir Disoproxil Fumarate for the Treatment of Patients with HBeAg-Negative Chronic Hepatitis B Virus Infection: A Randomised, Double-Blind, Phase 3, Non-Inferiority Trial. Lancet Gastroenterol. Hepatol. 2016, 1, 196–206. [Google Scholar] [CrossRef]
- Marcellin, P.; Wong, D.K.; Sievert, W.; Buggisch, P.; Petersen, J.; Flisiak, R.; Manns, M.; Kaita, K.; Krastev, Z.; Lee, S.S.; et al. Ten-year Efficacy and Safety of Tenofovir Disoproxil Fumarate Treatment for Chronic Hepatitis B Virus Infection. Liver Int. 2019, 39, 1868–1875. [Google Scholar] [CrossRef]
- Ono, A.; Suzuki, F.; Kawamura, Y.; Sezaki, H.; Hosaka, T.; Akuta, N.; Kobayashi, M.; Suzuki, Y.; Saitou, S.; Arase, Y.; et al. Long-Term Continuous Entecavir Therapy in Nucleos(t)Ide-Naïve Chronic Hepatitis B Patients. J. Hepatol. 2012, 57, 508–514. [Google Scholar] [CrossRef]
- Petersen, J.; Heyne, R.; Mauss, S.; Schlaak, J.; Schiffelholz, W.; Eisenbach, C.; Hartmann, H.; Wiese, M.; Boeker, K.; Loehr, H.-F.; et al. Effectiveness and Safety of Tenofovir Disoproxil Fumarate in Chronic Hepatitis B: A 3-Year Prospective Field Practice Study in Germany. Dig. Dis. Sci. 2016, 61, 3061–3071. [Google Scholar] [CrossRef]
- Ahn, S.H.; Marcellin, P.; Ma, X.; Caruntu, F.A.; Tak, W.Y.; Elkhashab, M.; Chuang, W.-L.; Tabak, F.; Mehta, R.; Petersen, J.; et al. Hepatitis B Surface Antigen Loss with Tenofovir Disoproxil Fumarate plus Peginterferon Alfa-2a: Week 120 Analysis. Dig. Dis. Sci. 2018, 63, 3487–3497. [Google Scholar] [CrossRef]
- Kao, J.-H.; Jeng, W.-J.; Ning, Q.; Su, T.-H.; Tseng, T.-C.; Ueno, Y.; Yuen, M.-F. APASL Guidance on Stopping Nucleos(t)Ide Analogues in Chronic Hepatitis B Patients. Hepatol. Int. 2021, 15, 833–851. [Google Scholar] [CrossRef] [PubMed]
- Manolakopoulos, S.; Kranidioti, H.; Kourikou, A.; Deutsch, M.-M.; Triantos, C.; Tsolias, C.; Manesis, E.K.; Mathou, N.; Alexopoulou, A.; Hadziyannis, E.; et al. Long-term Clinical Outcome of HBeAg-negative Chronic Hepatitis B Patients Who Discontinued Nucleos(t)Ide Analogues. Liver Int. 2021, 41, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Hirode, G.; Choi, H.S.J.; Chen, C.-H.; Su, T.-H.; Seto, W.-K.; Van Hees, S.; Papatheodoridi, M.; Lens, S.; Wong, G.; Brakenhoff, S.M.; et al. Off-Therapy Response after Nucleos(t)Ide Analogue Withdrawal in Patients with Chronic Hepatitis B: An International, Multicenter, Multiethnic Cohort (RETRACT-B Study). Gastroenterology 2022, 162, 757–771.e4. [Google Scholar] [CrossRef] [PubMed]
- van Bömmel, F.; Stein, K.; Heyne, R.; Petersen, J.; Buggisch, P.; Berg, C.; Zeuzem, S.; Stallmach, A.; Sprinzl, M.; Schott, E.; et al. A Multicenter Randomized-Controlled Trial of Nucleos(t)Ide Analogue Cessation in HBeAg-Negative Chronic Hepatitis B. J. Hepatol. 2023, 78, 926–936. [Google Scholar] [CrossRef]
- Berg, T.; Simon, K.-G.; Mauss, S.; Schott, E.; Heyne, R.; Klass, D.M.; Eisenbach, C.; Welzel, T.M.; Zachoval, R.; Felten, G.; et al. Long-Term Response after Stopping Tenofovir Disoproxil Fumarate in Non-Cirrhotic HBeAg-Negative Patients—FINITE Study. J. Hepatol. 2017, 67, 918–924. [Google Scholar] [CrossRef]
- Papatheodoridis, G.V.; Manolakopoulos, S.; Su, T.-H.; Siakavellas, S.; Liu, C.-J.; Kourikou, A.; Yang, H.-C.; Kao, J.-H. Significance of Definitions of Relapse after Discontinuation of Oral Antivirals in HBeAg-negative Chronic Hepatitis B. Hepatology 2018, 68, 415–424. [Google Scholar] [CrossRef] [PubMed]
- van Helden, J.; Weiskirchen, R. Diagnostischer Wert der quantitativen HBsAg-Bestimmung bei der Hepatitis B-Infektion. Z. Gastroenterol. 2016, 54, 319–326. [Google Scholar] [CrossRef]
- Maasoumy, B.; Wiegand, S.B.; Jaroszewicz, J.; Bremer, B.; Lehmann, P.; Deterding, K.; Taranta, A.; Manns, M.P.; Wedemeyer, H.; Glebe, D.; et al. Hepatitis B Core-Related Antigen (HBcrAg) Levels in the Natural History of Hepatitis B Virus Infection in a Large European Cohort Predominantly Infected with Genotypes A and D. Clin. Microbiol. Infect. 2015, 21, 606.e1–606.e10. [Google Scholar] [CrossRef]
- Inoue, T.; Tanaka, Y. The Role of Hepatitis B Core-Related Antigen. Genes 2019, 10, 357. [Google Scholar] [CrossRef]
- Inoue, T.; Kusumoto, S.; Iio, E.; Ogawa, S.; Suzuki, T.; Yagi, S.; Kaneko, A.; Matsuura, K.; Aoyagi, K.; Tanaka, Y. Clinical Efficacy of a Novel, High-Sensitivity HBcrAg Assay in the Management of Chronic Hepatitis B and HBV Reactivation. J. Hepatol. 2021, 75, 302–310. [Google Scholar] [CrossRef]
- Carey, I.; Gersch, J.; Wang, B.; Moigboi, C.; Kuhns, M.; Cloherty, G.; Dusheiko, G.; Agarwal, K. Pregenomic HBV RNA and Hepatitis B Core-related Antigen Predict Outcomes in Hepatitis B e Antigen–Negative Chronic Hepatitis B Patients Suppressed on Nucleos(T)Ide Analogue Therapy. Hepatology 2020, 72, 42–57. [Google Scholar] [CrossRef] [PubMed]
