
Experimental Infection of Chickens with H5N8 High Pathogenicity Avian Influenza Viruses Isolated in Japan in the Winter of 2020–2021
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Animal Experiments
2.3. Pathogenicity
2.4. Infectivity and Virulence
2.5. Transmissibility
2.6. Statistical Analysis
3. Results
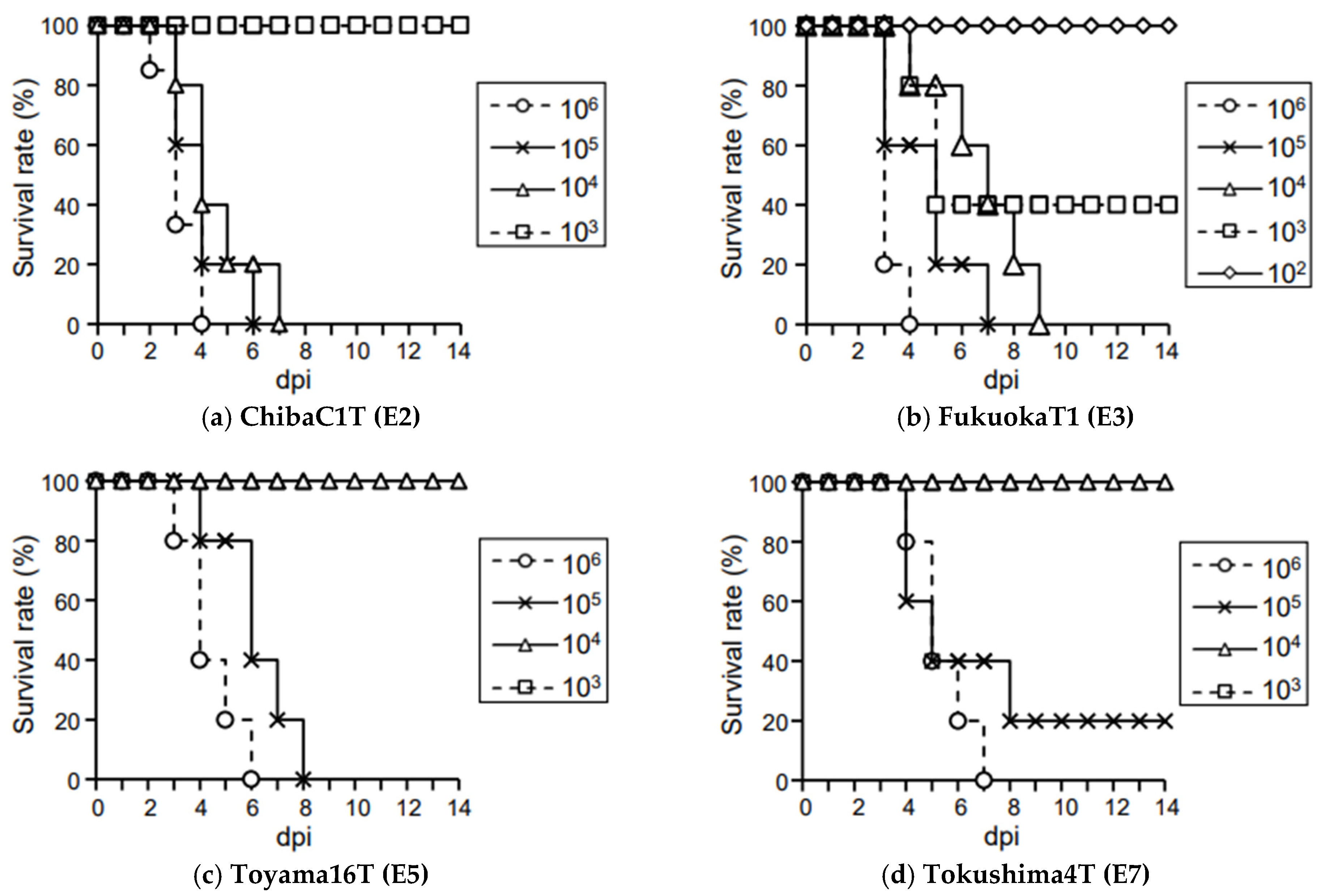
3.1. Virological Characteristics of Japanese H5N8 HPAIVs in Chickens
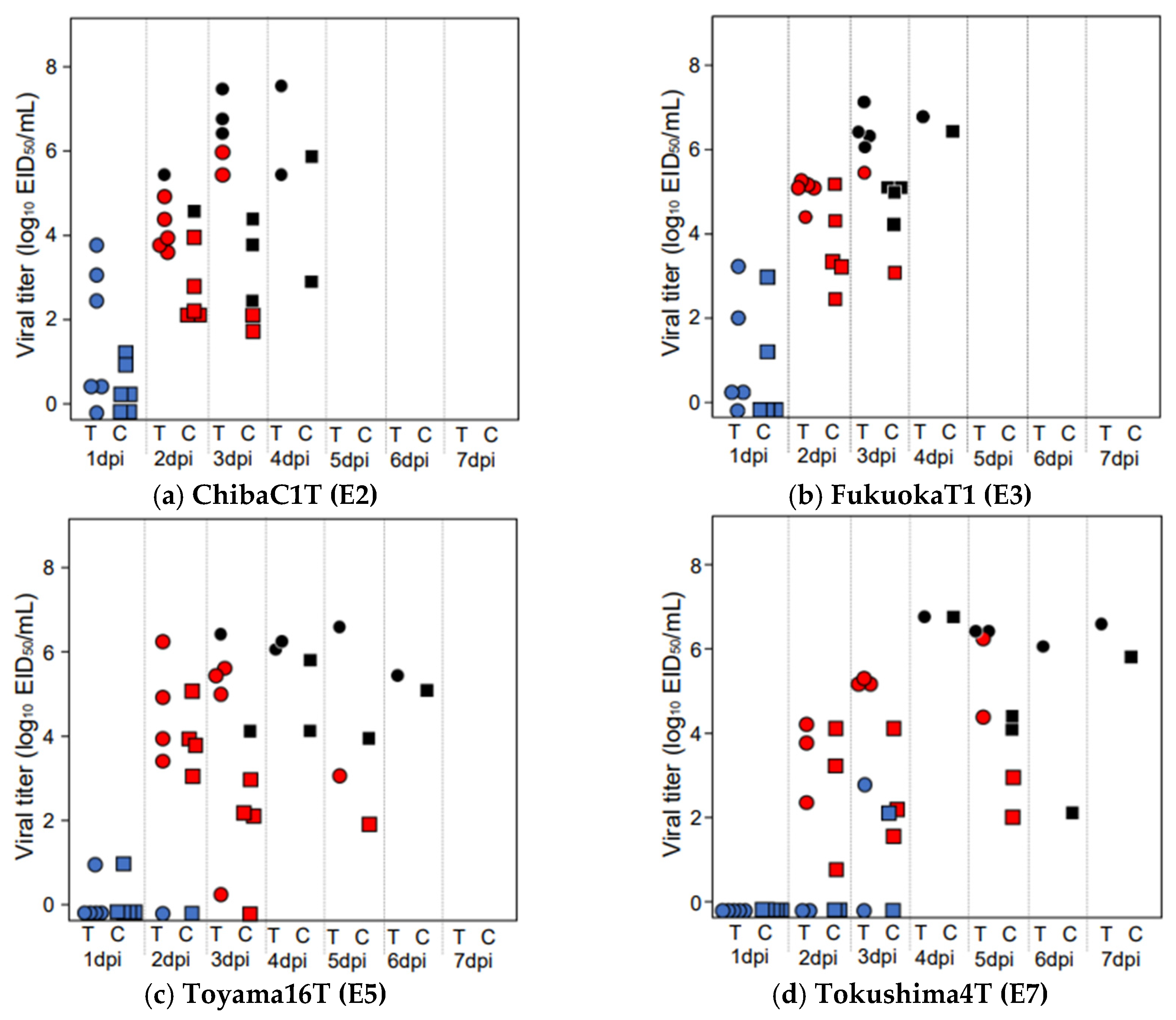
3.2. Virus Shedding in Chickens Infected with HPAIVs at a Dose of 106 EID50
3.3. Transmissibility of Japanese H5N8 Strains in Chickens
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, X.Y.; Subbarao, K.; Cox, N.J.; Guo, Y.J. Genetic characterization of the pathogenic influenza A/Goose/Guangdong/1/96 (H5N1) virus: Similarity of its hemagglutinin gene to those of H5N1 viruses from the 1997 outbreaks in Hong Kong. Virology 1999, 261, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Criado, M.F.; Swayne, D.E. Pathobiological origins and evolutionary history of highly pathogenic avian influenza viruses. Cold Spring Harb. Perspect. Med. 2021, 11, a038679. [Google Scholar] [CrossRef] [PubMed]
- Lycett, S.J.; Duchatel, F.; Digard, P. A brief history of bird flu. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180257. [Google Scholar] [CrossRef] [PubMed]
- Group WOFHNEW. Toward a unified nomenclature system for highly pathogenic avian influenza virus (H5N1). Emerg. Infect. Dis. 2008, 14, e1. [Google Scholar] [CrossRef] [PubMed]
- World Health Organizatio; World Organisation for Animal Health; Food and Agriculture Organization (WHO/OIE/FAO); H5N1 Evolution Working Group. Revised and updated nomenclature for highly pathogenic avian influenza A (H5N1) viruses. Influ. Other Respir. Viruses 2014, 8, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.J.; Donis, R.O.; World