
Phosphorylation of Human Polyomavirus Large and Small T Antigens: An Ignored Research Field

 , , and
, , and
Abstract
:1. Introduction
2. Phosphorylation of Large T Antigen
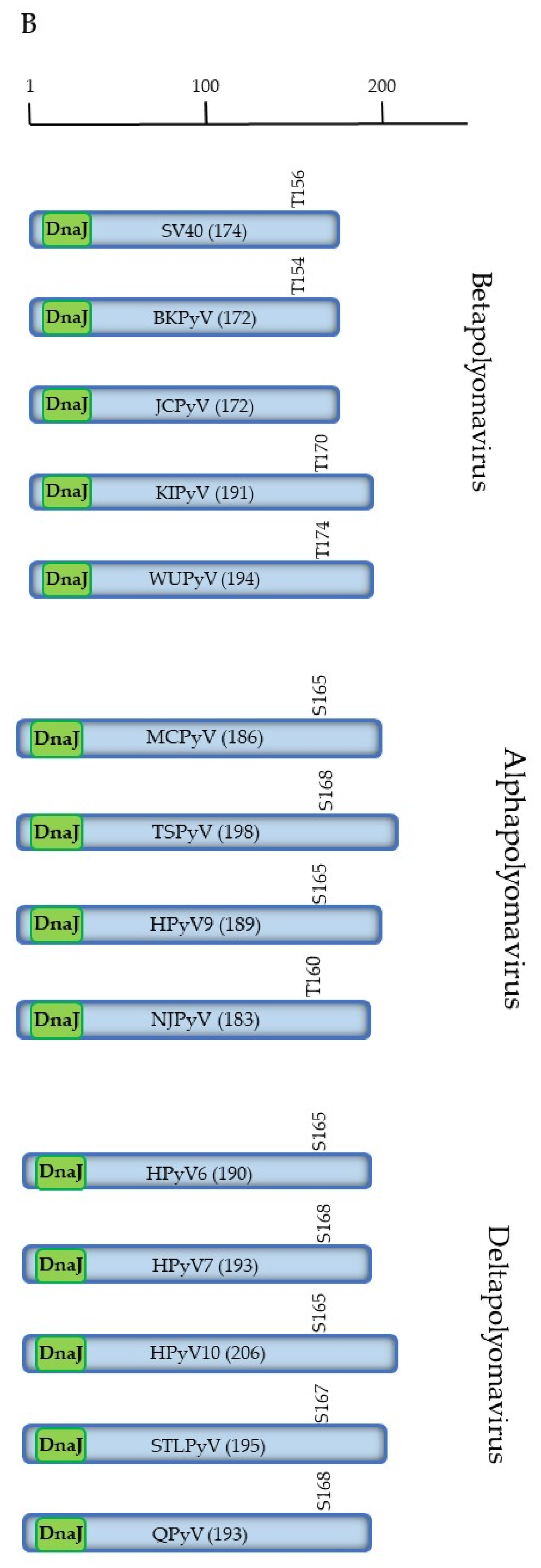
2.1. Functions of SV40 LTAg
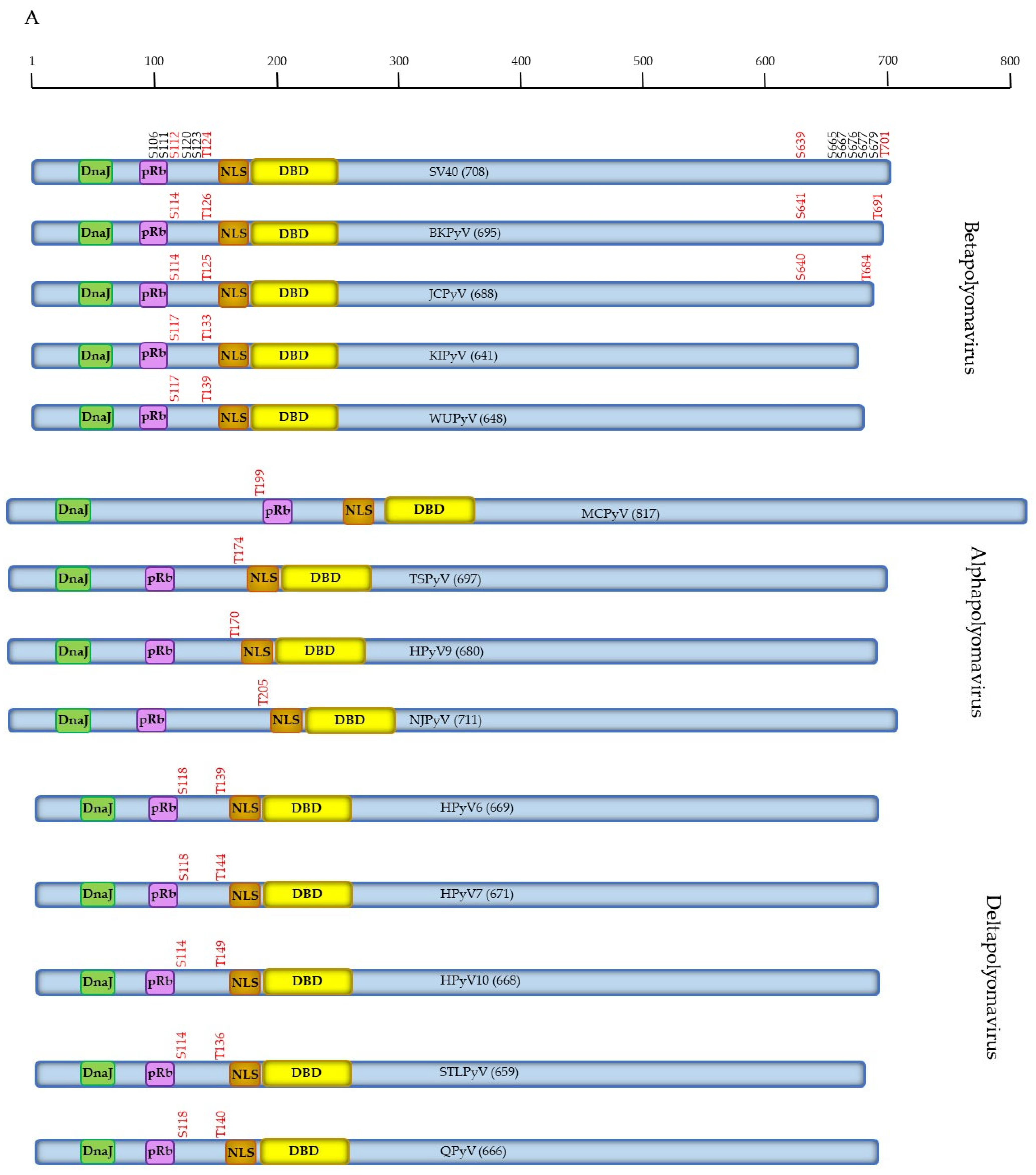
2.2. Putative and Proven Phosphorylation Sites in SV40 LTAg
2.3. Biological Implications of SV40 LTAg Phosphorylation
2.3.1. SV40 LTAg Phosphorylation and Subcellular Distribution
2.3.2. SV40 LTAg Phosphorylation and Replication
2.3.3. SV40 LTAg Phosphorylation and Protein Interaction
2.3.4. SV40 LTAg Phosphorylation and Transformation
2.4. Phosphorylation of HPyV LTAg
2.4.1. Phosphorylation of Human Betapolyomavirus LTAg
2.4.2. Phosphorylation of Human Alphapolyomavirus LTAg
Phosphorylation of Merkel Cell Polyomavirus Full-Length LTAg
Phosphorylation of Merkel Cell Polyomavirus Truncated LTAg
Phosphorylation of Human Alphapolyomavirus LTAg
2.4.3. Phosphorylation of Human Deltapolyomavirus LTAg
3. Phosphorylation of sTAg
3.1. Function of SV40 sTAg
3.2. Putative Phosphorylation Sites of SV40 sTAg
4. Gaps in Knowledge and Future Research Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Denu, J.M.; Stuckey, J.A.; Saper, M.A.; Dixon, J.E. Form and function in protein dephosphorylation. Cell 1996, 87, 361–364. [Google Scholar] [CrossRef]
- Ubersax, J.A.; Ferrell, J.E., Jr. Mechanisms of specificity in protein phosphorylation. Nat. Rev. Mol. Cell. Biol. 2007, 8, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The protein kinase complement of the human genome. Science 2002, 298, 1912–1934. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Dixon, J.E.; Manning, G. Genomics and evolution of protein phosphatases. Sci. Signal. 2017, 10, eaag1796. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P. The origins of protein phosphorylation. Nat. Cell. Biol. 2002, 4, E127–E130. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.V.; Blagoev, B.; Gnad, F.; Macek, B.; Kumar, C.; Mortensen, P.; Mann, M. Global, in vivo, and site-specific phosphorylation dynamics in signaling networks. Cell 2006, 127, 635–648. [Google Scholar] [CrossRef]
- Cohen, P. The regulation of protein function by multisite phosphorylation—A 25 year update. Trends Biochem. Sci. 2000, 25, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Nishi, H.; Hashimoto, K.; Panchenko, A.R. Phosphorylation in protein-protein binding: Effect on stability and function. Structure 2011, 19, 1807–1815. [Google Scholar] [CrossRef] [PubMed]
- Gjertsen, B.T.; Døskeland, S.O. Protein phosphorylation in apoptosis. Biochim. Biophys. Acta 1995, 1269, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, S.J.; James, D.E.; Mann, M. Protein Phosphorylation: A Major Switch Mechanism for Metabolic Regulation. Trends Endocrinol. Metab. 2015, 26, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Ardito, F.; Giuliani, M.; Perrone, D.; Troiano, G.; Lo Muzio, L. The crucial role of protein phosphorylation in cell signaling and its use as targeted therapy (Review). Int. J. Mol. Med. 2017, 40, 271–280. [Google Scholar] [CrossRef]
- Bilbrough, T.; Piemontese, E.; Seitz, O. Dissecting the role of protein phosphorylation: A chemical biology toolbox. Chem. Soc. Rev. 2022, 51, 5691–5730. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P. The role of protein phosphorylation in human health and disease. The Sir Hans Krebs Medal Lecture. Eur. J. Biochem. 2001, 268, 5001–5010. [Google Scholar] [CrossRef]
- Stebbing, J.; Lit, L.C.; Zhang, H.; Darrington, R.S.; Melaiu, O.; Rudraraju, B.; Giamas, G. The regulatory roles of phosphatases in cancer. Oncogene 2014, 33, 939–953. [Google Scholar] [CrossRef]
- Delom, F.; Chevet, E. Phosphoprotein analysis: From proteins to proteomes. Proteome Sci. 2006, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.R.; Veenstra, T.D. Characterization of Phosphorylated Proteins Using Mass Spectrometry. Curr. Protein Pept. Sci. 2021, 22, 148–157. [Google Scholar] [CrossRef]
- Fulcher, A.J.; Jans, D.A. Regulation of nucleocytoplasmic trafficking of viral proteins: An integral role in pathogenesis? Biochim. Biophys. Acta 2011, 1813, 2176–2190. [Google Scholar] [CrossRef] [PubMed]
- Yang, F. Post-translational Modification Control of HBV Biological Processes. Front. Microbiol. 2018, 9, 2661. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Mehta, D.; Mishra, N.; Nayak, D.; Sunil, S. Role of Host-Mediated Post-Translational Modifications (PTMs) in RNA Virus Pathogenesis. Int. J. Mol. Sci. 2020, 22, 323. [Google Scholar] [CrossRef]