- Tu, T.; Budzinska, M.; Shackel, N.; Urban, S. HBV DNA Integration: Molecular Mechanisms and Clinical Implications. Viruses 2017, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Sung, W.-K.; Zheng, H.; Li, S.; Chen, R.; Liu, X.; Li, Y.; Lee, N.P.; Lee, W.H.; Ariyaratne, P.N.; Tennakoon, C.; et al. Genome-Wide Survey of Recurrent HBV Integration in Hepatocellular Carcinoma. Nat. Genet. 2012, 44, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Wooddell, C.I.; Yuen, M.-F.; Chan, H.L.-Y.; Gish, R.G.; Locarnini, S.A.; Chavez, D.; Ferrari, C.; Given, B.D.; Hamilton, J.; Kanner, S.B.; et al. RNAi-Based Treatment of Chronically Infected Patients and Chimpanzees Reveals That Integrated Hepatitis B Virus DNA Is a Source of HBsAg. Sci. Transl. Med. 2017, 9, eaan0241. [Google Scholar] [CrossRef] [PubMed]
- Watashi, K.; Urban, S.; Li, W.; Wakita, T. NTCP and beyond: Opening the Door to Unveil Hepatitis B Virus Entry. Int. J. Mol. Sci. 2014, 15, 2892–2905. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Li, S.; Liang, W. Bona Fide Receptor for Hepatitis B and D Viral Infections: Mechanism, Research Models and Molecular Drug Targets. Emerg. Microbes Infect. 2018, 7, 134. [Google Scholar] [CrossRef]
- Wu, Q.; Glitscher, M.; Tonnemacher, S.; Schollmeier, A.; Raupach, J.; Zahn, T.; Eberle, R.; Krijnse-Locker, J.; Basic, M.; Hildt, E. Presence of Intact Hepatitis B Virions in Exosomes. Cell. Mol. Gastroenterol. Hepatol. 2023, 15, 237–259. [Google Scholar] [CrossRef]
- Wang, J.; Cao, D.; Yang, J. Exosomes in Hepatitis B Virus Transmission and Related Immune Response. Tohoku J. Exp. Med. 2020, 252, 309–320. [Google Scholar] [CrossRef]
- Sioson, V.A.; Kim, M.; Joo, J. Challenges in Delivery Systems for CRISPR-Based Genome Editing and Opportunities of Nanomedicine. Biomed. Eng. Lett. 2021, 11, 217–233. [Google Scholar] [CrossRef]
- Raguram, A.; Banskota, S.; Liu, D.R. Therapeutic In Vivo Delivery of Gene Editing Agents. Cell 2022, 185, 2806–2827. [Google Scholar] [CrossRef]
- Costa, J.R.; Bejcek, B.E.; McGee, J.E.; Fogel, A.I.; Brimacombe, K.R.; Ketteler, R. Genome Editing Using Engineered Nucleases and Their Use in Genomic Screening; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda MD, USA, 2017. [Google Scholar]
- Elbaradi, T.; Pieler, T. Zinc Finger Proteins: What We Know and What We Would like to Know. Mech. Dev. 1991, 35, 155–169. [Google Scholar] [CrossRef]
- Au-Yeung, B.B.; Deindl, S.; Hsu, L.-Y.; Palacios, E.H.; Levin, S.E.; Kuriyan, J.; Weiss, A. The Structure, Regulation, and Function of ZAP-70. Immunol. Rev. 2009, 228, 41–57. [Google Scholar] [CrossRef]
- Huber, R.G.; Fan, H.; Bond, P.J. The Structural Basis for Activation and Inhibition of ZAP-70 Kinase Domain. PLoS Comput. Biol. 2015, 11, e1004560. [Google Scholar] [CrossRef] [PubMed]
- Li, M.M.H.; Aguilar, E.G.; Michailidis, E.; Pabon, J.; Park, P.; Wu, X.; de Jong, Y.P.; Schneider, W.M.; Molina, H.; Rice, C.M.; et al. Characterization of Novel Splice Variants of Zinc Finger Antiviral Protein (ZAP). J. Virol. 2019, 93, 10–128. [Google Scholar] [CrossRef] [PubMed]
- Chou, S.-T.; Leng, Q.; Mixson, J. Zinc Finger Nucleases: Tailor-Made for Gene Therapy. Drugs Future 2012, 37, 183. [Google Scholar] [CrossRef]
- Carbery, I.D.; Ji, D.; Harrington, A.; Brown, V.; Weinstein, E.J.; Liaw, L.; Cui, X. Targeted Genome Modification in Mice Using Zinc-Finger Nucleases. Genetics 2010, 186, 451–459. [Google Scholar] [CrossRef]
- Klug, A. The Discovery of Zinc Fingers and Their Development for Practical Applications in Gene Regulation and Genome Manipulation. Q. Rev. Biophys. 2010, 43, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Carroll, D. Genome Engineering with Zinc-Finger Nucleases. Genetics 2011, 188, 773–782. [Google Scholar] [CrossRef]
- Zimmerman, K.A.; Fischer, K.P.; Joyce, M.A.; Tyrrell, D.L.J. Zinc Finger Proteins Designed to Specifically Target Duck Hepatitis B Virus Covalently Closed Circular DNA Inhibit Viral Transcription in Tissue Culture. J. Virol. 2008, 82, 8013–8021. [Google Scholar] [CrossRef]
- Weber, N.D.; Stone, D.; Sedlak, R.H.; De Silva Feelixge, H.S.; Roychoudhury, P.; Schiffer, J.T.; Aubert, M.; Jerome, K.R. AAV-Mediated Delivery of Zinc Finger Nucleases Targeting Hepatitis B Virus Inhibits Active Replication. PLoS ONE 2014, 9, e97579. [Google Scholar] [CrossRef]
- Cradick, T.J.; Keck, K.; Bradshaw, S.; Jamieson, A.C.; McCaffrey, A.P. Zinc-Finger Nucleases as a Novel Therapeutic Strategy for Targeting Hepatitis B Virus DNAs. Mol. Ther. 2010, 18, 947–954. [Google Scholar] [CrossRef]
- Chen, E.-Q.; Dai, J.; Bai, L.; Tang, H. The Efficacy of Zinc Finger Antiviral Protein against Hepatitis B Virus Transcription and Replication in Tansgenic Mouse Model. Virol. J. 2015, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Nie, H.; Cai, D.; Zhang, J.; Liu, H.; Yan, R.; Cuconati, A.; Block, T.M.; Guo, J.-T.; Guo, H. Inhibition of Hepatitis B Virus Replication by the Host Zinc Finger Antiviral Protein. PLoS Pathog. 2013, 9, e1003494. [Google Scholar] [CrossRef]