Health Organization; World Organisation for Animal Health; Food and Agriculture Organization (WHO/OIE/FAO); H5 Evolution Working Group. Nomenclature updates resulting from the evolution of avian influenza A(H5) virus clades 2.1.3.2a, 2.2.1, and 2.3.4 during 2013–2014. Influ. Other Respir. Viruses 2015, 9, 271–276. [Google Scholar] [CrossRef]
- World Health Organization. Genetic and Antigenic Characteristics of Zoonotic Influenza a Viruses and Development of Candidate Vaccine Viruses for Pandemic Preparedness. 2023. Available online: https://cdn.who.int/media/docs/default-source/influenza/who-influenza-recommendations/vcm-northern-hemisphere-recommendation-2023-2024/20230224_zoonotic_recommendations.pdf?sfvrsn=38c739fa_4 (accessed on 4 April 2023).
- Antigua, K.J.C.; Choi, W.S.; Baek, Y.H.; Song, M.S. The emergence and decennary distribution of Clade 2.3.4.4 HPAI H5Nx. Microorganisms 2019, 7, 156. [Google Scholar] [CrossRef]
- Lee, D.H.; Bertran, K.; Kwon, J.H.; Swayne, D.E. Evolution, global spread, and pathogenicity of highly pathogenic avian influenza H5Nx clade 2.3.4.4. J. Vet. Sci. 2017, 18, 269–280. [Google Scholar] [CrossRef]
- World Health Organization. Antigenic and Genetic Characteristics of Zoonotic Influenza a Viruses and Development of Candidate Vaccine Viruses for Pandemic Preparedness. 2021. Available online: https://cdn.who.int/media/docs/default-source/influenza/who-influenza-recommendations/vcm-northern-hemisphere-recommendation-2021-2022/202103_zoonotic_vaccinevirusupdate.pdf?sfvrsn=97ae1340_7 (accessed on 28 April 2023).
- Baek, Y.G.; Lee, Y.N.; Lee, D.H.; Shin, J.I.; Lee, J.H.; Chung, D.H.; Lee, E.K.; Heo, G.B.; Sagong, M.; Kye, S.J.; et al. Multiple reassortants of H5N8 Clade 2.3.4.4b Highly Pathogenic Avian Influenza Viruses Detected in South Korea during the Winter of 2020–2021. Viruses 2021, 13, 490. [Google Scholar] [CrossRef]