- Zhou, T.; Wang, M.; Cheng, A.; Yang, Q.; Tian, B.; Wu, Y.; Jia, R.; Chen, S.; Liu, M.; Zhao, X.X.; et al. Regulation of alphaherpesvirus protein via post-translational phosphorylation. Vet. Res. 2022, 53, 93. [Google Scholar] [CrossRef]
- Cheng, N.; Liu, M.; Li, W.; Sun, B.; Liu, D.; Wang, G.; Shi, J.; Li, L. Protein post-translational modification in SARS-CoV-2 and host interaction. Front. Immunol. 2023, 13, 1068449. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Ren, Z.; Wang, Y. Serine-arginine protein kinases and their targets in viral infection and their inhibition. Cell. Mol. Life Sci. 2023, 80, 153. [Google Scholar] [CrossRef] [PubMed]
- Guan, K.L.; Broyles, S.S.; Dixon, J.E. A Tyr/Ser protein phosphatase encoded by vaccinia virus. Nature 1991, 350, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Lemon, B.; Traktman, P. The dual-specificity phosphatase encoded by vaccinia virus, VH1, is essential for viral transcription in vivo and in vitro. J. Virol. 1995, 69, 7823–7834. [Google Scholar] [CrossRef]
- Guan, K.L.; Dixon, J.E. Bacterial and viral protein tyrosine phosphatases. Semin. Cell. Biol. 1993, 4, 389–396. [Google Scholar] [CrossRef]
- Leader, D.P. Viral protein kinases and protein phosphatases. Pharmacol. Ther. 1993, 59, 343–389. [Google Scholar] [CrossRef] [PubMed]
- Jacob, T.; Van den Broeke, C.; Favoreel, H.W. Viral serine/threonine protein kinases. J. Virol. 2011, 85, 1158–1173. [Google Scholar] [CrossRef]
- Segovia, D.; Haouz, A.; Porley, D.; Olivero, N.; Martínez, M.; Mariadassou, M.; Berois, M.; André-Leroux, G.; Villarino, A. OH1 from Orf Virus: A New Tyrosine Phosphatase that Displays Distinct Structural Features and Triple Substrate Specificity. J. Mol. Biol. 2017, 429, 2816–2824. [Google Scholar] [CrossRef]
- Polyomaviridae Study Group of the International Committee on Taxonomy of Viruses; Calvignac-Spencer, S.; Feltkamp, M.C.; Daugherty, M.D.; Moens, U.; Ramqvist, T.; Johne, R.; Ehlers, B. A taxonomy update for the family Polyomaviridae. Arch. Virol. 2016, 161, 1739–1750. [Google Scholar] [CrossRef]
- Moens, U.; Calvignac-Spencer, S.; Lauber, C.; Ramqvist, T.; Feltkamp, M.C.W.; Daugherty, M.D.; Verschoor, E.J.; Ehlers, B.; Ictv Report Consortium. ICTV Virus Taxonomy Profile: Polyomaviridae. J. Gen. Virol. 2017, 98, 1159–1160. [Google Scholar] [CrossRef]
- Gardner, S.D.; Field, A.M.; Coleman, D.V.; Hulme, B. New human papovavirus (B.K.) isolated from urine after renal transplantation. Lancet 1971, 1, 1253–1257. [Google Scholar] [CrossRef]
- Padgett, B.L.; Walker, D.L.; ZuRhein, G.M.; Eckroade, R.J.; Dessel, B.H. Cultivation of papova-like virus from human brain with progressive multifocal leucoencephalopathy. Lancet 1971, 1, 1257–1260. [Google Scholar] [CrossRef]
- Allander, T.; Andreasson, K.; Gupta, S.; Bjerkner, A.; Bogdanovic, G.; Persson, M.A.; Dalianis, T.; Ramqvist, T.; Andersson, B. Identification of a third human polyomavirus. J. Virol. 2007, 81, 4130–4136. [Google Scholar] [CrossRef]
- Gaynor, A.M.; Nissen, M.D.; Whiley, D.M.; Mackay, I.M.; Lambert, S.B.; Wu, G.; Brennan, D.C.; Storch, G.A.; Sloots, T.P.; Wang, D. Identification of a novel polyomavirus from patients with acute respiratory tract infections. PLoS Pathog. 2007, 3, e64. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Shuda, M.; Chang, Y.; Moore, P.S. Clonal integration of a polyomavirus in human Merkel cell carcinoma. Science 2008, 319, 1096–1100. [Google Scholar] [CrossRef] [PubMed]
- Schowalter, R.M.; Pastrana, D.V.; Pumphrey, K.A.; Moyer, A.L.; Buck, C.B. Merkel cell polyomavirus and two previously unknown polyomaviruses are chronically shed from human skin. Cell Host Microbe 2010, 7, 509–515. [Google Scholar] [CrossRef]
- van der Meijden, E.; Janssens, R.W.; Lauber, C.; Bouwes Bavinck, J.N.; Gorbalenya, A.E.; Feltkamp, M.C. Discovery of a new human polyomavirus associated with trichodysplasia spinulosa in an immunocompromized patient. PLoS Pathog. 2010, 6, e1001024. [Google Scholar] [CrossRef]
- Scuda, N.; Hofmann, J.; Calvignac-Spencer, S.; Ruprecht, K.; Liman, P.; Kühn, J.; Hengel, H.; Ehlers, B. A novel human polyomavirus closely related to the african green monkey-derived lymphotropic polyomavirus. J. Virol. 2011, 85, 4586–4590. [Google Scholar] [CrossRef] [PubMed]
- Buck, C.B.; Phan, G.Q.; Raiji, M.T.; Murphy, P.M.; McDermott, D.H.; McBride, A.A. Complete genome sequence of a tenth human polyomavirus. J. Virol. 2012, 86, 10887. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.S.; Reyes, A.; Antonio, M.; Saha, D.; Ikumapayi, U.N.; Adeyemi, M.; Stine, O.C.; Skelton, R.; Brennan, D.C.; Mkakosya, R.S.; et al. Discovery of STL polyomavirus, a polyomavirus of ancestral recombinant origin that encodes a unique T antigen by alternative splicing. Virology 2013, 436, 295–303. [Google Scholar] [CrossRef]