- Joung, J.K.; Sander, J.D. TALENs: A Widely Applicable Technology for Targeted Genome Editing. Nat. Rev. Mol. Cell Biol. 2013, 14, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Boch, J. TALE and TALEN Genome Editing Technologies. Gene Genome Ed. 2021, 2, 100007. [Google Scholar] [CrossRef]
- Boch, J.; Scholze, H.; Schornack, S.; Landgraf, A.; Hahn, S.; Kay, S.; Lahaye, T.; Nickstadt, A.; Bonas, U. Breaking the Code of DNA Binding Specificity of TAL-Type III Effectors. Science 2009, 326, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.C.; Zhang, L.; Xia, D.F.; Campo, J.J.; Ankoudinova, I.V.; Guschin, D.Y.; Babiarz, J.E.; Meng, X.; Hinkley, S.J.; Lam, S.C.; et al. Improved Specificity of TALE-Based Genome Editing Using an Expanded RVD Repertoire. Nat. Methods 2015, 12, 465–471. [Google Scholar] [CrossRef] [PubMed]
- González Castro, N.G.; Bjelic, J.; Malhotra, G.; Huang, C.; Alsaffar, S.H. Comparison of the Feasibility, Efficiency, and Safety of Genome Editing Technologies. Int. J. Mol. Sci. 2021, 22, 10355. [Google Scholar] [CrossRef]
- Weber, N.D.; Stone, D.; Jerome, K.R. TALENs Targeting HBV: Designer Endonuclease Therapies for Viral Infections. Mol. Ther. 2013, 21, 1819–1821. [Google Scholar] [CrossRef]
- Benjamin, R.; Berges, B.K.; Solis-Leal, A.; Igbinedion, O.; Strong, C.L.; Schiller, M.R. TALEN Gene Editing Takes Aim on HIV. Hum. Genet. 2016, 135, 1059–1070. [Google Scholar] [CrossRef]
- Noh, K.-W.; Park, J.; Kang, M.-S. Targeted Disruption of EBNA1 in EBV-Infected Cells Attenuated Cell Growth. BMB Rep. 2016, 49, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Shukla, S.; Yang, C.; Zhang, M.; Fatma, Z.; Lingamaneni, M.; Abesteh, S.; Lane, S.T.; Xiong, X.; Wang, Y.; et al. TALEN Outperforms Cas9 in Editing Heterochromatin Target Sites. Nat. Commun. 2021, 12, 606. [Google Scholar] [CrossRef] [PubMed]
- Bloom, K.; Ely, A.; Mussolino, C.; Cathomen, T.; Arbuthnot, P. Inactivation of Hepatitis B Virus Replication in Cultured Cells and In Vivo with Engineered Transcription Activator-like Effector Nucleases. Mol. Ther. 2013, 21, 1889–1897. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, W.; Lin, J.; Wang, F.; Wu, M.; Chen, C.; Zheng, Y.; Peng, X.; Li, J.; Yuan, Z. An Efficient Antiviral Strategy for Targeting Hepatitis B Virus Genome Using Transcription Activator-like Effector Nucleases. Mol. Ther. 2014, 22, 303–311. [Google Scholar] [CrossRef]
- Smith, T.; Singh, P.; Chmielewski, K.O.; Bloom, K.; Cathomen, T.; Arbuthnot, P.; Ely, A. Improved Specificity and Safety of Anti-Hepatitis B Virus TALENs Using Obligate Heterodimeric FokI Nuclease Domains. Viruses 2021, 13, 1344. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, T.; Nicholson, S.; Ely, A.; Arbuthnot, P.; Bloom, K. Improved Antiviral Efficacy Using TALEN-Mediated Homology Directed Recombination to Introduce Artificial Primary MiRNAs into DNA of Hepatitis B Virus. Biochem. Biophys. Res. Commun. 2016, 478, 1563–1568. [Google Scholar] [CrossRef]
- Allweiss, L.; Dandri, M. The Role of CccDNA in HBV Maintenance. Viruses 2017, 9, 156. [Google Scholar] [CrossRef]
- Li, F.; Cheng, L.; Murphy, C.M.; Reszka-Blanco, N.J.; Wu, Y.; Chi, L.; Hu, J.; Su, L. Minicircle HBV CccDNA with a Gaussia Luciferase Reporter for Investigating HBV CccDNA Biology and Developing CccDNA-Targeting Drugs. Sci. Rep. 2016, 6, 36483. [Google Scholar] [CrossRef]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F., III. ZFN, TALEN, and CRISPR/Cas-Based Methods for Genome Engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef]
- Li, T.; Yang, Y.; Qi, H.; Cui, W.; Zhang, L.; Fu, X.; He, X.; Liu, M.; Li, P.-F.; Yu, T. CRISPR/Cas9 Therapeutics: Progress and Prospects. Signal Transduct. Target. Ther. 2023, 8, 36. [Google Scholar] [CrossRef]
- Abdelaal, A.S.; Yazdani, S.S. Development and Use of CRISPR in Industrial Applications. In Genome Engineering via CRISPR-Cas9 System; Elsevier: Amsterdam, The Netherlands, 2020; pp. 177–197. ISBN 9780128181409. [Google Scholar]
- Sheridan, C. The World’s First CRISPR Therapy Is Approved: Who Will Receive It? Nat. Biotechnol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Akidil, E.; Albanese, M.; Buschle, A.; Ruhle, A.; Pich, D.; Keppler, O.T.; Hammerschmidt, W. Highly Efficient CRISPR-Cas9-Mediated Gene Knockout in Primary Human B Cells for Functional Genetic Studies of Epstein-Barr Virus Infection. PLoS Pathog. 2021, 17, e1009117. [Google Scholar] [CrossRef]
- Ashraf, M.U.; Salman, H.M.; Khalid, M.F.; Khan, M.H.F.; Anwar, S.; Afzal, S.; Idrees, M.; Chaudhary, S.U. CRISPR-Cas13a Mediated Targeting of Hepatitis C Virus Internal-Ribosomal Entry Site (IRES) as an Effective Antiviral Strategy. Biomed. Pharmacother. 2021, 136, 111239. [Google Scholar] [CrossRef]
- Hussein, M.; Molina, M.A.; Berkhout, B.; Herrera-Carrillo, E. A CRISPR-Cas Cure for HIV/AIDS. Int. J. Mol. Sci. 2023, 24, 1563. [Google Scholar] [CrossRef]