- Sakuma, S.; Uchida, Y.; Kajita, M.; Tanikawa, T.; Mine, J.; Tsunekuni, R.; Saito, T. First outbreak of an H5N8 highly pathogenic avian influenza virus on a chicken farm in Japan in 2020. Viruses 2021, 13, 489. [Google Scholar] [CrossRef]
- Mine, J.; Tsunekuni, R.; Tanikawa, T.; Uchida, Y.; Dubovitskiy, N.; Derko, A.; Sobolev, I.; Shestopalov, A.; Sharshov, K.; Saito, T. Genetics of Japanese H5N8 high pathogenicity avian influenza viruses isolated in winter 2020–2021 and their genetic relationship with avian influenza viruses in Siberia. Transbound. Emerg. Dis. 2022, 69, e2195–e2213. [Google Scholar] [CrossRef] [PubMed]
- Isoda, N.; Twabela, A.T.; Bazarragchaa, E.; Ogasawara, K.; Hayashi, H.; Wang, Z.J.; Kobayashi, D.; Watanabe, Y.; Saito, K.; Kida, H.; et al. Re-Invasion of H5N8 High Pathogenicity Avian influenza virus Clade 2.3.4.4b in Hokkaido, Japan, 2020. Viruses 2020, 12, 1439. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.M.; Fujimoto, Y.; Kojima, I.; Esaki, M.; Ri, K.; Masatani, T.; Matsui, T.; Ozawa, M. Genetic characterization of H5N8 highly pathogenic avian influenza viruses isolated from falcated ducks and environmental water in Japan in November 2020. Pathogens 2021, 10, 171. [Google Scholar] [CrossRef] [PubMed]
- Okuya, K.; Khalil, A.M.; Esaki, M.; Nishi, N.; Koyamada, D.; Saito, R.; Tokorozaki, K.; Hasegawa, T.; Ozawa, M. Newly emerged genotypes of highly pathogenic H5N8 avian influenza viruses in Kagoshima Prefecture, Japan during winter 2020/21. J. Gen. Virol. 2023, 104, 001870. [Google Scholar] [CrossRef] [PubMed]
- World Organization for Animal Health. Avian Influenza (Including Infection with High Pathogenicity Avian Influenza Viruses). Available online: https://www.woah.org/fileadmin/Home/fr/Health_standards/tahm/3.03.04_AI.pdf (accessed on 28 August 2023).