- Korup, S.; Rietscher, J.; Calvignac-Spencer, S.; Trusch, F.; Hofmann, J.; Moens, U.; Sauer, I.; Voigt, S.; Schmuck, R.; Ehlers, B. Identification of a novel human polyomavirus in organs of the gastrointestinal tract. PLoS ONE 2013, 8, e58021. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Pereira, M.; Rhodes, R.H.; An, P.; Pipas, J.M.; Jain, K.; Kapoor, A.; Briese, T.; Faust, P.L.; Lipkin, W.I. Identification of a novel polyomavirus in a pancreatic transplant recipient with retinal blindness and vasculitic myopathy. J. Infect. Dis. 2014, 210, 1595–1599. [Google Scholar] [CrossRef] [PubMed]
- Gheit, T.; Dutta, S.; Oliver, J.; Robitaille, A.; Hampras, S.; Combes, J.D.; McKay-Chopin, S.; Le Calvez-Kelm, F.; Fenske, N.; Cherpelis, B.; et al. Isolation and characterization of a novel putative human polyomavirus. Virology 2017, 506, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Ondov, B.D.; Starrett, G.J.; Sappington, A.; Kostic, A.; Koren, S.; Buck, C.B.; Phillippy, A.M. Mash Screen: High-throughput sequence containment estimation for genome discovery. Genome Biol. 2019, 20, 232. [Google Scholar] [CrossRef]
- Lednicky, J.A.; Arrington, A.S.; Stewart, A.R.; Dai, X.M.; Wong, C.; Jafar, S.; Murphey-Corb, M.; Butel, J.S. Natural isolates of simian virus 40 from immunocompromised monkeys display extensive genetic heterogeneity: New implications for polyomavirus disease. J. Virol. 1998, 72, 3980–3990. [Google Scholar] [CrossRef]
- Sweet, B.H.; Hilleman, M.R. The vacuolating virus, S.V.40. Proc. Soc. Exp. Biol. Med. 1960, 105, 420–427. [Google Scholar] [CrossRef]
- Koliaskina, G.I. Virological and serological data on latent infection caused by OB40 (SV40) virus in macaca rhesus monkeys. Vopr. Virusol. 1963, 29, 450–452. [Google Scholar] [PubMed]
- Fagrouch, Z.; Karremans, K.; Deuzing, I.; van Gessel, S.; Niphuis, H.; Bogers, W.; Verschoor, E.J. Molecular analysis of a novel simian virus 40 (SV40) type in rhesus macaques and evidence for double infections with the classical SV40 type. J. Clin. Microbiol. 2011, 49, 1280–1286. [Google Scholar] [CrossRef]
- Gedvilaite, A.; Tryland, M.; Ulrich, R.G.; Schneider, J.; Kurmauskaite, V.; Moens, U.; Preugschas, H.; Calvignac-Spencer, S.; Ehlers, B. Novel polyomaviruses in shrews (Soricidae) with close similarity to human polyomavirus 12. J. Gen. Virol. 2017, 98, 3060–3067. [Google Scholar] [CrossRef] [PubMed]
- Kamminga, S.; van der Meijden, E.; Pesavento, P.; Buck, C.B.; Feltkamp, M.C.W. Serology Identifies LIPyV as a Feline Rather than a Human Polyomavirus. Viruses 2023, 15, 1546. [Google Scholar] [CrossRef]
- Prezioso, C.; Van Ghelue, M.; Pietropaolo, V.; Moens, U. Detection of Quebec Polyomavirus DNA in Samples from Different Patient Groups. Microorganisms 2021, 9, 1082. [Google Scholar] [CrossRef] [PubMed]
- DeCaprio, J.A.; Garcea, R.L. A cornucopia of human polyomaviruses. Nat. Rev. Microbiol. 2013, 11, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Moens, U.; Krumbholz, A.; Ehlers, B.; Zell, R.; Johne, R.; Calvignac-Spencer, S.; Lauber, C. Biology, evolution, and medical importance of polyomaviruses: An update. Infect. Genet. Evol. 2017, 54, 18–38. [Google Scholar] [CrossRef] [PubMed]
- Prins, C.; Frisque, R.J. JC virus T′ proteins encoded by alternatively spliced early mRNAs enhance T antigen-mediated viral DNA replication in human cells. J. Neurovirol. 2001, 7, 250–264. [Google Scholar]
- Carter, J.J.; Daugherty, M.D.; Qi, X.; Bheda-Malge, A.; Wipf, G.C.; Robinson, K.; Roman, A.; Malik, H.S.; Galloway, D.A. Identification of an overprinting gene in Merkel cell polyomavirus provides evolutionary insight into the birth of viral genes. Proc. Natl. Acad. Sci. USA 2013, 110, 12744–12749. [Google Scholar] [CrossRef] [PubMed]
- van der Meijden, E.; Kazem, S.; Dargel, C.A.; van Vuren, N.; Hensbergen, P.J.; Feltkamp, M.C. Characterization of T Antigens, Including Middle T and Alternative T, Expressed by the Human Polyomavirus Associated with Trichodysplasia Spinulosa. J. Virol. 2015, 89, 9427–9439. [Google Scholar] [CrossRef] [PubMed]
- Ponder, B.A.; Robbins, A.K.; Crawford, L.V. Phophorylation of polyoma and SV40 virus proteins. J. Gen. Virol. 1977, 37, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.L.; Hsu, P.H.; Fang, C.Y.; Chang, C.F.; Ou, W.C.; Wang, M.; Chang, D. Phosphorylation of Ser-80 of VP1 and Ser-254 of VP2 is essential for human BK virus propagation in tissue culture. J. Gen. Virol. 2011, 92, 2637–2645. [Google Scholar] [CrossRef] [PubMed]