- Lin, S.-R.; Yang, H.-C.; Kuo, Y.-T.; Liu, C.-J.; Yang, T.-Y.; Sung, K.-C.; Lin, Y.-Y.; Wang, H.-Y.; Wang, C.-C.; Shen, Y.-C.; et al. The CRISPR/Cas9 System Facilitates Clearance of the Intrahepatic HBV Templates In Vivo. Mol. Ther. Nucleic Acids 2014, 3, e186. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Xie, K.; Xu, Y.; Wang, L.; Chen, K.; Zhang, L.; Fang, J. CRISPR/Cas9 Produces Anti-Hepatitis B Virus Effect in Hepatoma Cells and Transgenic Mouse. Virus Res. 2016, 217, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-C.; Chen, Y.-H.; Kao, J.-H.; Ching, C.; Liu, I.-J.; Wang, C.-C.; Tsai, C.-H.; Wu, F.-Y.; Liu, C.-J.; Chen, P.-J.; et al. Permanent Inactivation of HBV Genomes by CRISPR/Cas9-Mediated Non-Cleavage Base Editing. Mol. Ther. Nucleic Acids 2020, 20, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.; Feng, J.; Liu, X.; Wang, H.; Li, Q.; Li, J.; Xu, T.; Sajid, M.; Ullah, H.; Zhou, L.; et al. Inhibition of Hepatitis B Virus by AAV8-Derived CRISPR/SaCas9 Expressed from Liver-Specific Promoters. Front. Microbiol. 2021, 12, 665184. [Google Scholar] [CrossRef]
- Ramanan, V.; Shlomai, A.; Cox, D.B.T.; Schwartz, R.E.; Michailidis, E.; Bhatta, A.; Scott, D.A.; Zhang, F.; Rice, C.M.; Bhatia, S.N. CRISPR/Cas9 Cleavage of Viral DNA Efficiently Suppresses Hepatitis B Virus. Sci. Rep. 2015, 5, 10833. [Google Scholar] [CrossRef]
- Stone, D.; Long, K.R.; Loprieno, M.A.; De Silva Feelixge, H.S.; Kenkel, E.J.; Liley, R.M.; Rapp, S.; Roychoudhury, P.; Nguyen, T.; Stensland, L.; et al. CRISPR-Cas9 Gene Editing of Hepatitis B Virus in Chronically Infected Humanized Mice. Mol. Ther. Methods Clin. Dev. 2021, 20, 258–275. [Google Scholar] [CrossRef]
- Qi, Z.; Li, G.; Hu, H.; Yang, C.; Zhang, X.; Leng, Q.; Xie, Y.; Yu, D.; Zhang, X.; Gao, Y.; et al. Recombinant Covalently Closed Circular Hepatitis B Virus DNA Induces Prolonged Viral Persistence in Immunocompetent Mice. J. Virol. 2014, 88, 8045–8056. [Google Scholar] [CrossRef]
- Guo, C.; Ma, X.; Gao, F.; Guo, Y. Off-Target Effects in CRISPR/Cas9 Gene Editing. Front. Bioeng. Biotechnol. 2023, 11, 1143157. [Google Scholar] [CrossRef] [PubMed]
- Alves-Bezerra, M.; Furey, N.; Johnson, C.G.; Bissig, K.-D. Using CRISPR/Cas9 to Model Human Liver Disease. JHEP Rep. 2019, 1, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Kallimasioti-Pazi, E.M.; Thelakkad Chathoth, K.; Taylor, G.C.; Meynert, A.; Ballinger, T.; Kelder, M.J.E.; Lalevée, S.; Sanli, I.; Feil, R.; Wood, A.J. Heterochromatin Delays CRISPR-Cas9 Mutagenesis but Does Not Influence the Outcome of Mutagenic DNA Repair. PLoS Biol. 2018, 16, e2005595. [Google Scholar] [CrossRef]
- Sioud, M. RNA Interference: Story and Mechanisms. In Methods in Molecular Biology; Springer: New York, NY, USA, 2021; Volume 2282, pp. 1–15. ISBN 9781071612972. [Google Scholar]
- Zhang, X.; Goel, V.; Robbie, G.J. Pharmacokinetics of Patisiran, the First Approved RNA Interference Therapy in Patients with Hereditary Transthyretin-mediated Amyloidosis. J. Clin. Pharmacol. 2020, 60, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Setten, R.L.; Rossi, J.J.; Han, S.-P. The Current State and Future Directions of RNAi-Based Therapeutics. Nat. Rev. Drug Discov. 2019, 18, 421–446. [Google Scholar] [CrossRef] [PubMed]
- Levanova, A.; Poranen, M.M. RNA Interference as a Prospective Tool for the Control of Human Viral Infections. Front. Microbiol. 2018, 9, 2151. [Google Scholar] [CrossRef] [PubMed]
- Yuen, M.-F.; Schiefke, I.; Yoon, J.-H.; Ahn, S.H.; Heo, J.; Kim, J.H.; Chan, H.L.Y.; Yoon, K.T.; Klinker, H.; Manns, M.; et al. RNA Interference Therapy with ARC-520 Results in Prolonged Hepatitis B Surface Antigen Response in Patients with Chronic Hepatitis B Infection. Hepatology 2020, 72, 19–31. [Google Scholar] [CrossRef]
- Gane, E.; Schwabe, C.; Given, B.; Schluep, T.; Hamilton, J.; Lai, C.-L.; Locarnini, S.; Lau, J.; Ferrari, C.; Gish, R. A Phase 1 Study to Evaluate Safety and Tolerability of Escalating Single Doses of the Hepatitis B Virus RNA Interference Drug ARC-521 in a Healthy Volunteer Population. J. Hepatol. 2017, 66, S265. [Google Scholar] [CrossRef]
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X.-J. Therapeutic SiRNA: State of the Art. Signal Transduct. Target. Ther. 2020, 5, 101. [Google Scholar] [CrossRef]
- Agarwal, K.; Gane, E.; Cheng, C.; Sievert, W.; Roberts, S.K.; Ahn, S.H.; Kim, Y.J.; Streinu-Cercel, A.; Denning, J.; Symonds, W.; et al. HBcrAg, HBV-RNA Declines in A Phase 2a Study Evaluating the Multi-Dose Activity of ARB-1467 in HBeAg-Positive and Negative Virally Suppressed Patients with Hepatitis B. Available online: https://www.natap.org/2017/AASLD/HBcrAg,%20HBV-RNA%20Declines%20in%20A%20Phase%202a%20Study%20Evaluating%20the%20Multi-Dose%20Activity%20of%20ARB-1467%20in%20HBeAg-Positive%20and%20Negative%20Virally%20Suppressed%20Patients%20Wit.pdf (accessed on 4 December 2023).
