
Dengue Types 1 and 3 Identified in Travelers Returning from Kathmandu, Nepal, during the October 2022 Outbreak Are Related to Strains Recently Identified in India
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Patients
2.2. DENV Detection
2.3. Sequencing and Bioinformatics Analysis
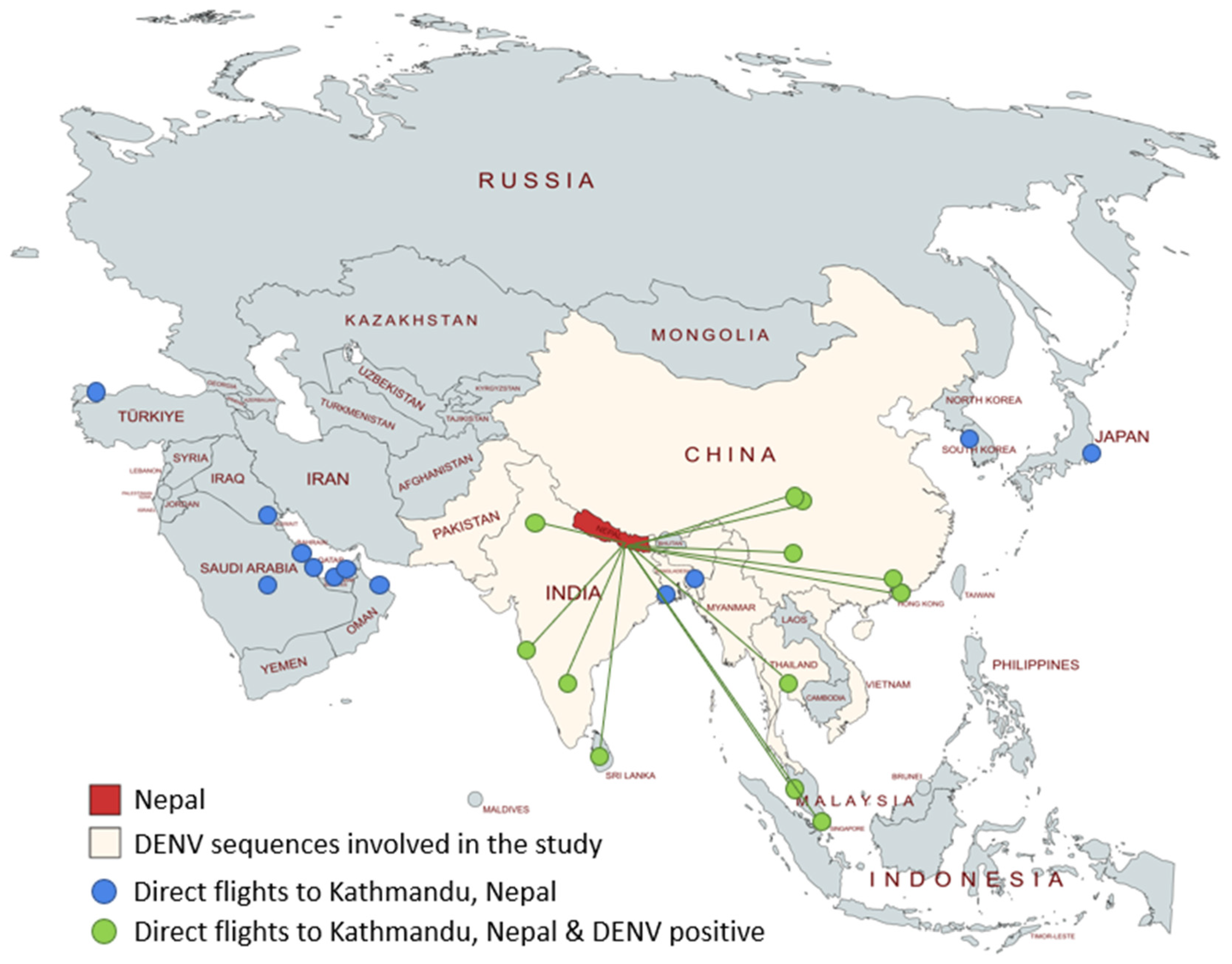
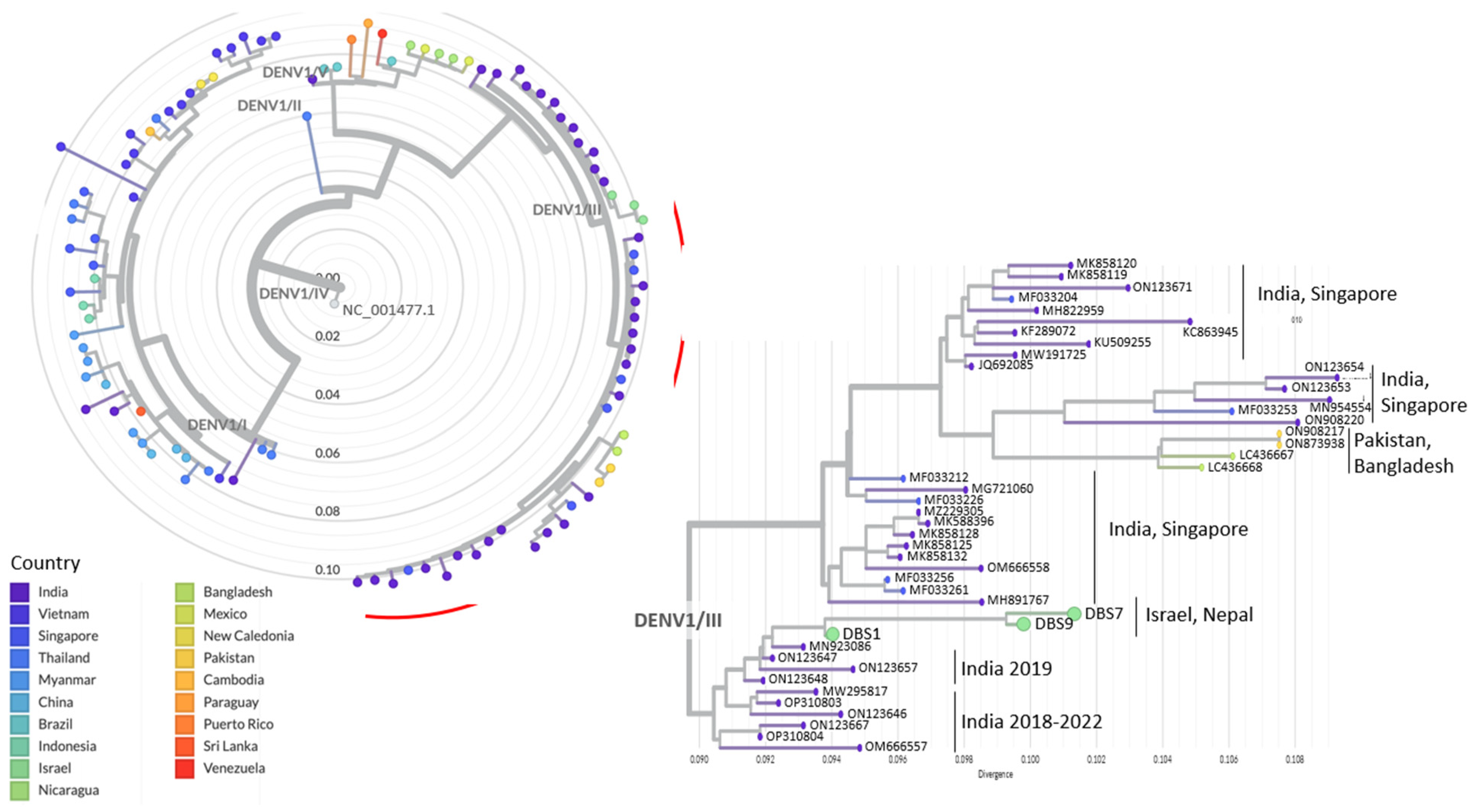
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Correction Statement
References
- Guzman, M.G.; Halstead, S.B.; Artsob, H.; Buchy, P.; Farrar, J.; Gubler, D.J.; Hunsperger, E.; Kroeger, A.; Margolis, H.S.; Martínez, E.; et al. Dengue: A continuing global threat. Nat. Rev. Microbiol. 2010, 8, S7–S16. [Google Scholar] [CrossRef]
- Kularatne, S.A.M. Dengue fever. BMJ 2015, 351, h4661. [Google Scholar] [CrossRef]
- Liu, B.; Li, N.L.; Wang, J.; Shi, P.-Y.; Wang, T.; Miller, M.A.; Li, K. Overlapping and Distinct Molecular Determinants Dictating the Antiviral Activities of TRIM56 against Flaviviruses and Coronavirus. J. Virol. 2014, 88, 13821–13835. [Google Scholar] [CrossRef]
- Lee, M.F.; Wu, Y.S.; Poh, C.L. Molecular Mechanisms of Antiviral Agents against Dengue Virus. Viruses 2023, 15, 705. [Google Scholar] [CrossRef]
- Tayal, A.; Kabra, S.K.; Lodha, R. Management of Dengue: An Updated Review. Indian J. Pediatr. 2023, 90, 168–177. [Google Scholar] [CrossRef]