- Simmons, D.T. SV40 large T antigen functions in DNA replication and transformation. Adv. Virus Res. 2000, 55, 75–134. [Google Scholar]
- Ali, S.H.; DeCaprio, J.A. Cellular transformation by SV40 large T antigen: Interaction with host proteins. Semin. Cancer Biol. 2001, 11, 15–23. [Google Scholar] [CrossRef]
- Sullivan, C.S.; Pipas, J.M. T antigens of simian virus 40: Molecular chaperones for viral replication and tumorigenesis. Microbiol. Mol. Biol. Rev. 2002, 66, 179–202. [Google Scholar] [CrossRef]
- Fanning, E. Simian virus 40 large T antigen: The puzzle, the pieces, and the emerging picture. J. Virol. 1992, 66, 1289–1293. [Google Scholar] [CrossRef]
- Ahuja, D.; Sáenz-Robles, M.T.; Pipas, J.M. SV40 large T antigen targets multiple cellular pathways to elicit cellular transformation. Oncogene 2005, 24, 7729–7745. [Google Scholar] [CrossRef] [PubMed]
- White, M.K.; Khalili, K. Interaction of retinoblastoma protein family members with large T-antigen of primate polyomaviruses. Oncogene 2006, 25, 5286–5293. [Google Scholar] [CrossRef] [PubMed]
- An, P.; Sáenz Robles, M.T.; Pipas, J.M. Large T antigens of polyomaviruses: Amazing molecular machines. Annu. Rev. Microbiol. 2012, 66, 213–236. [Google Scholar] [CrossRef]
- Topalis, D.; Andrei, G.; Snoeck, R. The large tumor antigen: A “Swiss Army knife” protein possessing the functions required for the polyomavirus life cycle. Antivir. Res. 2013, 97, 122–136. [Google Scholar] [CrossRef]
- Simmons, D.T.; Gai, D.; Parsons, R.; Debes, A.; Roy, R. Assembly of the replication initiation complex on SV40 origin DNA. Nucleic Acids Res. 2004, 32, 1103–1112. [Google Scholar] [CrossRef]
- Sowd, G.A.; Fanning, E. A wolf in sheep’s clothing: SV40 co-opts host genome maintenance proteins to replicate viral DNA. PLoS Pathog. 2012, 8, e1002994. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.V. SV40 and human cancer: A review of recent data. Int. J. Cancer 2007, 120, 215–223. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Mazzoni, E.; Bononi, I.; Tognon, M.; Martini, F. Association Between Simian Virus 40 and Human Tumors. Front. Oncol. 2019, 9, 670. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Gazdar, A.; Butel, J.S. SV40 and human mesothelioma. Transl. Lung Cancer Res. 2020, 9, S47–S59. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef]
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar] [CrossRef]
- Scheidtmann, K.H.; Buck, M.; Schneider, J.; Kalderon, D.; Fanning, E.; Smith, A.E. Biochemical characterization of phosphorylation site mutants of simian virus 40 large T antigen: Evidence for interaction between amino- and carboxy-terminal domains. J. Virol. 1991, 65, 1479–1490. [Google Scholar] [CrossRef] [PubMed]
- Grässer, F.A.; Scheidtmann, K.H.; Tuazon, P.T.; Traugh, J.A.; Walter, G. In vitro phosphorylation of SV40 large T antigen. Virology 1988, 165, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Scheidtmann, K.H.; Kaiser, A.; Carbone, A.; Walter, G. Phosphorylation of threonine in the proline-rich carboxy-terminal region of simian virus 40 large T antigen. J. Virol. 1981, 38, 59–69. [Google Scholar] [CrossRef]
- Cegielska, A.; Moarefi, I.; Fanning, E.; Virshup, D.M. T-antigen kinase inhibits simian virus 40 DNA replication by phosphorylation of intact T antigen on serines 120 and 123. J. Virol. 1994, 68, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Fanning, E. Mutations in the phosphorylation sites of simian virus 40 (SV40) T antigen alter its origin DNA-binding specificity for sites I or II and affect SV40 DNA replication activity. J. Virol. 1988, 62, 1598–1605. [Google Scholar] [CrossRef] [PubMed]
- Scheidtmann, K.H.; Schickedanz, J.; Walter, G.; Lanford, R.E.; Butel, J.S. Differential phosphorylation of cytoplasmic and nuclear variants of simian virus 40 large T antigen encoded by simian virus 40-adenovirus 7 hybrid viruses. J. Virol. 1984, 50, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Bierer, B.E.; Burakoff, S.J. T cell adhesion molecules. FASEB J. 1988, 2, 2584–2590. [Google Scholar] [CrossRef]
- Rihs, H.P.; Jans, D.A.; Fan, H.; Peters, R. The rate of nuclear cytoplasmic protein transport is determined by the casein kinase II site flanking the nuclear localization sequence of the SV40 T-antigen. EMBO J. 1991, 10, 633–639. [Google Scholar] [CrossRef]