- Bi-Weekly Dosing of ARB-1467 LNP SiRNA in HBeAgNegative, Virally Suppressed Patients with Chronic HBV Infection Leads to Deeper Declines in HBsAg and Potential Association with IL28b. Available online: https://www.natap.org/2017/AASLD/AASLD_64.htm (accessed on 25 October 2023).
- Mao, T.; Zhang, K.; Kloth, C.; Roelvink, P.; Suhy, D. Superior Suppression of Hepatitis B Virus DNA and Antigen Levels in a Chimeric Mouse Model When BB-103, a DNA-Directed RNA Interference Agent, Is Coupled with Standard of Care Drugs. J. Hepatol. 2017, 66, S260. [Google Scholar] [CrossRef]
- Thi, E.P.; Dhillon, A.P.; Ardzinski, A.; Bidirici-Ertekin, L.; Cobarrubias, K.D.; Cuconati, A.; Kondratowicz, A.S.; Kwak, K.; Li, A.H.L.; Miller, A.; et al. ARB-1740, a RNA Interference Therapeutic for Chronic Hepatitis B Infection. ACS Infect. Dis. 2019, 5, 725–737. [Google Scholar] [CrossRef]
- Yuen, M.-F.; Locarnini, S.; Given, B.; Schluep, T.; Hamilton, J.; Biermer, M.; Kalmeijer, R.; Beumont, M.; Lenz, O.; Cloherty, G.; et al. First Clinical Experience with RNA Interference-Based Triple Combination Therapy in Chronic Hepatitis B: JNJ-3989, JNJ-6379 and a Nucleos(t)Ide Analogue. Available online: https://ir.arrowheadpharma.com/index.php/static-files/6d7edbe7-c5a2-46c5-9990-a9cdfec854a9 (accessed on 25 October 2023).
- Wang, J.; Chen, R.; Zhang, R.; Ding, S.; Zhang, T.; Yuan, Q.; Guan, G.; Chen, X.; Zhang, T.; Zhuang, H.; et al. The GRNA-MiRNA-GRNA Ternary Cassette Combining CRISPR/Cas9 with RNAi Approach Strongly Inhibits Hepatitis B Virus Replication. Theranostics 2017, 7, 3090–3105. [Google Scholar] [CrossRef]
- Kadelka, S.; Dahari, H.; Ciupe, S.M. Understanding the Antiviral Effects of RNAi-Based Therapy in HBeAg-Positive Chronic Hepatitis B Infection. Sci. Rep. 2021, 11, 200. [Google Scholar] [CrossRef]
- Gane, E.; Lim, Y.-S.; Tangkijvanich, P.; O’Beirne, J.; Lim, T.H.; Bakardjiev, A.; Ding, X.; Connolly, L.; Huang, S.; Kim, J.; et al. Preliminary Safety and Antiviral Activity of VIR-2218, an X-Targeting HBV RNAi Therapeutic, in Chronic Hepatitis B Patients. J. Hepatol. 2020, 73, S50–S51. [Google Scholar] [CrossRef]
- Gane, E.; Locarnini, S.; Lim, T.H.; Strasser, S.; Sievert, W.; Cheng, W.; Thompson, A.; Given, B.; Schluep, T.; Hamilton, J.; et al. Short-Term Treatment with RNA Interference Therapy, JNJ-3989, Results in Sustained Hepatitis B Surface Antigen Supression in Patients with Chronic Hepatitis B Receiving Nucleos(t)Ide Analogue Treatment. J. Hepatol. 2020, 73, S20. [Google Scholar] [CrossRef]
- Yu, Y.; Koenig, A.; Livingston, C.; van Gijzel, H.; Elston, R.; Dickens, T.; Paff, M.T.; Rice, C.M.; You, S.; Schneider, W.M. Rna Launch System Enables In Vitro Evaluation of Bepirovirsen Efficacy against Hbv Variants. Available online: https://www.natap.org/2022/AASLD/AASLD_59.htm (accessed on 26 November 2023).
- You, S.; Elston, R.; Yuen, M.-F. Reply to: “Bepirovirsen/GSK3389404: Antisense or TLR9 Agonists”? J. Hepatol. 2023, 78, e108–e110. [Google Scholar] [CrossRef]
- Vaillant, A. Bepirovirsen/GSK3389404: Antisense or TLR9 Agonists? J. Hepatol. 2023, 78, e107–e108. [Google Scholar] [CrossRef]
- Yau, A.H.L.; Yoshida, E.M. Hepatitis C Drugs: The End of the Pegylated Interferon Era and the Emergence of All-Oral, Interferon-Free Antiviral Regimens: A Concise Review. Can. J. Gastroenterol. Hepatol. 2014, 28, 445–451. [Google Scholar] [CrossRef]
- Brzdęk, M.; Zarębska-Michaluk, D.; Rzymski, P.; Lorenc, B.; Kazek, A.; Tudrujek-Zdunek, M.; Janocha-Litwin, J.; Mazur, W.; Dybowska, D.; Berak, H.; et al. Changes in Characteristics of Patients with Hepatitis C Virus-Related Cirrhosis from the Beginning of the Interferon-Free Era. World J. Gastroenterol. 2023, 29, 2015–2033. [Google Scholar] [CrossRef]
- Afrin, H.; Bai, R.G.; Kumar, R.; Ahmad, S.S.; Agarwal, S.K.; Nurunnabi, M. Oral Delivery of RNAi for Cancer Therapy. Cancer Metastasis Rev. 2023, 42, 699–724. [Google Scholar] [CrossRef] [PubMed]
- Paunovska, K.; Loughrey, D.; Dahlman, J.E. Drug Delivery Systems for RNA Therapeutics. Nat. Rev. Genet. 2022, 23, 265–280. [Google Scholar] [CrossRef]
- Boden, D.; Pusch, O.; Lee, F.; Tucker, L.; Ramratnam, B. Human Immunodeficiency Virus Type 1 Escape from RNA Interference. J. Virol. 2003, 77, 11531–11535. [Google Scholar] [CrossRef] [PubMed]