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- van den Brand, J.M.A.; Verhagen, J.H.; Veldhuis Kroeze, E.J.B.; van de Bildt, M.W.G.; Bodewes, R.; Herfst, S.; Richard, M.; Lexmond, P.; Bestebroer, T.M.; Fouchier, R.A.M.; et al. Wild ducks excrete highly pathogenic avian influenza virus H5N8 (2014–2015) without clinical or pathological evidence of disease. Emerg. Microbes Infect. 2018, 7, 67. [Google Scholar] [CrossRef] [PubMed]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transpl. 2013, 48, 452–458. [Google Scholar] [CrossRef]
- Brown, J.D.; Goekjian, G.; Poulson, R.; Valeika, S.; Stallknecht, D.E. Avian influenza virus in water: Infectivity is dependent on pH, salinity and temperature. Vet. Microbiol. 2009, 136, 20–26. [Google Scholar] [CrossRef]
- Brown, J.; Stallknecht, D.; Lebarbenchon, C.; Swayne, D. Survivability of Eurasian H5N1 highly pathogenic avian influenza viruses in water varies between strains. Avian Dis. 2014, 58, 453–457. [Google Scholar] [CrossRef]
- Olsen, B.; Munster, V.J.; Wallensten, A.; Waldenström, J.; Osterhaus, A.D.; Fouchier, R.A. Global patterns of influenza A virus in wild birds. Science 2006, 312, 384–388. [Google Scholar] [CrossRef]
- Global Consortium for H5N8 and Related Influenza Viruses. Role for migratory wild birds in the global spread of avian influenza H5N8. Science 2016, 354, 213–217. [Google Scholar] [CrossRef]
- Verhagen, J.H.; Fouchier, R.A.M.; Lewis, N. Highly pathogenic avian influenza viruses at the wild-domestic bird interface in Europe: Future directions for research and surveillance. Viruses 2021, 13, 212. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, J.H.; Herfst, S.; Fouchier, R.A.M. Infectious disease. How a virus travels the world. Science 2015, 347, 616–617. [Google Scholar] [CrossRef] [PubMed]
- Beerens, N.; Germeraad, E.A.; Venema, S.; Verheij, E.; Pritz-Verschuren, S.B.E.; Gonzales, J.L. Comparative pathogenicity and environmental transmission of recent highly pathogenic avian influenza H5 viruses. Emerg. Microbes Infect. 2021, 10, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.M.; Lee, E.K.; Song, B.M.; Jeong, J.; Choi, J.G.; Jeong, J.; Moon, O.K.; Yoon, H.; Cho, Y.; Kang, Y.M.; et al. Novel reassortant influenza A(H5N8) viruses among inoculated domestic and wild ducks, South Korea, 2014. Emerg. Infect. Dis. 2015, 21, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Tanikawa, T.; Sakuma, S.; Yoshida, E.; Tsunekuni, R.; Nakayama, M.; Kobayashi, S. Comparative susceptibility of the common teal (Anas crecca) to infection with high pathogenic avian influenza virus strains isolated in Japan in 2004–2017. Vet. Microbiol. 2021, 263, 109266. [Google Scholar] [CrossRef] [PubMed]
- Tanikawa, T.; Fujii, K.; Sugie, Y.; Tsunekuni, R.; Nakayama, M.; Kobayashi, S. Comparative susceptibility of mallard (Anas platyrhynchos) to infection with high pathogenicity avian influenza virus strains (Gs/Gd lineage) isolated in Japan in 2004–2017. Vet. Microbiol. 2022, 272, 109496. [Google Scholar] [CrossRef] [PubMed]