- Chen, Y.R.; Lees-Miller, S.P.; Tegtmeyer, P.; Anderson, C.W. The human DNA-activated protein kinase phosphorylates simian virus 40 T antigen at amino- and carboxy-terminal sites. J. Virol. 1991, 65, 5131–5140. [Google Scholar] [CrossRef] [PubMed]
- Scheidtmann, K.H.; Virshup, D.M.; Kelly, T.J. Protein phosphatase 2A dephosphorylates simian virus 40 large T antigen specifically at residues involved in regulation of DNA-binding activity. J. Virol. 1991, 65, 209–2101. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, X.Y.; Wang, H.; Huq, M.S.; Iliakis, G. Roles of replication protein A and DNA-dependent protein kinase in the regulation of DNA replication following DNA damage. J. Biol. Chem. 1999, 274, 22060–22064. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Dodson, G.E.; Shaikh, S.; Rundell, K.; Tibbetts, R.S. Ataxia-telangiectasia-mutated (ATM) is a T-antigen kinase that controls SV40 viral replication in vivo. J. Biol. Chem. 2005, 280, 40195–40200. [Google Scholar] [CrossRef]
- McVey, D.; Brizuela, L.; Mohr, I.; Marshak, D.R.; Gluzman, Y.; Beach, D. Phosphorylation of large tumour antigen by cdc2 stimulates SV40 DNA replication. Nature 1989, 341, 503–507. [Google Scholar] [CrossRef]
- Fanning, E.; Nowak, B.; Burger, C. Detection and characterization of multiple forms of simian virus 40 large T antigen. J. Virol. 1981, 37, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Walser, A.; Rinke, Y.; Deppert, W. Only a minor fraction of plasma membrane-associated large T antigen in simian virus 40-transformed mouse tumor cells (mKSA) is exposed on the cell surface. J. Virol. 1989, 63, 3926–3933. [Google Scholar] [CrossRef] [PubMed]
- Kalderon, D.; Richardson, W.D.; Markham, A.F.; Smith, A.E. Sequence requirements for nuclear location of simian virus 40 large-T antigen. Nature 1984, 311, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.; Hunter, T. Phosphorylation of SV40 large T antigen in SV40 nucleoprotein complexes. Virology 1980, 107, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Fulcher, A.J.; Dias, M.M.; Jans, D.A. Binding of p110 retinoblastoma protein inhibits nuclear import of simian virus SV40 large tumor antigen. J. Biol. Chem. 2010, 285, 17744–17753. [Google Scholar] [CrossRef] [PubMed]
- Jans, D.A.; Ackermann, M.J.; Bischoff, J.R.; Beach, D.H.; Peters, R. p34cdc2-mediated phosphorylation at T124 inhibits nuclear import of SV-40 T antigen proteins. J. Cell Biol. 1991, 115, 1203–1212. [Google Scholar] [CrossRef]
- Hübner, S.; Xiao, C.Y.; Jans, D.A. The protein kinase CK2 site (Ser111/112) enhances recognition of the simian virus 40 large T-antigen nuclear localization sequence by importin. J. Biol. Chem. 1997, 272, 17191–17195. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.Y.; Hübner, S.; Jans, D.A. SV40 large tumor antigen nuclear import is regulated by the double-stranded DNA-dependent protein kinase site (serine 120) flanking the nuclear localization sequence. J. Biol. Chem. 1997, 272, 22191–22198. [Google Scholar] [CrossRef] [PubMed]
- Fulcher, A.J.; Roth, D.M.; Fatima, S.; Alvisi, G.; Jans, D.A. The BRCA-1 binding protein BRAP2 is a novel, negative regulator of nuclear import of viral proteins, dependent on phosphorylation flanking the nuclear localization signal. FASEB J. 2010, 24, 1454–1466. [Google Scholar] [CrossRef] [PubMed]
- Rihs, H.P.; Peters, R. Nuclear transport kinetics depend on phosphorylation-site-containing sequences flanking the karyophilic signal of the Simian virus 40 T-antigen. EMBO J. 1989, 8, 1479–1484. [Google Scholar] [CrossRef]
- Xiao, C.Y.; Jans, P.; Jans, D.A. Negative charge at the protein kinase CK2 site enhances recognition of the SV40 large T-antigen NLS by importin: Effect of conformation. FEBS Lett. 1998, 440, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Jans, D.A.; Jans, P. Negative charge at the casein kinase II site flanking the nuclear localization signal of the SV40 large T-antigen is mechanistically important for enhanced nuclear import. Oncogene 1994, 9, 2961–2968. [Google Scholar] [PubMed]
- Dean, F.B.; Borowiec, J.A.; Eki, T.; Hurwitz, J. The simian virus 40 T antigen double hexamer assembles around the DNA at the replication origin. J. Biol. Chem. 1992, 267, 14129–14137. [Google Scholar] [CrossRef] [PubMed]
- Valle, M.; Gruss, C.; Halmer, L.; Carazo, J.M.; Donate, L.E. Large T-antigen double hexamers imaged at the simian virus 40 origin of replication. Mol. Cell. Biol. 2000, 20, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Baumann, E.A. DNA-binding properties of phosphorylated and dephosphorylated D2-T antigen, a simian-virus-40 T-antigen-related protein. Eur. J. Biochem. 1985, 147, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Prives, C.; Beck, Y.; Gidoni, D.; Oren, M.; Shure, H. DNA binding and sedimentation properties of SV40 T antigens synthesized in vivo and in vitro. Cold Spring Harb. Symp. Quant. Biol. 1980, 44, 123–130. [Google Scholar] [CrossRef]