- Gupta, A.; Rijhwani, P.; Pahadia, M.R.; Kalia, A.; Choudhary, S.; Bansal, D.P.; Gupta, D.; Agarwal, P.; Jat, R.K. Prevalence of Dengue Serotypes and Its Correlation With the Laboratory Profile at a Tertiary Care Hospital in Northwestern India. Cureus 2021, 13, e15029. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Nakase, T.; Giovanetti, M.; Obolski, U.; Lourenço, J. Global transmission suitability maps for dengue virus transmitted by Aedes aegypti from 1981 to 2019. Sci. Data 2023, 10, 275. [Google Scholar] [CrossRef] [PubMed]
- Mutheneni, S.R.; Morse, A.P.; Caminade, C.; Upadhyayula, S.M. Dengue burden in India: Recent trends and importance of climatic parameters. Emerg. Microbes Infect. 2017, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.G.; Reiner, R.C., Jr.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Buchs, A.; Conde, A.; Frank, A.; Gottet, C.; Hedrich, N.; Lovey, T.; Shindleman, H.; Schlagenhauf, P. The threat of dengue in Europe. New Microbes New Infect. 2022, 49–50, 101061. [Google Scholar] [CrossRef]
- Roche, B.; Léger, L.; L’ambert, G.; Lacour, G.; Foussadier, R.; Besnard, G.; Barré-Cardi, H.; Simard, F.; Fontenille, D. The Spread of Aedes albopictus in Metropolitan France: Contribution of Environmental Drivers and Human Activities and Predictions for a Near Future. PLoS ONE 2015, 10, e0125600. [Google Scholar] [CrossRef]
- La Ruche, G.; Souarès, Y.; Armengaud, A.; Peloux-Petiot, F.; Delaunay, P.; Desprès, P.; Lenglet, A.; Jourdain, F.; Leparc-Goffart, I.; Charlet, F.; et al. First two autochthonous dengue virus infections in metropolitan France, September 2010. Eurosurveillance 2010, 15, 19676. [Google Scholar] [CrossRef]