- Hiono, T.; Okamatsu, M.; Yamamoto, N.; Ogasawara, K.; Endo, M.; Kuribayashi, S.; Shichinohe, S.; Motohashi, Y.; Chu, D.H.; Suzuki, M.; et al. Experimental infection of highly and low pathogenic avian influenza viruses to chicken’s, ducks, tree sparrows, jungle crows, and black rats for the evaluation of their roles in virus transmission. Vet. Microbiol. 2016, 182, 108–115. [Google Scholar] [CrossRef]
- Keawcharoen, J.; van Riel, D.; van Amerongen, G.; Bestebroer, T.; Beyer, W.E.; van Lavieren, R.; Osterhaus, A.D.M.E.; Fouchier, R.A.M.; Kuiken, T. Wild ducks as long-distance vectors of highly pathogenic avian influenza virus (H5N1). Emerg. Infect. Dis. 2008, 14, 600–607. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Hiraoka, E.; Fujita, M.; Hijikata, N.; Ueta, M.; Takagi, K.; Konno, S.; Okuyama, M.; Watanabe, Y.; Osa, Y.; et al. Spring migration routes of mallards (Anas platyrhynchos) that winter in Japan, determined from satellite telemetry. Zool. Sci. 2008, 25, 875–881. [Google Scholar] [CrossRef]
- Ministry of the Environment of Japan. Information on Migratory Birds. Available online: https://www.env.go.jp/nature/dobutsu/bird_flu/migratory/index.html (accessed on 14 June 2023).



{kind=link}
{kind=link}
| Virus Strain | Genotype | Outbreaks in Japan in 2020–2021 a | |||||
|---|---|---|---|---|---|---|---|
| Bird Type | Number of Occurrences | Number of Prefectures Where Outbreaks Occurred | Date of Occurrence | ||||
| First | Last | Period (Days) | |||||
| Kagawa11C | E1 | Poultry | 11 | 2 | 2020.11.04 | 2020.12.01 | 27 |
| Wild birds/ environment | 4 | 4 | 2020.10.24 | 2020.12.23 | 60 | ||
| ChibaC1T | E2 | Poultry | 12 | 3 | 2021.01.20 | 2021.02.14 | 25 |
| Wild birds/ environment | 4 | 3 | 2021.01.16 | 2021.02.14 | 29 | ||
| FukuokaT1 | E3 | Poultry | 23 | 13 | 2020.11.24 | 2021.03.13 | 109 |
| Wild birds/ environment | 2 | 2 | 2020.12.22 | 2021.02.24 | 64 | ||
| Toyama16T | E5 | Poultry | 1 | 1 | 2021.02.24 | - | - |
| Wild birds/ environment | 1 | 1 | 2021.02.10 | - | - | ||
| Tokushima4T | E7 | Poultry | 5 | 4 | 2020.12.18 | 2021.02.01 | 45 |
| Wild birds/ environment | 5 | 2 | 2021.02.08 | 2021.03.03 | 23 | ||
| Virus Strain | Genotype | CLD50 a (log10EID50) | Chickens Inoculated with HPAIVs at a Dose of 106 EID50 | |||||
|---|---|---|---|---|---|---|---|---|
| MDT (dpi) b | Viral Shedding of Chickens Inoculated with HPAIVs at a Dose of 106 EID50 | |||||||
| Swab | Shedding Period (days) c | Maximum Viral Titer (log10 EID50/mL) d | Area under the Viral Shedding Curve (AUC) d | |||||
| AUC of the Entire Observation Period | AUC of the Early Stages of Infection (1–3 dpi) | |||||||
| Kagawa11C e | E1 | 4.63 | 5.60 (4–6) | Trachea | 5 (4–5) | 5.32 (4.32–6.01) | 163,355 (60,078–681,723) | 10,012 (2099–94,938) |