- Scheidtmann, K.H.; Hardung, M.; Echle, B.; Walter, G. DNA-binding activity of simian virus 40 large T antigen correlates with a distinct phosphorylation state. J. Virol. 1984, 50, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Simmons, D.T.; Chou, W.; Rodgers, K. Phosphorylation downregulates the DNA-binding activity of simian virus 40 T antigen. J. Virol. 1986, 60, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Grässer, F.A.; Mann, K.; Walter, G. Removal of serine phosphates from simian virus 40 large T antigen increases its ability to stimulate DNA replication in vitro but has no effect on ATPase and DNA binding. J. Virol. 1987, 61, 3373–3380. [Google Scholar] [CrossRef] [PubMed]
- Mohr, I.J.; Stillman, B.; Gluzman, Y. Regulation of SV40 DNA replication by phosphorylation of T antigen. EMBO J. 1987, 6, 153–160. [Google Scholar] [CrossRef]
- Shaw, S.B.; Tegtmeyer, P. Binding of dephosphorylated A protein to SV40 DNA. Virology 1981, 115, 88–96. [Google Scholar] [CrossRef]
- Enserink, J.M.; Kolodner, R.D. An overview of Cdk1-controlled targets and processes. Cell Div. 2010, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- McVey, D.; Ray, S.; Gluzman, Y.; Berger, L.; Wildeman, A.G.; Marshak, D.R.; Tegtmeyer, P. cdc2 phosphorylation of threonine 124 activates the origin-unwinding functions of simian virus 40 T antigen. J. Virol. 1993, 67, 5206–5215. [Google Scholar] [CrossRef] [PubMed]
- Barbaro, B.A.; Sreekumar, K.R.; Winters, D.R.; Prack, A.E.; Bullock, P.A. Phosphorylation of simian virus 40 T antigen on Thr 124 selectively promotes double-hexamer formation on subfragments of the viral core origin. J. Virol. 2000, 74, 8601–8613. [Google Scholar] [CrossRef]
- Moarefi, I.F.; Small, D.; Gilbert, I.; Höpfner, M.; Randall, S.K.; Schneider, C.; Russo, A.A.; Ramsperger, U.; Arthur, A.K.; Stahl, H.; et al. Mutation of the cyclin-dependent kinase phosphorylation site in simian virus 40 (SV40) large T antigen specifically blocks SV40 origin DNA unwinding. J. Virol. 1993, 67, 4992–5002. [Google Scholar] [CrossRef] [PubMed]
- Scheidtmann, K.H. Phosphorylation of simian virus 40 large T antigen: Cytoplasmic and nuclear phophorylation sites differ in their metabolic stability. Virology 1986, 150, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Virshup, D.M.; Kauffman, M.G.; Kelly, T.J. Activation of SV40 DNA replication in vitro by cellular protein phosphatase 2A. EMBO J. 1989, 8, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Virshup, D.M.; Russo, A.A.; Kelly, T.J. Mechanism of activation of simian virus 40 DNA replication by protein phosphatase 2A. Mol. Cell. Biol. 1992, 12, 4883–4895. [Google Scholar] [PubMed]
- Scheidtmann, K.H.; Mumby, M.C.; Rundell, K.; Walter, G. Dephosphorylation of simian virus 40 large-T antigen and p53 protein by protein phosphatase 2A: Inhibition by small-t antigen. Mol. Cell. Biol. 1991, 11, 1996–2003. [Google Scholar] [PubMed]
- Welcker, M.; Clurman, B.E. FBW7 ubiquitin ligase: A tumour suppressor at the crossroads of cell division, growth and differentiation. Nat. Rev. Cancer 2008, 8, 83–93. [Google Scholar] [CrossRef]
- Welcker, M.; Clurman, B.E. The SV40 large T antigen contains a decoy phosphodegron that mediates its interactions with Fbw7/hCdc4. J. Biol. Chem. 2005, 280, 7654–7658. [Google Scholar] [CrossRef] [PubMed]
- Kalderon, D.; Smith, A.E. In vitro mutagenesis of a putative DNA binding domain of SV40 large-T. Virology 1984, 139, 109–137. [Google Scholar] [CrossRef]
- Prives, C. The replication functions of SV40 T antigen are regulated by phosphorylation. Cell 1990, 61, 735–738. [Google Scholar] [CrossRef]
- Swenson, J.J.; Frisque, R.J. Biochemical characterization and localization of JC virus large T antigen phosphorylation domains. Virology 1995, 212, 295–308. [Google Scholar] [CrossRef]
- Swenson, J.J.; Trowbridge, P.W.; Frisque, R.J. Replication activity of JC virus large T antigen phosphorylation and zinc finger domain mutants. J. Neurovirol. 1996, 2, 78–86. [Google Scholar] [CrossRef]
- Tyagarajan, S.K.; Frisque, R.J. Stability and function of JC virus large T antigen and T′ proteins are altered by mutation of their phosphorylated threonine 125 residues. J. Virol. 2006, 80, 2083–2091. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, S.Y.; Spiegelman, V.S.; Kumar, K.G. The many faces of beta-TrCP E3 ubiquitin ligases: Reflections in the magic mirror of cancer. Oncogene 2004, 23, 2028–2036. [Google Scholar] [CrossRef]