- ECDC. Autochthonous Vectorial Transmission of Dengue Virus in Mainland EU/EEA, 2010–Present. Available online: https://www.ecdc.europa.eu/en/all-topics-z/dengue/surveillance-and-disease-data/autochthonous-transmission-dengue-virus-eueea (accessed on 21 September 2023).
- Pandey, B.D.; Rai, S.K.; Morita, K.; Kurane, I. First case of Dengue virus infection in Nepal. Nepal. Med. Coll. J. 2004, 6, 157–159. [Google Scholar] [PubMed]
- Dhimal, M.; Gautam, I.; Joshi, H.D.; O’hara, R.B.; Ahrens, B.; Kuch, U. Risk Factors for the Presence of Chikungunya and Dengue Vectors (Aedes aegypti and Aedes albopictus), Their Altitudinal Distribution and Climatic Determinants of Their Abundance in Central Nepal. PLoS Neglected Trop. Dis. 2015, 9, e0003545. [Google Scholar] [CrossRef] [PubMed]
- Pandey, B.D.; Pandey, K.; Dumre, S.P.; Morita, K.; Costello, A. Struggling with a new dengue epidemic in Nepal. Lancet Infect. Dis. 2023, 23, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Bagcchi, S. Nepal faces an outbreak of dengue. Lancet Infect. Dis. 2023, 23, 35. [Google Scholar] [CrossRef] [PubMed]
- Lachish, T.; Lustig, Y.; Leshem, E.; Katz-Likvornik, S.; Biber, A.; Nadir, E.; Schwartz, E. High incidence of dengue in Israel travelers to Kathmandu, Nepal, in 2019. J. Travel Med. 2020, 27, taz105. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Health and Population Department of Health Services Epidemiology and Disease Control Division. Situation Update of Dengue 2022 (As of 31 December 2022). Available online: https://edcd.ekbana.info/news/download/situation-updates-of-dengue-as-of-30-nov-2022 (accessed on 12 July 2023).
