
In Vivo and In Vitro Characterization of the Recently Emergent PRRSV 1-4-4 L1C Variant (L1C.5) in Comparison with Other PRRSV-2 Lineage 1 Isolates

 , , , , , , , ,
, , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Virus Isolates
2.3. Animal Source and Approval
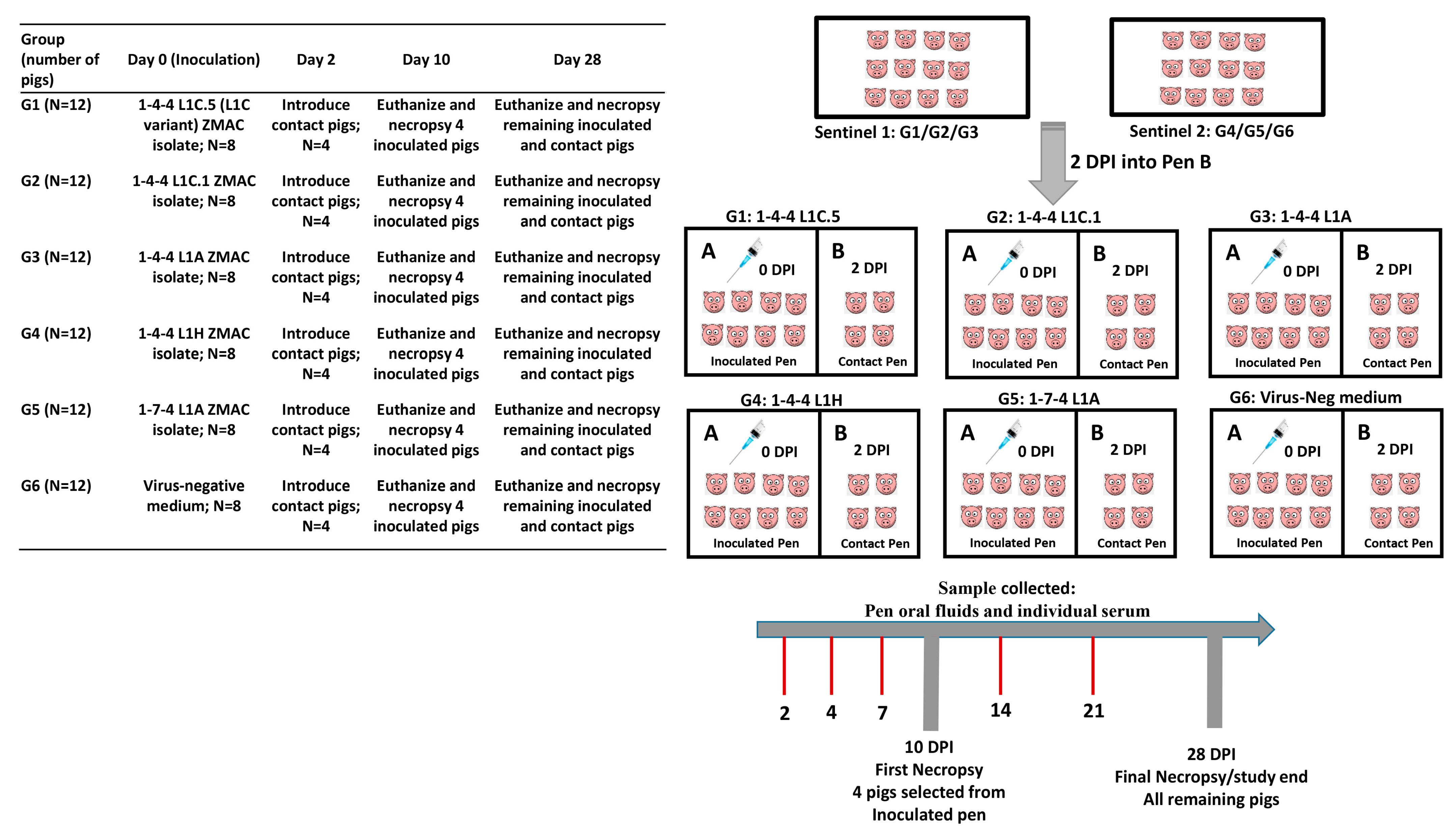
2.4. Animal Study Design
2.5. Sample Processing
2.6. Nucleic Acid Extraction and Quantitative PRRSV Real-Time RT-PCR
2.7. PRRSV Antibody Measurement
2.8. Bacterial Culture on Lung and Brain Homogenates
2.9. Gross Lung Pathology, Histopathology, and Immunohistochemistry
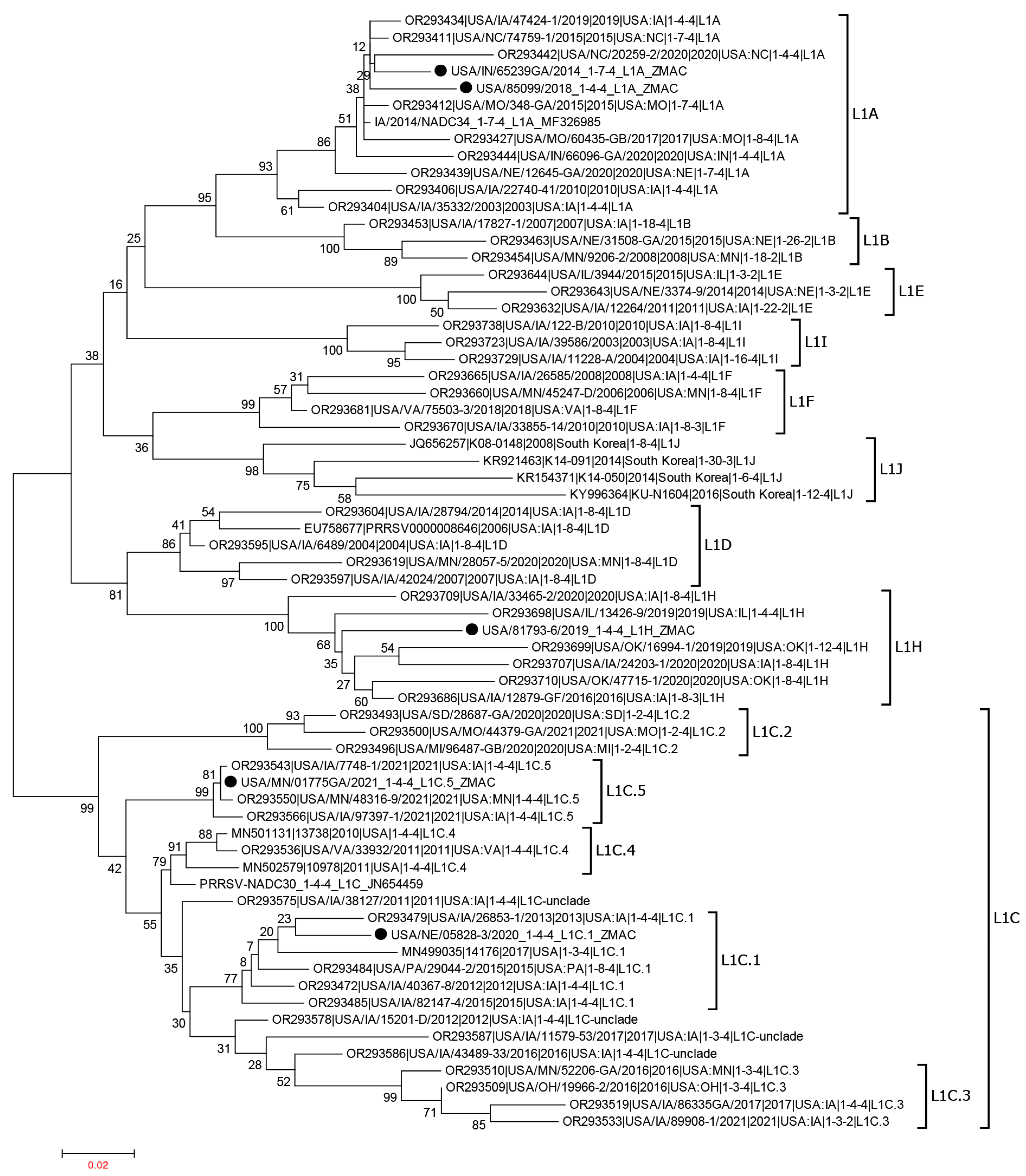
2.10. Whole Genome Sequence Analysis of PRRSV Isolates
2.11. Multistep Growth Curve in Cell Culture
2.12. Statistical Analysis
3. Results
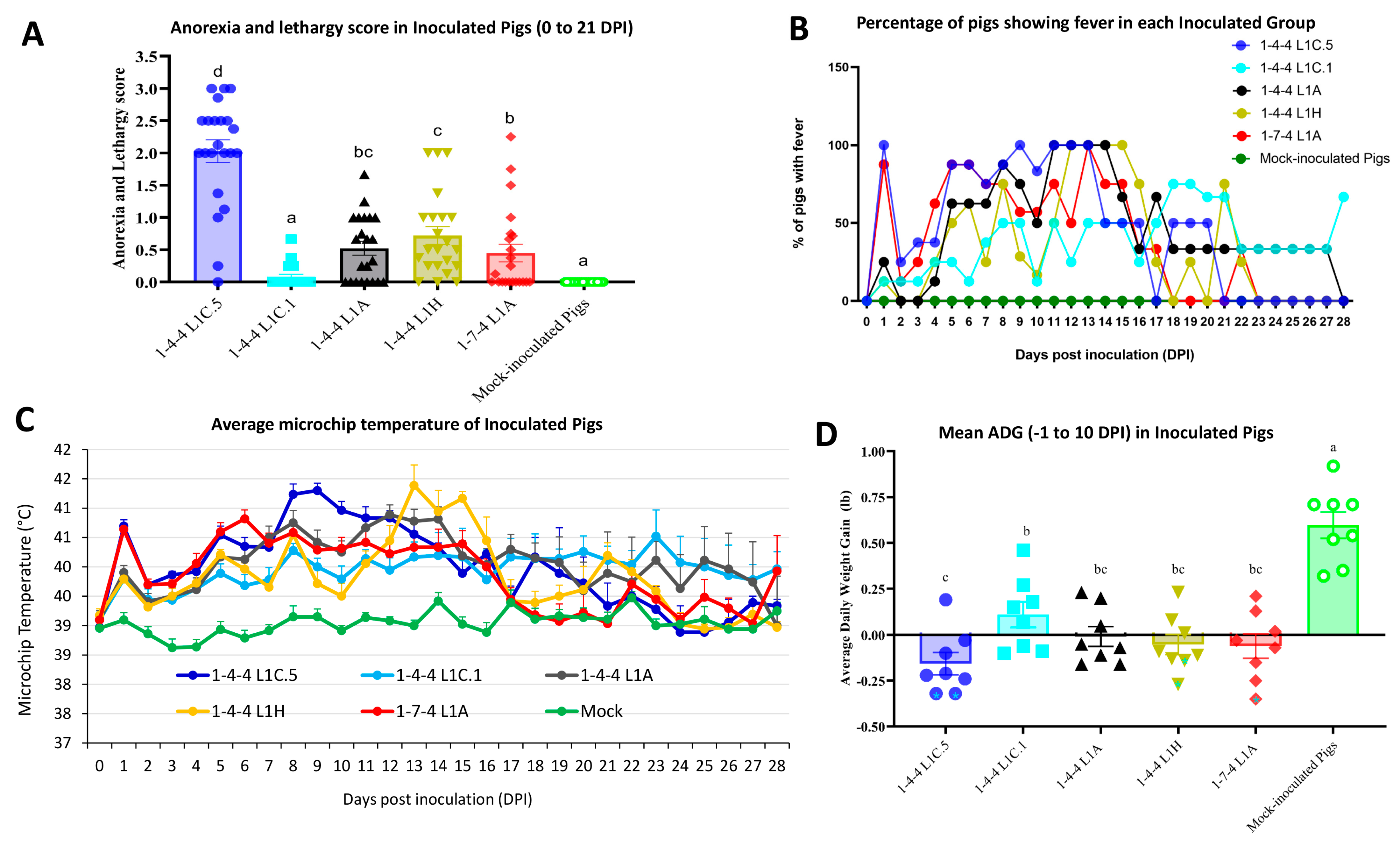
3.1. Clinical Observations (Anorexia and Lethargy, Mortality, Microchip Temperature, and ADG) in Inoculated Pigs
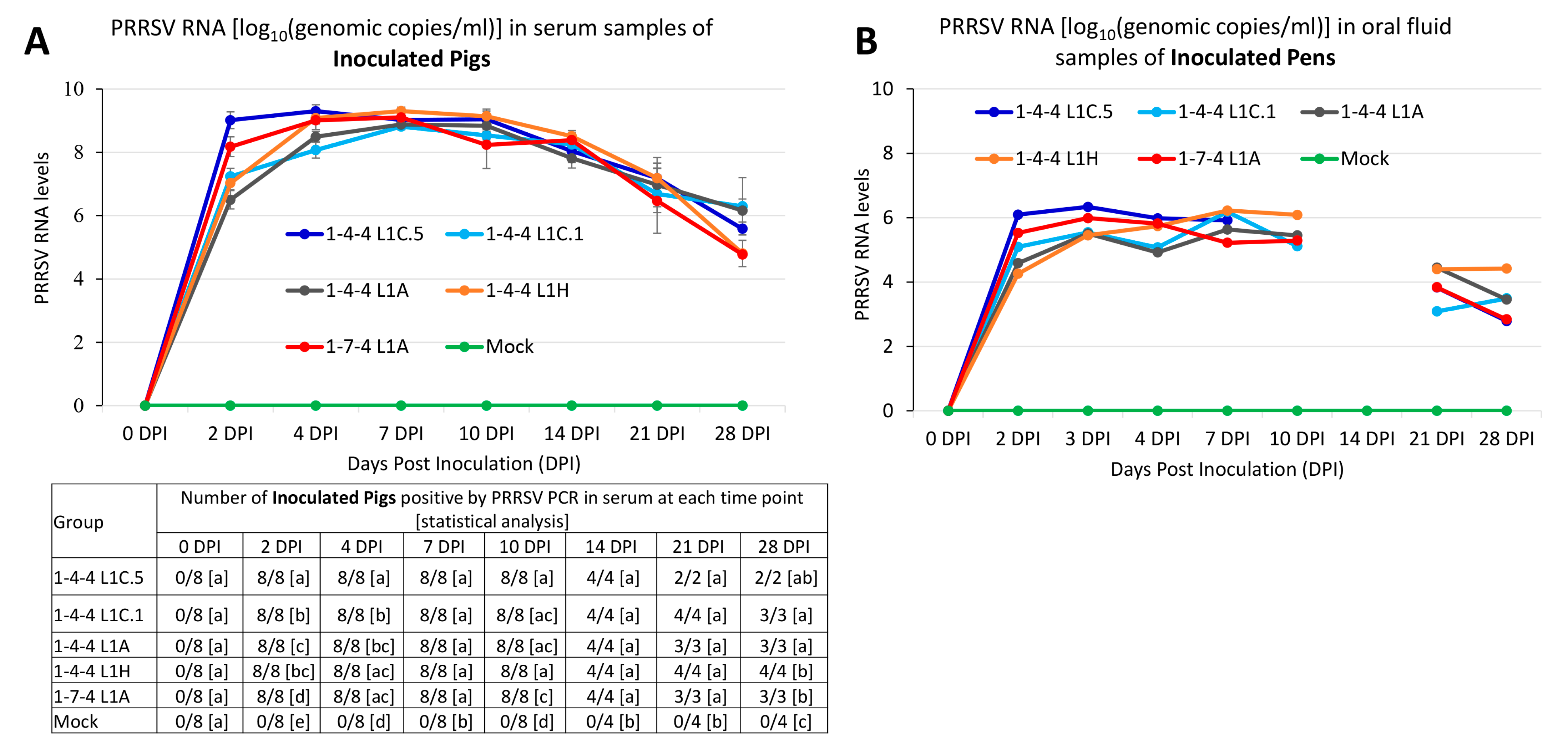
3.2. Viral Loads in Serum and Oral Fluid Samples of Inoculated Pigs
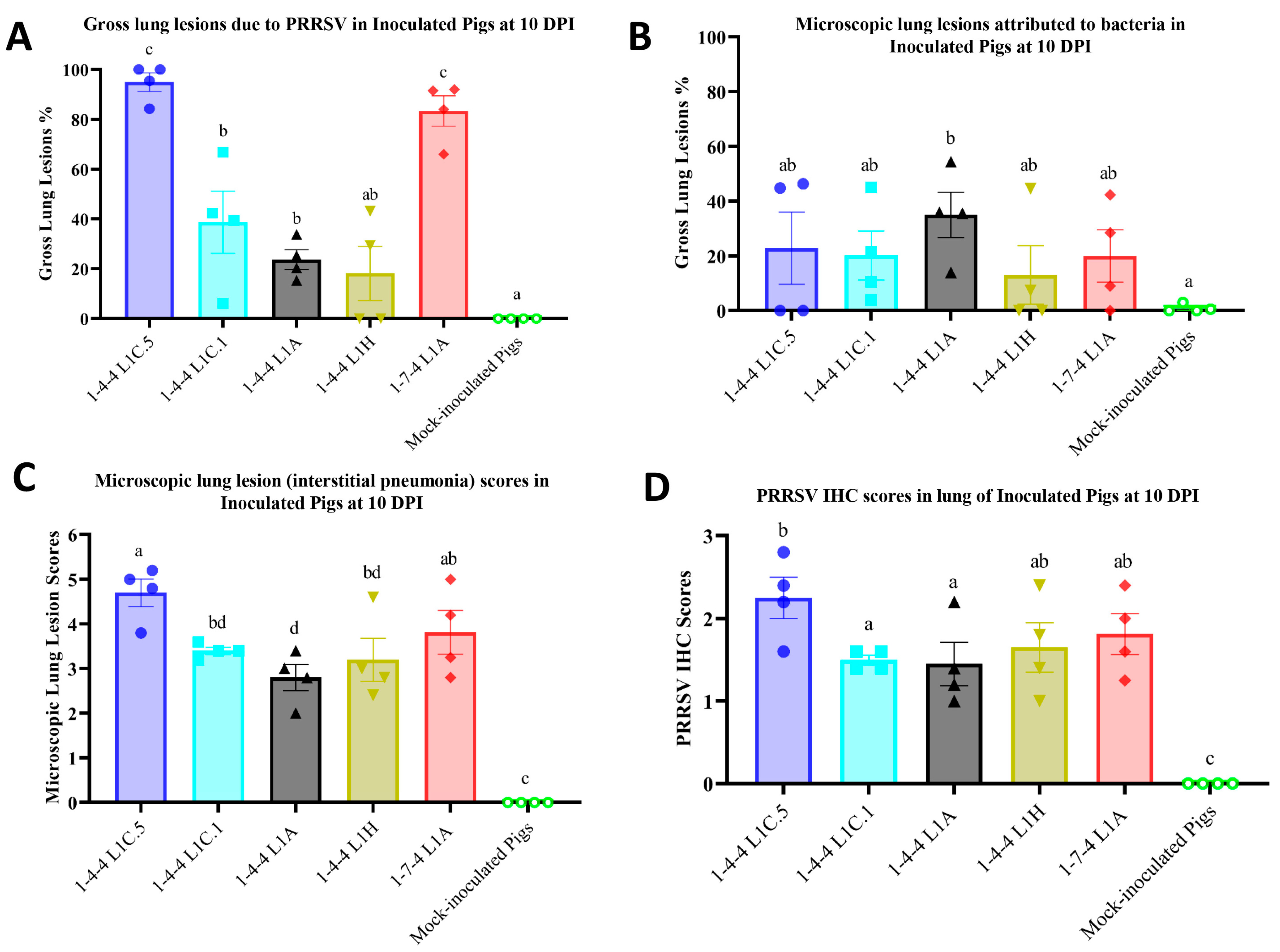
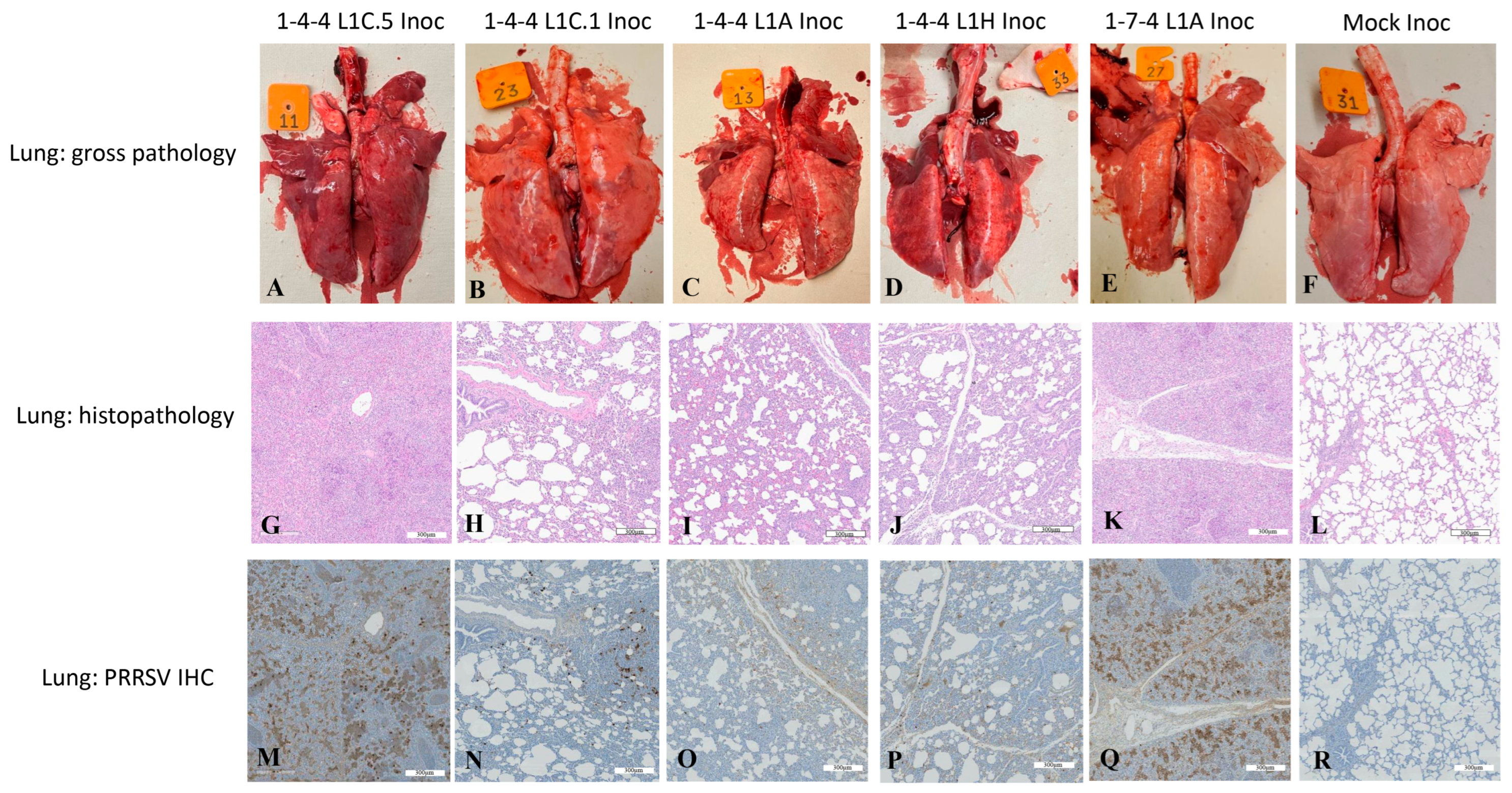
3.3. Gross Lung Lesions of Inoculated Pigs
3.4. Lung Microscopic Lesions and Immunohistochemistry Scores in Inoculated Pigs
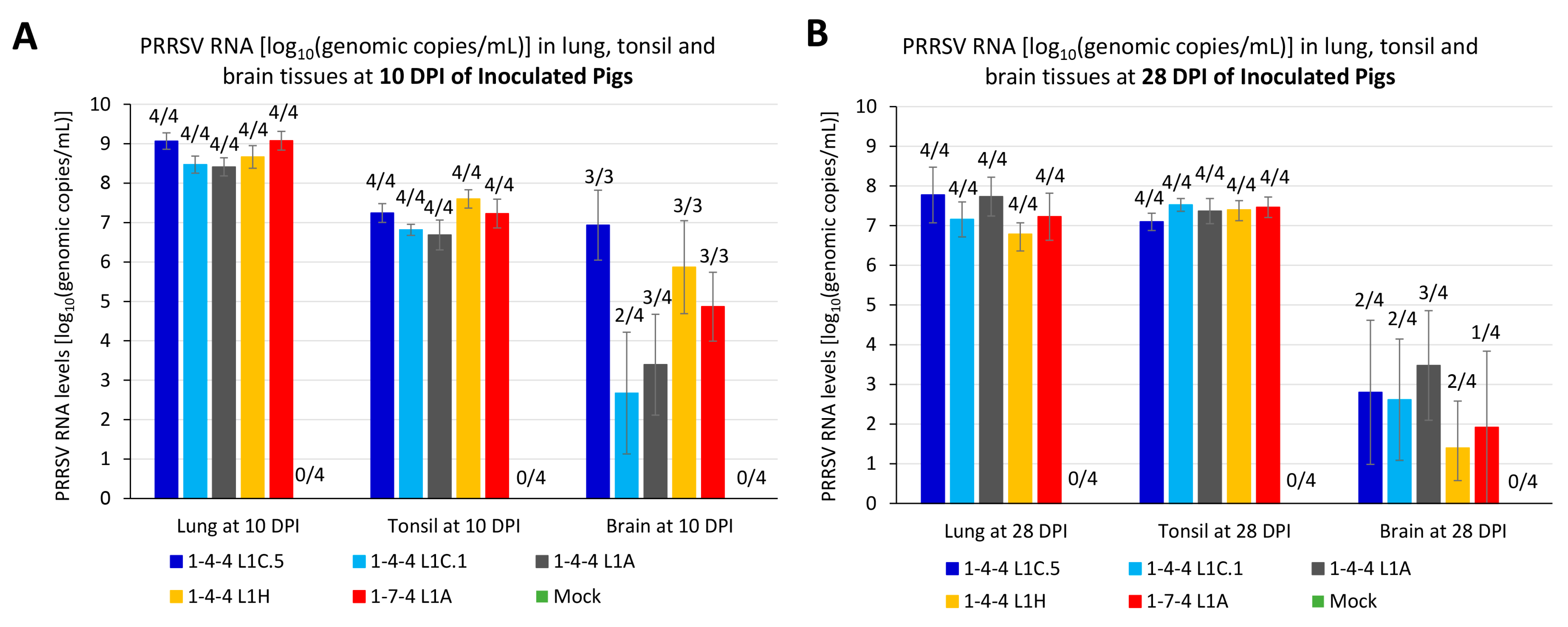
3.5. PRRSV RNA Load in Tissue Samples of Inoculated Pigs
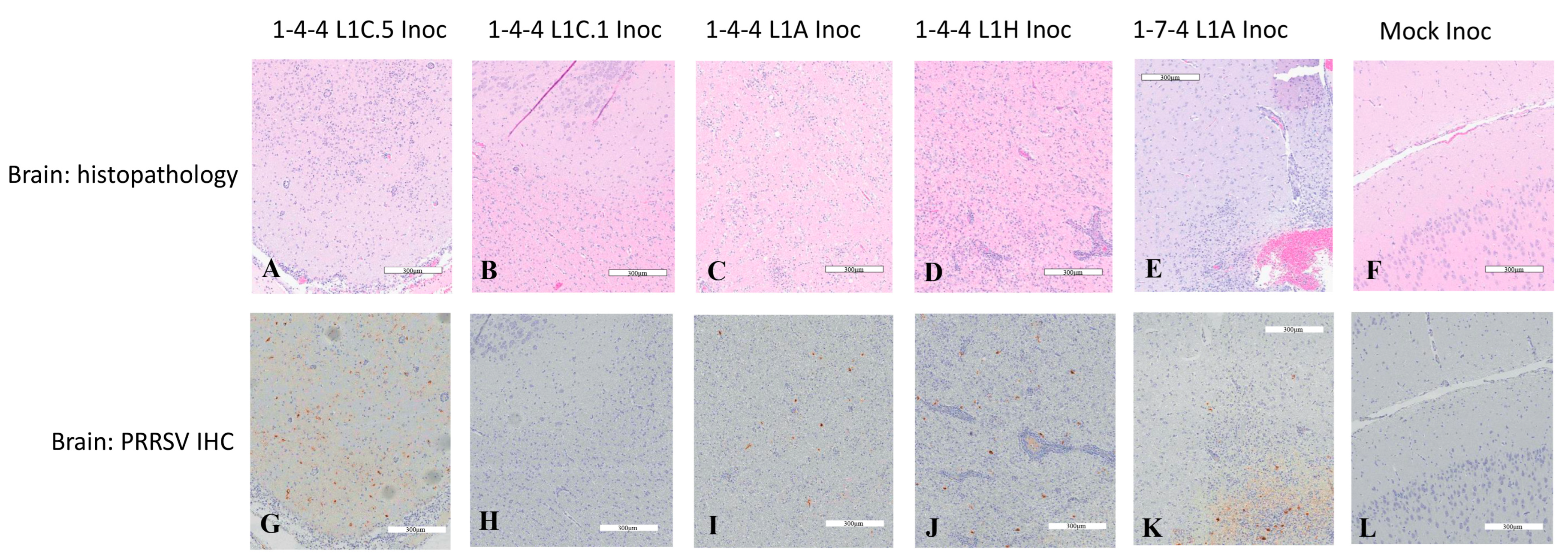
3.6. PRRSV Immunohistochemistry Staining in Brain Tissues
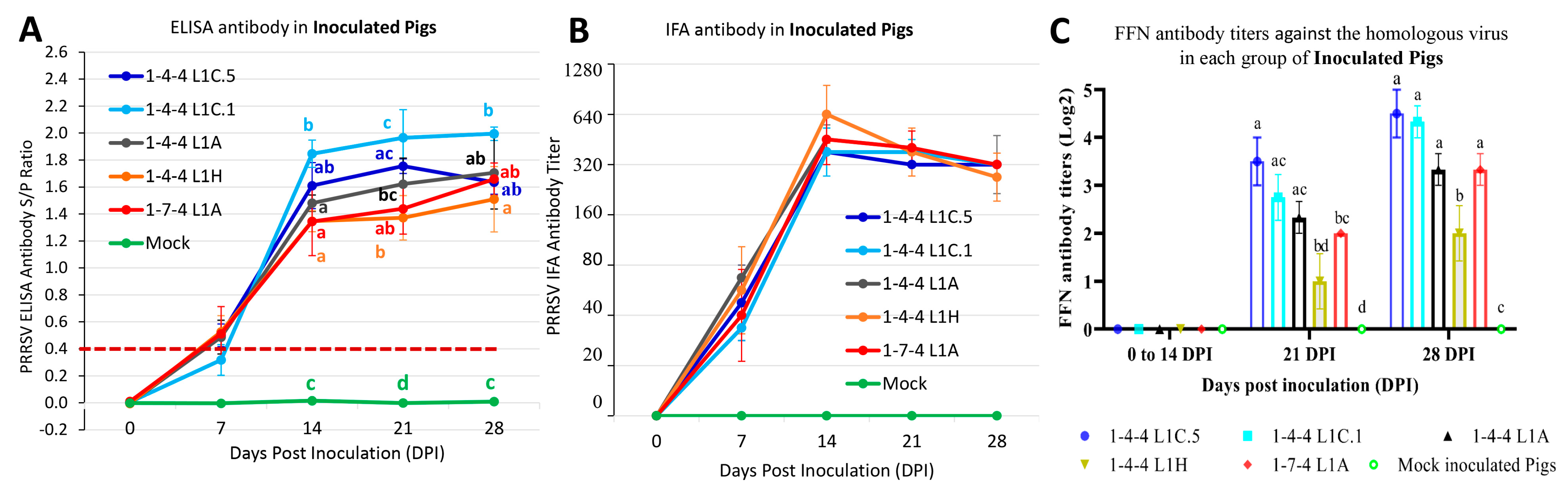
3.7. Antibody Responses in Inoculated Pigs
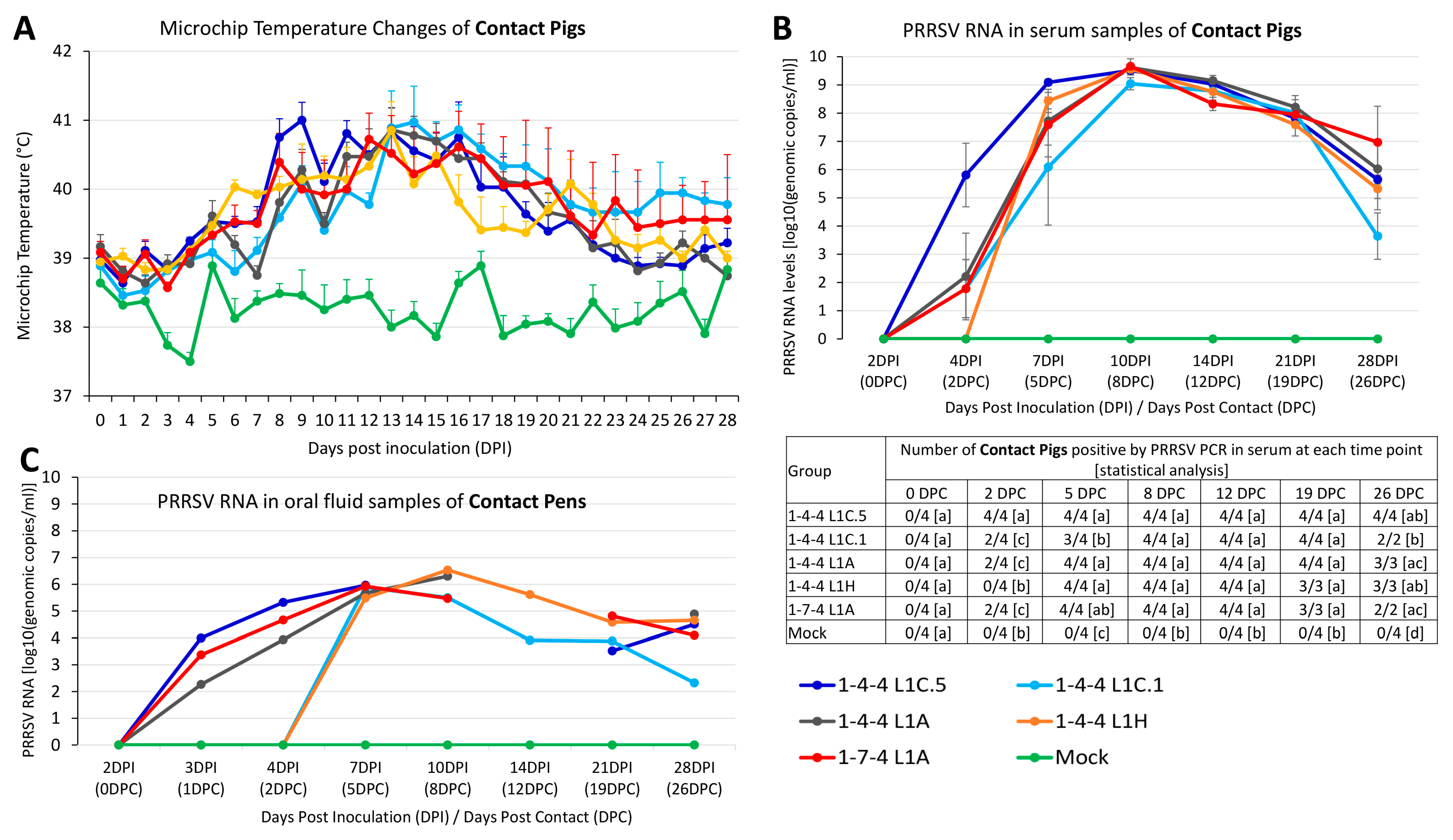
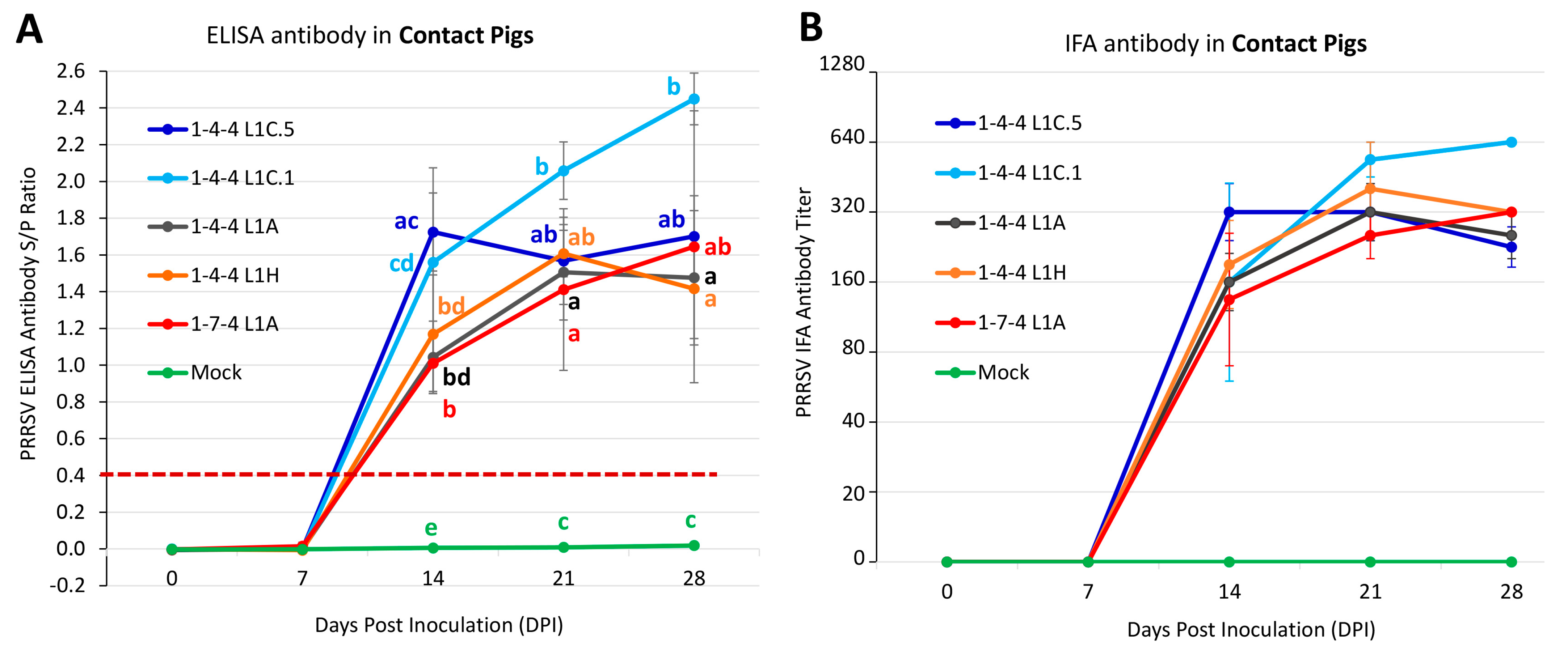
3.8. Outcomes in Contact Pigs
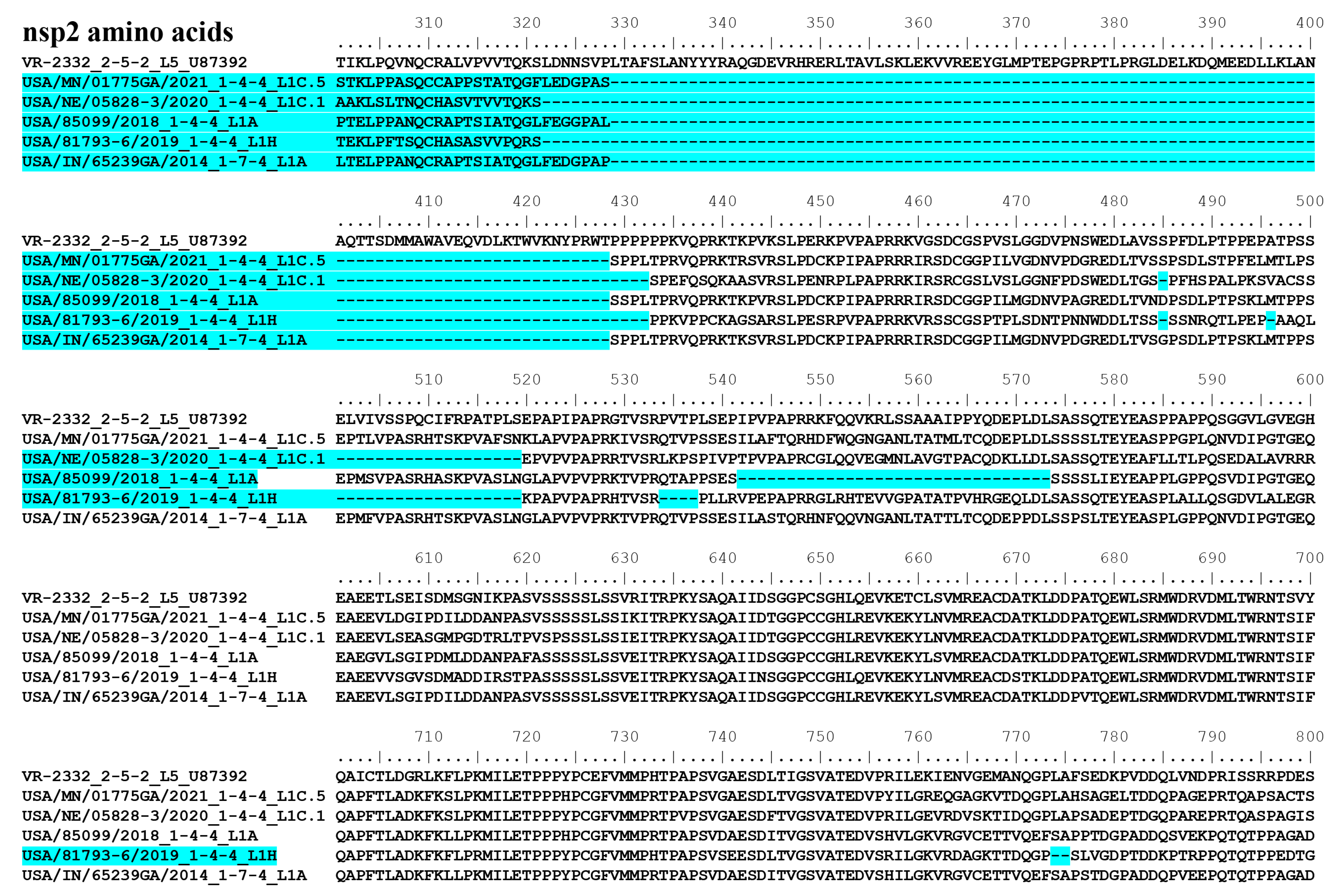
3.9. Whole Genome Sequence Comparison of PRRSV-2 Isolates Evaluated in This Study
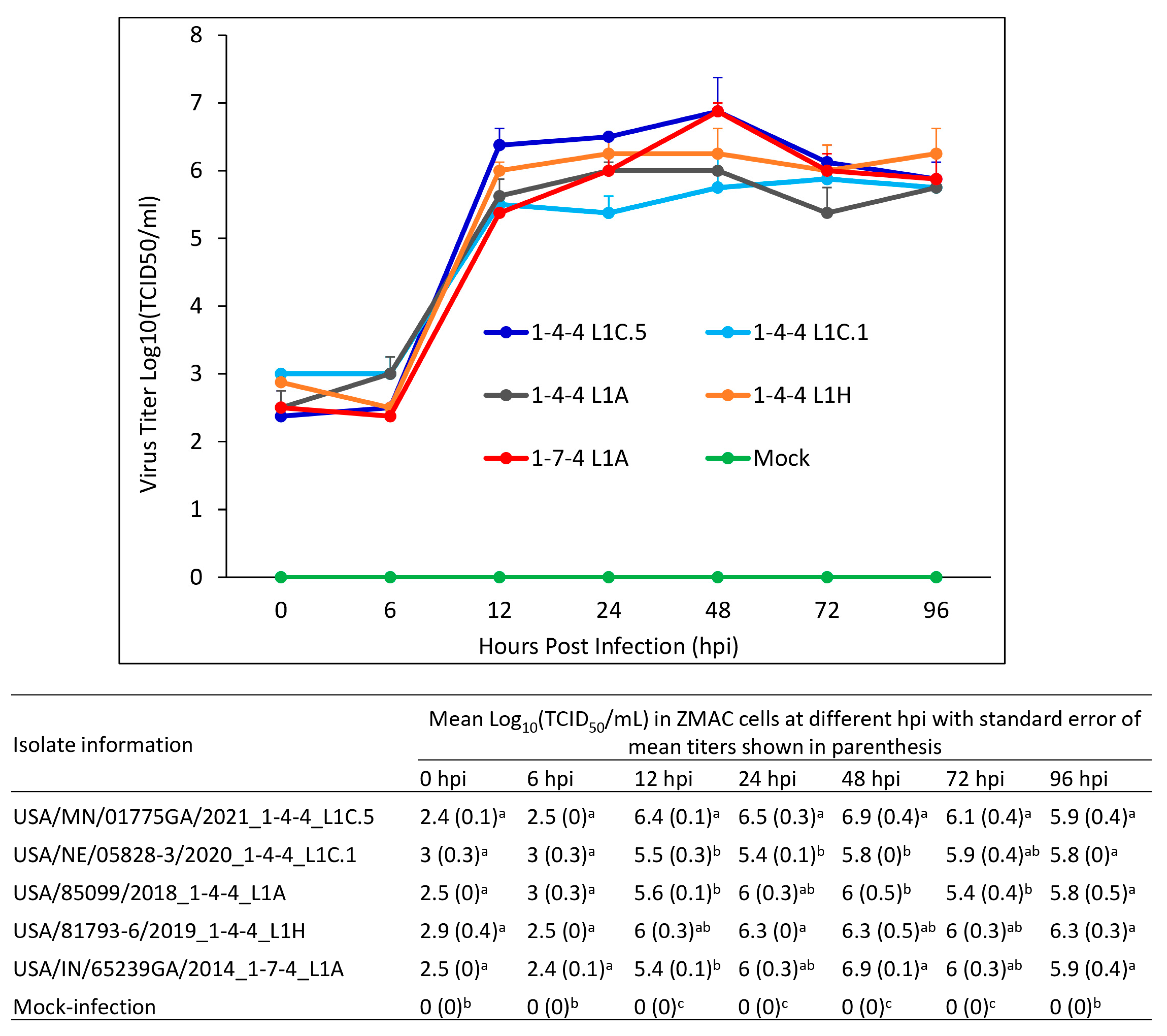
3.10. Growth Characteristics of Virus Isolates in ZMAC Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brinton, M.A.; Gulyaeva, A.A.; Balasuriya, U.B.R.; Dunowska, M.; Faaberg, K.S.; Goldberg, T.; Leung, F.C.C.; Nauwynck, H.J.; Snijder, E.J.; Stadejek, T.; et al. ICTV Virus Taxonomy Profile: Arteriviridae 2021. J. Gen. Virol. 2021, 102, 001632. [Google Scholar] [CrossRef]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef]
- Fang, Y.; Treffers, E.E.; Li, Y.; Tas, A.; Sun, Z.; van der Meer, Y.; de Ru, A.H.; van Veelen, P.A.; Atkins, J.F.; Snijder, E.J.; et al. Efficient -2 frameshifting by mammalian ribosomes to synthesize an additional arterivirus protein. Proc. Natl. Acad. Sci. USA 2012, 109, E2920–E2928. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, J.; Dee, S.; Holtkamp, D.J.; Murtaugh, M.; Stadejek, T.; Stevenson, G.; Torremorell, M.; Yang, H.; Zhang, J. Porcine reproductive and respiratory syndrome viruses (porcine arteriviruses). In Diseases of Swine, 11th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 685–708. [Google Scholar]