| Cloaca | 5 (3–5) | 4.32 (3.87–6.02) | 41,285 (7419–574,373) | 1638 (340–7339) | ||||
| ChibaC1T | E2 | 3.50 | 3.17 (2–4) | Trachea | 3 (1–4) | 6.70 (5.53–7.70) | 2,743,748 (340,644–25,348,652) | 1,203,339 (178,330–17,073,701) |
| Cloaca | 3 (2–4) | 4.16 (2.53–6.02) | 15,162 (346–520,507) | 2378 (189–49,999) | ||||
| FukuokaT1 | E3 | 2.75 | 3.20 (3–4) | Trachea | 3 (2–4) | 6.53 (6.20–7.20) | 1,916,183 (948,681–8,146,128) | 1,359,707 (204,386–8,146,128) |
| Cloaca | 3 (2–3) | 5.20 (4.32–6.53) | 103,086 (13,371–1,705,148) | 82,347 (1129–270,130) | ||||
| Toyama16T | E5 | 4.50 | 4.40 (3–6) | Trachea | 3 (3–4) | 6.32 (5.53–6.70) | 1,269,541 (172,692–3,811,719) | 261,854 (0–3,811,719) |
| Cloaca | 3 (2–4) | 5.20 (4.02–5.87) | 79,210 (12,695–369,713) | 7416 (0–158,217) | ||||
| Tokushima4T | E7 | 4.63 | 5.40 (4–7) | Trachea | 4 (3–4) | 6.53 (6.32–6.87) | 3,901,427 (1,921,413–5,068,124) | 112,761 (0–126,507) |
| Cloaca | 4 (3–4) | 4.53 (2.20–6.87) | 36,484 (489–3,701,117) | 78 (0–23,715) | ||||
| Virus Strain | Genotype | Transmission Rate (%) a | Viral Shedding in the Virus-Infected Cohabiting Chickens b | |||
|---|---|---|---|---|---|---|
| Swab | Shedding Period (Days) | Maximum Viral Titer (log10 EID50/mL) | Area under the Viral Shedding Curve (AUC) | |||
| Kagawa11C | E1 | 16.7 (1/6) | Trachea | 5 | 6.32 | 531,852 |
| Cloaca | 5 | 5.01 | 361 | |||
| ChibaC1T | E2 | 100 (6/6) | Trachea | 3 (1–3) | 6.76 (5.45–7.87) | 7,407,674 (281,323–73,394,176) |
| Cloaca | 2.5 (1–3) | 3.77 (2.87–4.70) | 9391 (1151–49,998) | |||
| FukuokaT1 | E3 | 83.3 (5/6) | Trachea | 3 (2–5) | 7.32 (7.02–7.87) | 21,085,137 (6,979,946–50,148,240) |
| Cloaca | 3 (2–4) | 5.02 (3.53–5.70) | 83,638 (3433–568,124) | |||
| Toyama16T | E5 | 100 (6/6) | Trachea | 2 (1–3) | 6.11 (5.32–6.32) | 1,570,800 (105,424–3,529,670) |
| Cloaca | 1 (1–3) | 5.09 (4.20–5.87) | 145,522 (10,542–1,100,847) | |||
| Tokushima4T | E7 | 0 (0/6) | Trachea | - | - | - |
| Cloaca | - | - | - | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakuma, S.; Tanikawa, T.; Tsunekuni, R.; Mine, J.; Kumagai, A.; Miyazawa, K.; Takadate, Y.; Uchida, Y. Experimental Infection of Chickens with H5N8 High Pathogenicity Avian Influenza Viruses Isolated in Japan in the Winter of 2020–2021. Viruses 2023, 15, 2293. https://doi.org/10.3390/v15122293
Sakuma S, Tanikawa T, Tsunekuni R, Mine J, Kumagai A, Miyazawa K, Takadate Y, Uchida Y. Experimental Infection of Chickens with H5N8 High Pathogenicity Avian Influenza Viruses Isolated in Japan in the Winter of 2020–2021. Viruses. 2023; 15(12):2293. https://doi.org/10.3390/v15122293
Chicago/Turabian StyleSakuma, Saki, Taichiro Tanikawa, Ryota Tsunekuni, Junki Mine, Asuka Kumagai, Kohtaro Miyazawa, Yoshihiro Takadate, and Yuko Uchida. 2023. "Experimental Infection of Chickens with H5N8 High Pathogenicity Avian Influenza Viruses Isolated in Japan in the Winter of 2020–2021" Viruses 15, no. 12: 2293. https://doi.org/10.3390/v15122293