- Das, A.T.; Brummelkamp, T.R.; Westerhout, E.M.; Vink, M.; Madiredjo, M.; Bernards, R.; Berkhout, B. Human Immunodeficiency Virus Type 1 Escapes from RNA Interference-Mediated Inhibition. J. Virol. 2004, 78, 2601–2605. [Google Scholar] [CrossRef]
- Gitlin, L.; Stone, J.K.; Andino, R. Poliovirus Escape from RNA Interference: Short Interfering RNA-Target Recognition and Implications for Therapeutic Approaches. J. Virol. 2005, 79, 1027–1035. [Google Scholar] [CrossRef]
- Dallas, A.; Ilves, H.; Ma, H.; Chin, D.J.; Maclachlan, I.; Klumpp, K.; Johnston, B.H. Inhibition of Hepatitis C Virus in Chimeric Mice by Short Synthetic Hairpin RNAs: Sequence Analysis of Surviving Virus Shows Added Selective Pressure of Combination Therapy. J. Virol. 2014, 88, 4647–4656. [Google Scholar] [CrossRef]
- Wilson, R.; Purcell, D.; Netter, H.J.; Revill, P.A. Does RNA Interference Provide New Hope for Control of Chronic Hepatitis B Infection? Antivir. Ther. 2009, 14, 879–889. [Google Scholar] [CrossRef]
- van den Berg, F.; Limani, S.W.; Mnyandu, N.; Maepa, M.B.; Ely, A.; Arbuthnot, P. Advances with RNAi-Based Therapy for Hepatitis B Virus Infection. Viruses 2020, 12, 851. [Google Scholar] [CrossRef]
- Liu, Y.; Maya, S.; Ploss, A. Animal Models of Hepatitis B Virus Infection–Success, Challenges, and Future Directions. Viruses 2021, 13, 777. [Google Scholar] [CrossRef]
- Hui, R.W.-H.; Mak, L.-Y.; Seto, W.-K.; Yuen, M.-F. RNA Interference as a Novel Treatment Strategy for Chronic Hepatitis B Infection. Clin. Mol. Hepatol. 2022, 28, 408–424. [Google Scholar] [CrossRef]
- Hong, J.; Martinez Montero, S.; Tan, H.; De Costa, T.; Nie, Y.; Pandey, R.; Rajwanshi, V.K.; Bhattacharya, A.; Kao, C.; Kang, H.; et al. ALG-125755, a Small Interfering RNA (SiRNA) Against Hepatitis B Virus (HBV) Effectively Inhibits Hepatitis B Surface Antigen (HBsAg) Secretion in HBV Cell Models and the AAV-HBV Mouse Model. J. Hepatol. 2021, 75, S742. [Google Scholar]
- Taylor, N.P. Aligos Culls Another Hepatitis B Asset, Citing Inconclusive Efficacy Data. Available online: https://www.fiercebiotech.com/biotech/aligos-culls-another-hepatitis-b-asset-citing-inconclusive-efficacy-data (accessed on 4 December 2023).
- Jucov, A.; Gane, E.J.; Fitzgerald, M.; Le, K.; Wang, S.; Ammar, L.; Burnett, C.; Haceatrean, A.; Gupta, K.; Clark, D.; et al. Safety, Pharmacokinetics, and Antiviral Activity of Single Ascending Doses of ALG-125755, a GalNAc-Conjugated Small Interfering RNA, in Subjects with Chronic Hepatitis B. J. Hepatol. 2023, 78, S1162–S1163. [Google Scholar] [CrossRef]
- Gane, E.; Fitzgerald, M.; Le, K.; Wang, S.; Ammar, L.; Gupta, K.; Schwabe, C.; Venkatraman, M.; Lin, T.-I.; Fry, J. Safety, Tolerability and Pharmacokinetics (PK) of Single Ascending Doses of ALG-125755, a GalNAc-Conjugated Small Interfering RNA (SiRNA), in Healthy Volunteers (HV). Available online: https://www.aligos.com/wp-content/uploads/2023/02/1_ALG-125755_APASL-2023_Clinical-Poster.pdf (accessed on 4 December 2023).
- Yuen, M.-F.; Wong, D.K.-H.; Schluep, T.; Lai, C.-L.; Ferrari, C.; Locarnini, S.; Lo, R.C.-L.; Gish, R.G.; Hamilton, J.; Wooddell, C.I.; et al. Long-Term Serological, Virological and Histological Responses to RNA Inhibition by ARC-520 in Chinese Chronic Hepatitis B Patients on Entecavir Treatment. Gut 2022, 71, 789–797. [Google Scholar] [CrossRef]
- Streinu-Cercel, A.; Gane, E.; Cheng, W.; Sievert, W.; Roberts, S.; Ahn, S.H.; Kim, Y.J.; Agarwal, K.; Niforos, D.; Symonds, B.; et al. A Phase 2a Study Evaluating the Multi-Dose Activity of ARB-1467 in HBeAg-Positive and Negative Virally Suppressed Subjects with Hepatitis B. J. Hepatol. 2017, 66, S688–S689. [Google Scholar] [CrossRef]
- Yuen, M.-F.; Heo, J.; Nahass, R.; Wong, G.; Burda, T.; Bhamidimarri, K.R.; Hu, T.-H.; Nguyen, T.; Lim, Y.-S.; Chen, C.-Y.; et al. Preliminary Safety and Antiviral Activity of AB-729 Combination Treatment with Pegylated Interferon Alfa-2a in Virally Suppressed, HBeAg-Negative Subjects with Chronic HBV Infection. Available online: https://www.arbutusbio.com/wp-content/uploads/2023/06/EASL-2023_AB-729-201-IFN-LB-poster_FINAL.pdf (accessed on 23 October 2023).
- Thi, E.P.; Yuen, M.-F.; Gane, E.; Sevinsky, H.; Sims, K.D.; Anderson, M.; Lam, A.M.; Sofia, M.J.; Cloherty, G.A.; Picchio, G. Hepatitis B Patients Correlates with Suppression of All HBsAg Isoforms and HBV RNA. Available online: https://www.postersessiononline.eu/173580348_eu/congresos/ILC2021/aula/-PO_2822_ILC2021.pdf (accessed on 4 December 2023).