- Yim-im, W.; Anderson, T.K.; Paploski, I.; VanderWaal, K.; Gauger, P.; Kreuger, K.; Shi, M.; Main, R.; Zhang, J. Refining PRRSV-2 genetic classification based on global ORF5 sequences and investigation of their geographic distributions and temporal changes. Microbiol. Spectr. 2023, e0291623. [Google Scholar] [CrossRef]
- Wesley, R.D.; Mengeling, W.L.; Lager, K.M.; Clouser, D.F.; Landgraf, J.G.; Frey, M.L. Differentiation of a porcine reproductive and respiratory syndrome virus vaccine strain from North American field strains by restriction fragment length polymorphism analysis of ORF 5. J. Vet. Diagn. Investig. 1998, 10, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.H.; Chang, C.C.; Yoon, K.J. Instability of the restriction fragment length polymorphism pattern of open reading frame 5 of porcine reproductive and respiratory syndrome virus during sequential pig-to-pig passages. J. Clin. Microbiol. 2004, 42, 4462–4467. [Google Scholar] [CrossRef]
- Trevisan, G.; Sharma, A.; Gauger, P.; Harmon, K.M.; Zhang, J.; Main, R.; Zeller, M.; Linhares, L.C.M.; Linhares, D.C.L. PRRSV2 genetic diversity defined by RFLP patterns in the United States from 2007 to 2019. J. Vet. Diagn. Investig. 2021, 33, 920–931. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lam, T.T.; Hon, C.C.; Murtaugh, M.P.; Davies, P.R.; Hui, R.K.; Li, J.; Wong, L.T.; Yip, C.W.; Jiang, J.W.; et al. Phylogeny-based evolutionary, demographical, and geographical dissection of North American type 2 porcine reproductive and respiratory syndrome viruses. J. Virol. 2010, 84, 8700–8711. [Google Scholar] [CrossRef]
- Paploski, I.A.D.; Corzo, C.; Rovira, A.; Murtaugh, M.P.; Sanhueza, J.M.; Vilalta, C.; Schroeder, D.C.; VanderWaal, K. Temporal Dynamics of Co-circulating Lineages of Porcine Reproductive and Respiratory Syndrome Virus. Front. Microbiol. 2019, 10, 2486. [Google Scholar] [CrossRef]
- Paploski, I.; Pamornchainavakul, N.; Makau, D.N.; Rovira, A.; Corzo, C.; Schroeder, D.C.; Cheeran, M.; Doeschl-Wilson, A.; Kao, R.R.; Lycett, S.; et al. Phylogenetic Structure and Sequential Dominance of Sub-Lineages of PRRSV Type-2 Lineage 1 in the United States. Vaccines 2021, 9, 608. [Google Scholar] [CrossRef]