- WHO. Dengue-Nepal. Available online: https://www.who.int/emergencies/disease-outbreak-news/item/2022-DON412 (accessed on 12 July 2023).
- Gyawali, N.; Johnson, B.J.; Dixit, S.M.; Devine, G.J. Patterns of dengue in Nepal from 2010–2019 in relation to elevation and climate. Trans. R. Soc. Trop. Med. Hyg. 2021, 115, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Amatya, B.; Schwartz, E.; Biber, A.; Erster, O.; Lustig, Y.; Pradhan, R.; Khadka, B.; Pandey, P. Dengue serotype characterization during the 2022 dengue epidemic in Kathmandu, Nepal. J. Travel Med. 2023, 30, taad034. [Google Scholar] [CrossRef]
- Santiago, G.A.; Vergne, E.; Quiles, Y.; Cosme, J.; Vazquez, J.; Medina, J.F.; Medina, F.; Colón, C.; Margolis, H.; Muñoz-Jordán, J.L. Analytical and Clinical Performance of the CDC Real Time RT-PCR Assay for Detection and Typing of Dengue Virus. PLoS Neglected Trop. Dis. 2013, 7, e2311. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, J.; Megill, C.; Bell, S.M.; Huddleston, J.; Potter, B.; Callender, C.; Sagulenko, P.; Bedford, T.; Neher, R.A. Nextstrain: Real-time tracking of pathogen evolution. Bioinformatics 2018, 34, 4121–4123. [Google Scholar] [CrossRef]
- Kazutaka, K.; Misakwa, K.; Kei-ichi, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Boratyn, G.M.; Camacho, C.; Cooper, P.S.; Coulouris, G.; Fong, A.; Ma, N.; Madden, T.L.; Matten, W.T.; McGinnis, S.D.; Merezhuk, Y.; et al. BLAST: A more efficient report with usability improvements. Nucleic Acids Res. 2013, 41, W29–W33. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name | Ct Value | Traveler/Local | % Coverage | Mean Depth | |
|---|---|---|---|---|---|
| DENV1 | DBS9 | 24.5 | local | 99.5 | 3647 |
| DBS1 | 16.08 | traveler | 85 | 62.9 | |
| DBS7 | 22.82 | traveler | 83 | 48 | |
| DENV3 | DBS4 | 22.03 | local | 94.1 | 77.2 |
| DBS38 | 23.65 | traveler | 99.6 | 565.6 | |
| DBS47 | 23.3 | traveler | 99.5 | 514.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuckerman, N.S.; Schwartz, E.; Pandey, P.; Erster, O.; Halpern, O.; Bucris, E.; Morad-Eliyahu, H.; Wax, M.; Lustig, Y. Dengue Types 1 and 3 Identified in Travelers Returning from Kathmandu, Nepal, during the October 2022 Outbreak Are Related to Strains Recently Identified in India. Viruses 2023, 15, 2334. https://doi.org/10.3390/v15122334
Zuckerman NS, Schwartz E, Pandey P, Erster O, Halpern O, Bucris E, Morad-Eliyahu H, Wax M, Lustig Y. Dengue Types 1 and 3 Identified in Travelers Returning from Kathmandu, Nepal, during the October 2022 Outbreak Are Related to Strains Recently Identified in India. Viruses. 2023; 15(12):2334. https://doi.org/10.3390/v15122334
Chicago/Turabian StyleZuckerman, Neta S., Eli Schwartz, Prativa Pandey, Oran Erster, Osnat Halpern, Efrat Bucris, Hagar Morad-Eliyahu, Marina Wax, and Yaniv Lustig. 2023. "Dengue Types 1 and 3 Identified in Travelers Returning from Kathmandu, Nepal, during the October 2022 Outbreak Are Related to Strains Recently Identified in India" Viruses 15, no. 12: 2334. https://doi.org/10.3390/v15122334