- Gupta, S.V.; Fanget, M.C.; MacLauchlin, C.; Clausen, V.A.; Li, J.; Cloutier, D.; Shen, L.; Robbie, G.J.; Mogalian, E. Clinical and Preclinical Single-Dose Pharmacokinetics of VIR-2218, an RNAi Therapeutic Targeting HBV Infection. Drugs R D 2021, 21, 455–465. [Google Scholar] [CrossRef]
- Gane, E.; Yuen, M.-F.; Kakuda, T.N.; Ogawa, T.; Takahashi, Y.; Goeyvaerts, N.; Lonjon-Domanec, I.; Vaughan, T.; Schluep, T.; Hamilton, J.; et al. JNJ-73763989 Pharmacokinetics and Safety: Liver-Targeted SiRNAs against Hepatitis B Virus, in Japanese and Non-Japanese Healthy Adults, and Combined with JNJ-56136379 and a Nucleos(t)Ide Analogue in Patients with Chronic Hepatitis B. Antivir. Ther. 2022, 27, 135965352210938. [Google Scholar] [CrossRef]
- Yuen, M.-F.; Locarnini, S.; Lim, T.H.; Strasser, S.I.; Sievert, W.; Cheng, W.; Thompson, A.J.; Given, B.D.; Schluep, T.; Hamilton, J.; et al. Combination Treatments Including the Small-Interfering RNA JNJ-3989 Induce Rapid and Sometimes Prolonged Viral Responses in Patients with CHB. J. Hepatol. 2022, 77, 1287–1298. [Google Scholar] [CrossRef]
- Kakuda, T.N.; Halabi, A.; Klein, G.; Sanga, M.; Guinard-Azadian, C.; Kowalik, M.; Nedoschinsky, K.; Nangosyah, J.; Ediage, E.N.; Hillewaert, V.; et al. Pharmacokinetics of JNJ-73763989 and JNJ-56136379 (Bersacapavir) in Participants with Moderate Hepatic Impairment. J. Clin. Pharmacol. 2023, 63, 732–741. [Google Scholar] [CrossRef]
- Ye, X.; Tateno, C.; Thi, E.P.; Kakuni, M.; Snead, N.M.; Ishida, Y.; Barnard, T.R.; Sofia, M.J.; Shimada, T.; Lee, A.C.H. Hepatitis B Virus Therapeutic Agent ARB-1740 Has Inhibitory Effect on Hepatitis Delta Virus in a New Dually-Infected Humanized Mouse Model. ACS Infect. Dis. 2019, 5, 738–749. [Google Scholar] [CrossRef]
- Lewandowska, M.; Piekarska, A. New Directions in Hepatitis B Therapy Research. Clin. Exp. Hepatol. 2017, 3, 119–126. [Google Scholar] [CrossRef]
- Yuen, M.-F.; Heo, J.; Jang, J.-W.; Yoon, J.-H.; Kweon, Y.-O.; Park, S.-J.; Tami, Y.; You, S.; Yates, P.; Tao, Y.; et al. Safety, Tolerability and Antiviral Activity of the Antisense Oligonucleotide Bepirovirsen in Patients with Chronic Hepatitis B: A Phase 2 Randomized Controlled Trial. Nat. Med. 2021, 27, 1725–1734. [Google Scholar] [CrossRef]
- Yuen, M.-F.; Lim, S.-G.; Plesniak, R.; Tsuji, K.; Janssen, H.L.A.; Pojoga, C.; Gadano, A.; Popescu, C.P.; Stepanova, T.; Asselah, T.; et al. Efficacy and Safety of Bepirovirsen in Chronic Hepatitis B Infection. N. Engl. J. Med. 2022, 387, 1957–1968. [Google Scholar] [CrossRef]
- Yuen, M.-F.; Heo, J.; Kumada, H.; Suzuki, F.; Suzuki, Y.; Xie, Q.; Jia, J.; Karino, Y.; Hou, J.; Chayama, K.; et al. Phase IIa, Randomised, Double-Blind Study of GSK3389404 in Patients with Chronic Hepatitis B on Stable Nucleos(t)Ide Therapy. J. Hepatol. 2022, 77, 967–977. [Google Scholar] [CrossRef]
- Han, K.; Cremer, J.; Elston, R.; Oliver, S.; Baptiste-Brown, S.; Chen, S.; Gardiner, D.; Davies, M.; Saunders, J.; Hamatake, R.; et al. A Randomized, Double-blind, Placebo-controlled, First-time-in-human Study to Assess the Safety, Tolerability, and Pharmacokinetics of Single and Multiple Ascending Doses of GSK3389404 in Healthy Subjects. Clin. Pharmacol. Drug Dev. 2019, 8, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Gane, E.; Yuen, M.-F.; Kim, D.J.; Chan, H.L.-Y.; Surujbally, B.; Pavlovic, V.; Das, S.; Triyatni, M.; Kazma, R.; Grippo, J.F.; et al. Clinical Study of Single-stranded Oligonucleotide RO7062931 in Healthy Volunteers and Patients with Chronic Hepatitis B. Hepatology 2021, 74, 1795–1808. [Google Scholar] [CrossRef] [PubMed]
- Gane, E.J.; Kim, W.; Lim, T.H.; Tangkijvanich, P.; Yoon, J.-H.; Sievert, W.; Sukeepaisarnjaroen, W.; Thompson, A.J.; Pavlovic, V.; Surujbally, B.; et al. First-in-Human Randomized Study of RNAi Therapeutic RG6346 for Chronic Hepatitis B Virus Infection. J. Hepatol. 2023, 79, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Bazinet, M.; Pântea, V.; Cebotarescu, V.; Cojuhari, L.; Jimbei, P.; Albrecht, J.; Schmid, P.; Le Gal, F.; Gordien, E.; Krawczyk, A.; et al. Safety and Efficacy of REP 2139 and Pegylated Interferon Alfa-2a for Treatment-Naive Patients with Chronic Hepatitis B Virus and Hepatitis D Virus Co-Infection (REP 301 and REP 301-LTF): A Non-Randomised, Open-Label, Phase 2 Trial. Lancet Gastroenterol. Hepatol. 2017, 2, 877–889. [Google Scholar] [CrossRef] [PubMed]
- Vaillant, A. REP 2139: Antiviral Mechanisms and Applications in Achieving Functional Control of HBV and HDV Infection. ACS Infect. Dis. 2019, 5, 675–687. [Google Scholar] [CrossRef]
- Bazinet, M.; Pântea, V.; Placinta, G.; Moscalu, I.; Cebotarescu, V.; Cojuhari, L.; Jimbei, P.; Iarovoi, L.; Smesnoi, V.; Musteata, T.; et al. Safety and Efficacy of 48 Weeks REP 2139 or REP 2165, Tenofovir Disoproxil, and Pegylated Interferon Alfa-2a in Patients with Chronic HBV Infection Naïve to Nucleos(t)Ide Therapy. Gastroenterology 2020, 158, 2180–2194. [Google Scholar] [CrossRef]