- Murtaugh, M.P.; Stadejek, T.; Abrahante, J.E.; Lam, T.T.Y.; Leung, F.C.C. The ever-expanding diversity of porcine reproductive and respiratory syndrome virus. Virus Res. 2010, 154, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479–480, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Hanada, K.; Suzuki, Y.; Nakane, T.; Hirose, O.; Gojobori, T. The Origin and Evolution of Porcine Reproductive and Respiratory Syndrome Viruses. Mol. Biol. Evol. 2005, 22, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Halbur, P.G.; Paul, P.S.; Frey, M.L.; Landgraf, J.; Eernisse, K.; Meng, X.J.; Lum, M.A.; Andrews, J.J.; Rathje, J.A. Comparison of the pathogenicity of two US porcine reproductive and respiratory syndrome virus isolates with that of the Lelystad virus. Vet. Pathol. 1995, 32, 648–660. [Google Scholar] [CrossRef]
- Johnson, W.; Roof, M.; Vaughn, E.; Christopher-Hennings, J.; Johnson, C.R.; Murtaugh, M.P. Pathogenic and humoral immune responses to porcine reproductive and respiratory syndrome virus (PRRSV) are related to viral load in acute infection. Vet. Immunol. Immunopathol. 2004, 102, 233–247. [Google Scholar] [CrossRef]
- Brockmeier, S.L.; Loving, C.L.; Vorwald, A.C.; Kehrli, M.E., Jr.; Baker, R.B.; Nicholson, T.L.; Lager, K.M.; Miller, L.C.; Faaberg, K.S. Genomic sequence and virulence comparison of four Type 2 porcine reproductive and respiratory syndrome virus strains. Virus Res. 2012, 169, 212–221. [Google Scholar] [CrossRef]
- Alkhamis, M.A.; Perez, A.M.; Murtaugh, M.P.; Wang, X.; Morrison, R.B. Applications of Bayesian Phylodynamic Methods in a Recent U.S. Porcine Reproductive and Respiratory Syndrome Virus Outbreak. Front. Microbiol. 2016, 7, 67. [Google Scholar] [CrossRef]
- van Geelen, A.G.M.; Anderson, T.K.; Lager, K.M.; Das, P.B.; Otis, N.J.; Montiel, N.A.; Miller, L.C.; Kulshreshtha, V.; Buckley, A.C.; Brockmeier, S.L.; et al. Porcine reproductive and respiratory disease virus: Evolution and recombination yields distinct ORF5 RFLP 1-7-4 viruses with individual pathogenicity. Virology 2018, 513, 168–179. [Google Scholar] [CrossRef]
- Tian, K.; Yu, X.; Zhao, T.; Feng, Y.; Cao, Z.; Wang, C.; Hu, Y.; Chen, X.; Hu, D.; Tian, X.; et al. Emergence of Fatal PRRSV Variants: Unparalleled Outbreaks of Atypical PRRS in China and Molecular Dissection of the Unique Hallmark. PLoS ONE 2007, 2, e526. [Google Scholar] [CrossRef]
- Jantafong, T.; Sangtong, P.; Saenglub, W.; Mungkundar, C.; Romlamduan, N.; Lekchareonsuk, C.; Lekcharoensuk, P. Genetic diversity of porcine reproductive and respiratory syndrome virus in Thailand and Southeast Asia from 2008 to 2013. Vet. Microbiol. 2015, 176, 229–238. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, X.X.; Li, R.; Qiao, S.; Zhang, G. The prevalent status and genetic diversity of porcine reproductive and respiratory syndrome virus in China: A molecular epidemiological perspective. Virol. J. 2018, 15, 2. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, G.; Yu, L.; Li, L.; Zhang, Y.; Zhou, Y.; Tong, W.; Liu, C.; Gao, F.; Tong, G. Genetic Diversity of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) From 1996 to 2017 in China. Front. Microbiol. 2020, 11, 618. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.C.; Chang, C.Y.; Huang, T.S.; Tsai, H.J.; Chang, C.; Wang, F.I.; Huang, Y.L. Molecular epidemiology of porcine reproductive and respiratory syndrome viruses isolated from 1991 to 2013 in Taiwan. Arch. Virol. 2015, 160, 2709–2718. [Google Scholar] [CrossRef] [PubMed]
- Do, H.Q.; Trinh, D.T.; Nguyen, T.L.; Vu, T.T.; Than, D.D.; Van Lo, T.; Yeom, M.; Song, D.; Choe, S.; An, D.J.; et al. Molecular evolution of type 2 porcine reproductive and respiratory syndrome viruses circulating in Vietnam from 2007 to 2015. BMC Vet. Res. 2016, 12, 256. [Google Scholar] [CrossRef]
- Lee, M.A.; Jayaramaiah, U.; You, S.H.; Shi, E.G.; Song, S.M.; Ju, L.; Kang, S.J.; Hyun, B.H.; Lee, H.S. Molecular Characterization of Porcine Reproductive and Respiratory Syndrome Virus in Korea from 2018 to 2022. Pathogens 2023, 12, 757. [Google Scholar] [CrossRef]
- Xu, H.; Xiang, L.; Tang, Y.D.; Li, C.; Zhao, J.; Gong, B.; Sun, Q.; Leng, C.; Peng, J.; Wang, Q.; et al. Genome-Wide Characterization of QYYZ-Like PRRSV During 2018-2021. Front. Vet. Sci. 2022, 9, 945381. [Google Scholar] [CrossRef]
- Yu, F.; Yan, Y.; Shi, M.; Liu, H.Z.; Zhang, H.L.; Yang, Y.B.; Huang, X.Y.; Gauger, P.C.; Zhang, J.; Zhang, Y.H.; et al. Phylogenetics, Genomic Recombination, and NSP2 Polymorphic Patterns of Porcine Reproductive and Respiratory Syndrome Virus in China and the United States in 2014-2018. J. Virol. 2020, 94, e01813–e01819. [Google Scholar] [CrossRef]
- Cui, X.; Xia, D.; Huang, X.; Sun, Y.; Shi, M.; Zhang, J.; Li, G.; Yang, Y.; Wang, H.; Cai, X.; et al. Analysis of Recombinant Characteristics Based on 949 PRRSV-2 Genomic Sequences Obtained from 1991 to 2021 Shows That Viral Multiplication Ability Contributes to Dominant Recombination. Microbiol. Spectr. 2022, 10, e0293422. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, Q.; Zheng, Y.; Sha, H.; Li, G.; Kong, W.; Huang, L.; Zhao, M. Genetic Variability and Recombination of the NSP2 Gene of PRRSV-2 Strains in China from 1996 to 2021. Vet. Sci. 2023, 10, 325. [Google Scholar] [CrossRef]
- Trevisan, G.; Li, G.; Moura, C.A.A.; Coleman, K.; Thomas, P.; Zhang, J.; Gauger, P.; Zeller, M.; Linhares, D. Complete Coding Genome Sequence of a Novel Porcine Reproductive and Respiratory Syndrome Virus 2 Restriction Fragment Length Polymorphism 1-4-4 Lineage 1C Variant Identified in Iowa, USA. Microbiol. Resour. Announc. 2021, 10, e0044821. [Google Scholar] [CrossRef]