- Reviriego-Mendoza, M.M.; Frisque, R.J. Interaction and co-localization of JC virus large T antigen and the F-box protein β-transducin-repeat containing protein. Virology 2011, 410, 119–128. [Google Scholar] [CrossRef]
- Chang, Y.; Moore, P.S. Merkel cell carcinoma: A virus-induced human cancer. Annu. Rev. Pathol. 2012, 7, 123–144. [Google Scholar] [CrossRef]
- Starrett, G.J.; Buck, C.B. The case for BK polyomavirus as a cause of bladder cancer. Curr. Opin. Virol. 2019, 39, 8–15. [Google Scholar] [CrossRef]
- Klufah, F.; Mobaraki, G.; Liu, D.; Alharbi, R.A.; Kurz, A.K.; Speel, E.J.M.; Winnepenninckx, V.; Zur Hausen, A. Emerging role of human polyomaviruses 6 and 7 in human cancers. Infect. Agent Cancer. 2021, 16, 35. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.Y.; Chen, H.Y.; Wang, M.; Chen, P.L.; Chang, C.F.; Chen, L.S.; Shen, C.H.; Ou, W.C.; Tsai, M.D.; Hsu, P.H.; et al. Global analysis of modifications of the human BK virus structural proteins by LC-MS/MS. Virology 2010, 402, 164–176. [Google Scholar] [CrossRef]
- Harris, K.F.; Christensen, J.B.; Imperiale, M.J. BK virus large T antigen: Interactions with the retinoblastoma family of tumor suppressor proteins and effects on cellular growth control. J. Virol. 1996, 70, 2378–2386. [Google Scholar] [CrossRef]
- Becker, J.C.; Stang, A.; DeCaprio, J.A.; Cerroni, L.; Lebbé, C.; Veness, M.; Nghiem, P. Merkel cell carcinoma. Nat. Rev. Dis. Primers 2017, 3, 17077. [Google Scholar] [CrossRef] [PubMed]
- Schrama, D.; Sarosi, E.M.; Adam, C.; Ritter, C.; Kaemmerer, U.; Klopocki, E.; König, E.M.; Utikal, J.; Becker, J.C.; Houben, R. Characterization of six Merkel cell polyomavirus-positive Merkel cell carcinoma cell lines: Integration pattern suggest that large T antigen truncating events occur before or during integration. Int. J. Cancer. 2019, 145, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Schrama, D.; Hesbacher, S.; Angermeyer, S.; Schlosser, A.; Haferkamp, S.; Aue, A.; Adam, C.; Weber, A.; Schmidt, M.; Houben, R. Serine 220 phosphorylation of the Merkel cell polyomavirus large T antigen crucially supports growth of Merkel cell carcinoma cells. Int. J. Cancer. 2016, 138, 1153–1162. [Google Scholar] [CrossRef]
- Diaz, J.; Wang, X.; Tsang, S.H.; Jiao, J.; You, J. Phosphorylation of large T antigen regulates merkel cell polyomavirus replication. Cancers 2014, 6, 1464–1486. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Diaz, J.; Wang, X.; Tsang, S.H.; You, J. Phosphorylation of Merkel cell polyomavirus large tumor antigen at serine 816 by ATM kinase induces apoptosis in host cells. J. Biol. Chem. 2015, 290, 1874–1884. [Google Scholar] [CrossRef] [PubMed]
- Falquet, M.; Prezioso, C.; Ludvigsen, M.; Bruun, J.A.; Passerini, S.; Sveinbjørnsson, B.; Pietropaolo, V.; Moens, U. Regulation of Transcriptional Activity of Merkel Cell Polyomavirus Large T-Antigen by PKA-Mediated Phosphorylation. Int. J. Mol. Sci. 2023, 24, 895. [Google Scholar] [CrossRef]
- Kwun, H.J.; Chang, Y.; Moore, P.S. Protein-mediated viral latency is a novel mechanism for Merkel cell polyomavirus persistence. Proc. Natl. Acad. Sci. USA 2017, 114, E4040–E4047. [Google Scholar] [CrossRef]
- Nwogu, N.; Ortiz, L.E.; Kwun, H.J. Merkel Cell Polyomavirus Large T Antigen Unique Domain Regulates Its Own Protein Stability and Cell Growth. Viruses 2020, 12, 1043. [Google Scholar] [CrossRef]
- Alvarez Orellana, J.; Kwun, H.J.; Artusi, S.; Chang, Y.; Moore, P.S. Sirolimus and Other Mechanistic Target of Rapamycin Inhibitors Directly Activate Latent Pathogenic Human Polyomavirus Replication. J. Infect. Dis. 2021, 224, 1160–1169. [Google Scholar] [CrossRef] [PubMed]
- Caplan, S.; Hartnell, L.M.; Aguilar, R.C.; Naslavsky, N.; Bonifacino, J.S. Human Vam6p promotes lysosome clustering and fusion in vivo. J. Cell Biol. 2001, 154, 109–122. [Google Scholar] [CrossRef]
- Liu, X.; Hein, J.; Richardson, S.C.; Basse, P.H.; Toptan, T.; Moore, P.S.; Gjoerup, O.V.; Chang, Y. Merkel cell polyomavirus large T antigen disrupts lysosome clustering by translocating human Vam6p from the cytoplasm to the nucleus. J. Biol. Chem. 2011, 286, 17079–17090. [Google Scholar] [CrossRef]
- Moens, U.; Seternes, O.M.; Johansen, B.; Rekvig, O.P. Mechanisms of transcriptional regulation of cellular genes by SV40 large T- and small T-antigens. Virus Genes 1997, 15, 135–154. [Google Scholar] [CrossRef]