- Bazinet, M.; Pântea, V.; Cebotarescu, V.; Cojuhari, L.; Jimbei, P.; Anderson, M.; Gersch, J.; Holzmayer, V.; Elsner, C.; Krawczyk, A.; et al. Persistent Control of Hepatitis B Virus and Hepatitis Delta Virus Infection Following REP 2139-Ca and Pegylated Interferon Therapy in Chronic Hepatitis B Virus/Hepatitis Delta Virus Coinfection. Hepatol. Commun. 2021, 5, 189–202. [Google Scholar] [CrossRef]
- Bazinet, M.; Pântea, V.; Placinta, G.; Moscalu, I.; Cebotarescu, V.; Cojuhari, L.; Jimbei, P.; Iarovoi, L.; Smesnoi, V.; Musteata, T.; et al. Benefit of Transaminase Elevations in Establishing Functional Cure of HBV Infection during Nap-Based Combination Therapy. J. Viral Hepat. 2021, 28, 817–825. [Google Scholar] [CrossRef]
- Bazinet, M.; Anderson, M.; Pântea, V.; Placinta, G.; Moscalu, I.; Cebotarescu, V.; Cojuhari, L.; Jimbei, P.; Iarovoi, L.; Smesnoi, V.; et al. Analysis of HBsAg Immunocomplexes and CccDNA Activity during and Persisting after NAP-Based Therapy. Hepatol. Commun. 2021, 5, 1873–1887. [Google Scholar] [CrossRef] [PubMed]
- Bazinet, M.; Anderson, M.; Pântea, V.; Placinta, G.; Moscalu, I.; Cebotarescu, V.; Cojuhari, L.; Jimbei, P.; Iarovoi, L.; Smesnoi, V.; et al. HBsAg Isoform Dynamics during NAP-Based Therapy of HBeAg-Negative Chronic HBV and HBV/HDV Infection. Hepatol. Commun. 2022, 6, 1870–1880. [Google Scholar] [CrossRef] [PubMed]
- Real, C.I.; Werner, M.; Paul, A.; Gerken, G.; Schlaak, J.F.; Vaillant, A.; Broering, R. Nucleic Acid-Based Polymers Effective against Hepatitis B Virus Infection in Patients Don’t Harbor Immunostimulatory Properties in Primary Isolated Liver Cells. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hershkovich, L.; Shekhtman, L.; Bazinet, M.; Pântea, V.; Placinta, G.; Cotler, S.J.; Vaillant, A.; Dahari, H. Rapid Monophasic HBsAg Decline during Nucleic-Acid Polymer–Based Therapy Predicts Functional Cure. Hepatol. Commun. 2023, 7, e0205. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, S.; Tam, D.; Dick, R.; Appleby, T.; Birkus, G.; Willkom, M.; Delaney, W.E.; Notte, G.T.; Feierbach, B. Antiviral Activity of GS-5801, a Liver-Targeted Prodrug of a Lysine Demethylase 5 Inhibitor, in a Hepatitis B Virus Primary Human Hepatocyte Infection Model. J. Hepatol. 2017, 66, S690–S691. [Google Scholar] [CrossRef]
- Yu, H.-B.; Jiang, H.; Cheng, S.-T.; Hu, Z.-W.; Ren, J.-H.; Chen, J. AGK2, A SIRT2 Inhibitor, Inhibits Hepatitis B Virus Replication In Vitro and In Vivo. Int. J. Med. Sci. 2018, 15, 1356–1364. [Google Scholar] [CrossRef]
- Cheng, S.-T.; Hu, J.-L.; Ren, J.-H.; Yu, H.-B.; Zhong, S.; Wong, V.K.W.; Law, B.Y.K.; Chen, W.-X.; Xu, H.-M.; Zhang, Z.-Z.; et al. Dicoumarol, an NQO1 Inhibitor, Blocks CccDNA Transcription by Promoting Degradation of HBx. J. Hepatol. 2021, 74, 522–534. [Google Scholar] [CrossRef]
- Singh, P.; Kairuz, D.; Arbuthnot, P.; Bloom, K. Silencing Hepatitis B Virus Covalently Closed Circular DNA: The Potential of an Epigenetic Therapy Approach. World J. Gastroenterol. 2021, 27, 3182–3207. [Google Scholar] [CrossRef]
- Luo, W.; Wang, J.; Xu, D.; Bai, H.; Zhang, Y.; Zhang, Y.; Li, X. Engineered Zinc-Finger Transcription Factors Inhibit the Replication and Transcription of HBV In Vitro and In Vivo. Int. J. Mol. Med. 2018, 41, 2169–2176. [Google Scholar] [CrossRef]
- Karimova, M.; Beschorner, N.; Dammermann, W.; Chemnitz, J.; Indenbirken, D.; Bockmann, J.-H.; Grundhoff, A.; Lüth, S.; Buchholz, F.; Wiesch, J.S.Z.; et al. CRISPR/Cas9 Nickase-Mediated Disruption of Hepatitis B Virus Open Reading Frame S and X. Sci. Rep. 2015, 5, 13734. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sheng, C.; Wang, S.; Yang, L.; Liang, Y.; Huang, Y.; Liu, H.; Li, P.; Yang, C.; Yang, X.; et al. Removal of Integrated Hepatitis B Virus DNA Using CRISPR-Cas9. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Rawal, P.; Tripathi, D.M.; Hemati, H.; Kumar, J.; Sarin, S.K.; Nain, V.; Kaur, S. Targeted HBx Gene Editing by CRISPR/Cas9 System Effectively Reduces Epithelial to Mesenchymal Transition and HBV Replication in Hepatoma Cells. Res. Sq. 2022. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasianchuk, N.; Dobrowolska, K.; Harkava, S.; Bretcan, A.; Zarębska-Michaluk, D.; Jaroszewicz, J.; Flisiak, R.; Rzymski, P. Gene-Editing and RNA Interference in Treating Hepatitis B: A Review. Viruses 2023, 15, 2395. https://doi.org/10.3390/v15122395
Kasianchuk N, Dobrowolska K, Harkava S, Bretcan A, Zarębska-Michaluk D, Jaroszewicz J, Flisiak R, Rzymski P. Gene-Editing and RNA Interference in Treating Hepatitis B: A Review. Viruses. 2023; 15(12):2395. https://doi.org/10.3390/v15122395
Chicago/Turabian StyleKasianchuk, Nadiia, Krystyna Dobrowolska, Sofiia Harkava, Andreea Bretcan, Dorota Zarębska-Michaluk, Jerzy Jaroszewicz, Robert Flisiak, and Piotr Rzymski. 2023. "Gene-Editing and RNA Interference in Treating Hepatitis B: A Review" Viruses 15, no. 12: 2395. https://doi.org/10.3390/v15122395