- Kikuti, M.; Paploski, I.A.D.; Pamornchainavakul, N.; Picasso-Risso, C.; Schwartz, M.; Yeske, P.; Leuwerke, B.; Bruner, L.; Murray, D.; Roggow, B.D.; et al. Emergence of a New Lineage 1C Variant of Porcine Reproductive and Respiratory Syndrome Virus 2 in the United States. Front. Vet. Sci. 2021, 8, 752938. [Google Scholar] [CrossRef] [PubMed]
- Linhares, D.C.L.; Betlach, C.; Morrison, R.B. Effect of immunologic solutions on sows and gilts on time to stability, and production losses in breeding herds infected with 1-7-4 PRRSv. Prev. Vet. Med. 2017, 144, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Calzada-Nova, G.; Husmann, R.J.; Schnitzlein, W.M.; Zuckermann, F.A. Effect of the host cell line on the vaccine efficacy of an attenuated porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 2012, 148, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Kwang, J.; Yoon, I.J.; Joo, H.S.; Frey, M.L. Enhanced replication of porcine reproductive and respiratory syndrome (PRRS) virus in a homogeneous subpopulation of MA-104 cell line. Arch. Virol. 1993, 133, 477–483. [Google Scholar] [CrossRef]
- Yim-im, W.; Huang, H.; Park, J.; Wang, C.; Calzada, G.; Gauger, P.; Harmon, K.; Main, R.; Zhang, J. Comparison of ZMAC and MARC-145 Cell Lines for Improving Porcine Reproductive and Respiratory Syndrome Virus Isolation from Clinical Samples. J. Clin. Microbiol. 2021, 59, e01720–e01757. [Google Scholar] [CrossRef]
- Yim-im, W.; Huang, H.; Zheng, Y.; Li, G.; Rawal, G.; Gauger, P.; Krueger, K.; Main, R.; Zhang, J. Characterization of PRRSV in clinical samples and the corresponding cell culture isolates. Transbound. Emerg. Dis. 2022, 69, e3045–e3059. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Opriessnig, T.; Rawal, G.; McKeen, L.; Filippsen Favaro, P.; Halbur, P.G.; Gauger, P.C. Evaluation of the intranasal route for porcine reproductive and respiratory disease modified-live virus vaccination. Vaccine 2021, 39, 6852–6859. [Google Scholar] [CrossRef]
- Prickett, J.; Simer, R.; Christopher-Hennings, J.; Yoon, K.J.; Evans, R.B.; Zimmerman, J.J. Detection of Porcine reproductive and respiratory syndrome virus infection in porcine oral fluid samples: A longitudinal study under experimental conditions. J. Vet. Diagn. Investig. 2008, 20, 156–163. [Google Scholar] [CrossRef]
- Rawal, G.; Yim-Im, W.; Chamba, F.; Smith, C.; Okones, J.; Francisco, C.; Zhang, J. Development and validation of a reverse transcription real-time PCR assay for specific detection of PRRSGard vaccine-like virus. Transbound. Emerg. Dis. 2022, 69, 1212–1226. [Google Scholar] [CrossRef] [PubMed]