- Montano, X.; Millikan, R.; Milhaven, J.M.; Newsom, D.A.; Ludlow, J.W.; Arthur, A.K.; Fanning, E.; Bikel, I.; Livingston, D.M. Simian virus 40 small tumor antigen and an amino-terminal domain of large tumor antigen share a common transforming function. Proc. Natl. Acad. Sci. USA 1990, 87, 7448–7452. [Google Scholar] [CrossRef]
- Sontag, J.M.; Sontag, E. Regulation of cell adhesion by PP2A and SV40 small tumor antigen: An important link to cell transformation. Cell. Mol. Life Sci. 2006, 63, 2979–2991. [Google Scholar] [CrossRef]
- Pietropaolo, V.; Prezioso, C.; Moens, U. Merkel Cell Polyomavirus and Merkel Cell Carcinoma. Cancers 2020, 12, 1774. [Google Scholar] [CrossRef] [PubMed]
- Schwyzer, M.; Weil, R.; Frank, G.; Zuber, H. Amino acid sequence analysis of fragments generated by partial proteolysis from large simian virus 40 tumor antigen. J. Biol. Chem. 1980, 255, 5627–5634. [Google Scholar] [CrossRef] [PubMed]
- Hassauer, M.; Scheidtmann, K.H.; Walter, G. Mapping of phosphorylation sites in polyomavirus large T antigen. J. Virol. 1986, 58, 805–816. [Google Scholar] [CrossRef]
- Chatterjee, A.; Bockus, B.J.; Gjørup, O.V.; Schaffhausen, B.S. Phosphorylation sites in polyomavirus large T antigen that regulate its function in viral, but not cellular, DNA synthesis. J. Virol. 1997, 71, 6472–6478. [Google Scholar] [CrossRef] [PubMed]
- Daly, L.A.; Clarke, C.J.; Po, A.; Oswald, S.O.; Eyers, C.E. Considerations for defining +80 Da mass shifts in mass spectrometry-based proteomics: Phosphorylation and beyond. Chem. Commun. 2023, 59, 11484–11499. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Zhang, J.; Chen, Z.; Pan, W.; Chen, Z.; Yan, Y.; Dai, J. Glycosylation of viral proteins: Implication in virus-host interaction and virulence. Virulence 2022, 13, 670–683. [Google Scholar] [CrossRef]
- Dalianis, T.; Hirsch, H.H. Human polyomaviruses in disease and cancer. Virology 2013, 437, 63–72. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Residue Mutant | Protein Kinase | Support of Virion Production 1 | Transforming Activity 2 | DNA Rep 3 | Site I Binding 4 | Site II Binding 4 | Expression Levels 5 | Nuclear Localization 6 | ATPase Activity 7 | References |
|---|---|---|---|---|---|---|---|---|---|---|
| S106A | CKI, CKII, GSK3 | comparable | comparable | same | same | same | slightly reduced | same | intact | [74,75,78,79,80] |
| S106F | CKI, CKII, GSK3 | NT 8 | reduced | same | NT | NT | NT | NT | NT | [74] |
| S111A/ S112C | CKII, DNAPK | reduced | comparable | reduced | same | same | same | reduced | intact | [75,78,79,81,82] |
| S112D | CKII, DNAPK | comparable | comparable | comparable | same | same | NT | enhanced | NT | [74,75,79,81,82] |
| S120A | ATM, CKI, DNAPK | none | comparable | none | same | same | same | reduced | intact | [74,77,78,80,82,83,84,85] |
| S123A | CKI, DNAPK | none | comparable | none | same | enhanced | same | Same | intact | [74,75,77,78,79,82,83] |
| T124A | CKI, CDK1 | none | comparable | none | same | none | slightly reduced | increased | intact | [74,75,78,79,86] |
| T124E | CKI, CDK1 | not tested | comparable | low | same | none | same | same | intact | [78] |
| S639A | enhanced reduced | enhanced | reduced | same | same | same | same | intact | [74,78,79] | |
| S665 | DNAPK | NT | NT | NT | NT | NT | NT | NT | NT | [82] |
| S667 | DNAPK | NT | NT | NT | NT | NT | NT | NT | NT | [82] |
| S676A 9 | CKI | enhanced reduced | enhanced | reduced | reduced | same | same | same | intact | [74,75,77,78,79] |
| S677A | CKI, DNAPK | reduced | reduced | same | none | same | same | same | intact | [74,75,78,79,82,83,87] |
| S679A | CKI | reduced | reduced | enhanced | same | enhanced | same | same | intact | [74,75,78,79,83,87] |
| T701A | comparable | comparable | same | same | same | same | same | intact | [74,78,79] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moens, U.; Passerini, S.; Falquet, M.; Sveinbjørnsson, B.; Pietropaolo, V. Phosphorylation of Human Polyomavirus Large and Small T Antigens: An Ignored Research Field. Viruses 2023, 15, 2235. https://doi.org/10.3390/v15112235
Moens U, Passerini S, Falquet M, Sveinbjørnsson B, Pietropaolo V. Phosphorylation of Human Polyomavirus Large and Small T Antigens: An Ignored Research Field. Viruses. 2023; 15(11):2235. https://doi.org/10.3390/v15112235
Chicago/Turabian StyleMoens, Ugo, Sara Passerini, Mar Falquet, Baldur Sveinbjørnsson, and Valeria Pietropaolo. 2023. "Phosphorylation of Human Polyomavirus Large and Small T Antigens: An Ignored Research Field" Viruses 15, no. 11: 2235. https://doi.org/10.3390/v15112235