- Yoon, I.J.; Joo, H.S.; Christianson, W.T.; Kim, H.S.; Collins, J.E.; Morrison, R.B.; Dial, G.D. An indirect fluorescent antibody test for the detection of antibody to swine infertility and respiratory syndrome virus in swine sera. J. Vet. Diagn. Investig. 1992, 4, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yoon, K.J.; Li, Y.; Lee, J.H.; Zimmerman, J.J.; Frey, M.L.; Harmon, K.M.; Platt, K.B. Antigenic and genetic variations of the 15 kD nucleocapsid protein of porcine reproductive and respiratory syndrome virus isolates. Arch. Virol. 1999, 144, 525–546. [Google Scholar] [CrossRef] [PubMed]
- Halbur, P.G.; Paul, P.S.; Meng, X.J.; Lum, M.A.; Andrews, J.J.; Rathje, J.A. Comparative pathogenicity of nine US porcine reproductive and respiratory syndrome virus (PRRSV) isolates in a five-week-old cesarean-derived, colostrum-deprived pig model. J. Vet. Diagn. Investig. 1996, 8, 11–20. [Google Scholar] [CrossRef]
- Halbur, P.G.; Andrews, J.J.; Huffman, E.L.; Paul, P.S.; Meng, X.J.; Niyo, Y. Development of a streptavidin-biotin immunoperoxidase procedure for the detection of porcine reproductive and respiratory syndrome virus antigen in porcine lung. J. Vet. Diagn. Investig. 1994, 6, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zheng, Y.; Xia, X.Q.; Chen, Q.; Bade, S.A.; Yoon, K.J.; Harmon, K.M.; Gauger, P.C.; Main, R.G.; Li, G. High-throughput whole genome sequencing of Porcine reproductive and respiratory syndrome virus from cell culture materials and clinical specimens using next-generation sequencing technology. J. Vet. Diagn. Investig. 2017, 29, 41–50. [Google Scholar] [CrossRef]
- Gupta, R.; Brunak, S. Prediction of glycosylation across the human proteome and the correlation to protein function. Pac. Symp. Biocomput. 2002, 7, 310–322. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Yeske, P. Experiences with PRRS V1C. In Proceedings of the ISU James D. McKean Swine Disease Conference, Ames, IA, USA, 4–5 November 2021; pp. 34–38. [Google Scholar]
- Thomas, S.R.; Elkinton, J.S. Pathogenicity and virulence. J. Invertebr. Pathol. 2004, 85, 146–151. [Google Scholar] [CrossRef]
- Hibbs, J.; Young, N.S. Viruses, virulence and pathogenicity. Baillieres Clin. Haematol. 1995, 8, 1–23. [Google Scholar] [CrossRef]
- Ruedas-Torres, I.; Rodriguez-Gomez, I.M.; Sanchez-Carvajal, J.M.; Larenas-Munoz, F.; Pallares, F.J.; Carrasco, L.; Gomez-Laguna, J. The jigsaw of PRRSV virulence. Vet. Microbiol. 2021, 260, 109168. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; He, Y.; Xu, X.; Leng, X.; Li, S.; Wen, Y.; Wang, F.; Xia, M.; Cheng, S.; Wu, H. Pathological and immunological characteristics of piglets infected experimentally with a HP-PRRSV TJ strain. BMC Vet. Res. 2016, 12, 230. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Hao, X.F.; Tian, Z.J.; Tong, G.Z.; Yoo, D.; An, T.Q.; Zhou, T.; Li, G.X.; Qiu, H.J.; Wei, T.C.; et al. Highly virulent porcine reproductive and respiratory syndrome virus emerged in China. Transbound. Emerg. Dis. 2008, 55, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, X.; Bo, K.; Tang, B.; Yang, B.; Jiang, W.; Jiang, P. Emergence of a highly pathogenic porcine reproductive and respiratory syndrome virus in the Mid-Eastern region of China. Vet. J. 2007, 174, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Rossow, K.D.; Bautista, E.M.; Goyal, S.M.; Molitor, T.W.; Murtaugh, M.P.; Morrison, R.B.; Benfield, D.A.; Collins, J.E. Experimental porcine reproductive and respiratory syndrome virus infection in one-, four-, and 10-week-old pigs. J. Vet. Diagn. Investig. 1994, 6, 3–12. [Google Scholar] [CrossRef]
- Li, X.; Galliher-Beckley, A.; Wang, L.; Nietfeld, J.; Feng, W.; Shi, J. Comparison of Immune Responses in Pigs Infected with Chinese Highly Pathogenic PRRS Virus Strain HV and North American Strain NADC-20. Open Virol. J. 2017, 11, 73–82. [Google Scholar] [CrossRef]
- Canelli, E.; Catella, A.; Borghetti, P.; Ferrari, L.; Ogno, G.; De Angelis, E.; Corradi, A.; Passeri, B.; Bertani, V.; Sandri, G.; et al. Phenotypic characterization of a highly pathogenic Italian porcine reproductive and respiratory syndrome virus (PRRSV) type 1 subtype 1 isolate in experimentally infected pigs. Vet. Microbiol. 2017, 210, 124–133. [Google Scholar] [CrossRef]
- Frydas, I.S.; Trus, I.; Kvisgaard, L.K.; Bonckaert, C.; Reddy, V.R.; Li, Y.; Larsen, L.E.; Nauwynck, H.J. Different clinical, virological, serological and tissue tropism outcomes of two new and one old Belgian type 1 subtype 1 porcine reproductive and respiratory virus (PRRSV) isolates. Vet. Res. 2015, 46, 37. [Google Scholar] [CrossRef]
- Morgan, S.B.; Graham, S.P.; Salguero, F.J.; Sanchez Cordon, P.J.; Mokhtar, H.; Rebel, J.M.; Weesendorp, E.; Bodman-Smith, K.B.; Steinbach, F.; Frossard, J.P. Increased pathogenicity of European porcine reproductive and respiratory syndrome virus is associated with enhanced adaptive responses and viral clearance. Vet. Microbiol. 2013, 163, 13–22. [Google Scholar] [CrossRef]
- Allende, R.; Lewis, T.L.; Lu, Z.; Rock, D.L.; Kutish, G.F.; Ali, A.; Doster, A.R.; Osorio, F.A. North American and European porcine reproductive and respiratory syndrome viruses differ in non-structural protein coding regions. J. Gen. Virol. 1999, 80, 307–315. [Google Scholar] [CrossRef]
- Han, J.; Rutherford, M.S.; Faaberg, K.S. The porcine reproductive and respiratory syndrome virus nsp2 cysteine protease domain possesses both trans- and cis-cleavage activities. J. Virol. 2009, 83, 9449–9463. [Google Scholar] [CrossRef] [PubMed]
- Snijder, E.J.; Wassenaar, A.L.M.; Spaan, W.J.M.; Gorbalenya, A.E. The arterivirus Nsp2 protease: An unusual cysteine protease with primary structure similarities to both papain-like and chymotrypsin-like proteases. J. Biol. Chem. 1995, 270, 16671–16676. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Aravind, L.; Koonin, E.V. A novel superfamily of predicted cysteine proteases from eukaryotes, viruses and Chlamydia pneumoniae. Trends Biochem. Sci. 2000, 25, 50–52. [Google Scholar] [CrossRef]
- Sun, Z.; Chen, Z.; Lawson, S.R.; Fang, Y. The Cysteine Protease Domain of Porcine Reproductive and Respiratory Syndrome Virus Nonstructural Protein 2 Possesses Deubiquitinating and Interferon Antagonism Functions. J. Virol. 2010, 84, 7832–7846. [Google Scholar] [CrossRef] [PubMed]
- Faaberg, K.S.; Hocker, J.D.; Erdman, M.M.; Harris, D.L.; Nelson, E.A.; Torremorell, M.; Plagemann, P.G. Neutralizing antibody responses of pigs infected with natural GP5 N-glycan mutants of porcine reproductive and respiratory syndrome virus. Viral Immunol. 2006, 19, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Paploski, I.A.D.; Makau, D.N.; Pamornchainavakul, N.; Baker, J.P.; Schroeder, D.; Rovira, A.; VanderWaal, K. Potential Novel N-Glycosylation Patterns Associated with the Emergence of New Genetic Variants of PRRSV-2 in the U.S. Vaccines 2022, 10, 2021. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate ID | Passage No. | RFLP and Lineage | PRRSV PCR CT | TCID50/mL in ZMAC Cells | TCID50/mL in MARC-145 Cells |
|---|---|---|---|---|---|
| USA/MN/01775GA/2021 | P4 | 1-4-4 L1C.5 (L1C variant) | 16.5 | 106.75 | 104.25 |
| USA/NE/05828-3/2020 | P4 | 1-4-4 L1C.1 | 17.1 | 106 | 103.75 |
| USA/85099/2018 | P4 | 1-4-4 L1A | 15.1 | 106.25 | 103.75 |
| USA/81793-6/2019 | P4 | 1-4-4 L1H | 21.3 | 106.5 | 103.5 |
| USA/IN/65239GA/2014 | P4 | 1-7-4 L1A | 15.3 | 106.25 | 104.75 |
| Inoculation Group | Mortality | Occurrence Day |
|---|---|---|
| 1-4-4 L1C.5 (L1C variant) | 6/8 | 1 (8 DPI); 1 (9 DPI); 2 (10 DPI); 2 (14 DPI) |
| 1-4-4 L1C.1 | 1/8 | 1 (20 DPI) |
| 1-4-4 L1A | 1/8 | 1 (12 DPI) |
| 1-4-4 L1H | 2/8 | 2 (9 DPI) |
| 1-7-4 L1A | 2/8 | 1 (9 DPI); 1 (15 DPI) |
| Mock-inoculation | 0/8 |
| Group | Pig ID | Euthanasia Date | Brain | Ataxia or Could Not Stand Up | |||
|---|---|---|---|---|---|---|---|
| PRRSV PCR CT in Brain | PRRSV IHC Score in Brain | Average Microscopic Lesion Score * | Bacterial Culture | ||||
| 1-4-4 L1C.5, Inoculated Pigs | 6 | 8 DPI | × | × | × | × | |
| 11 | 10 DPI | 27.9 | 1 | 1 | No growth | ||
| 16 | 9 DPI | 17.5 | 3 | 1.75 | Low Strep equisimilis | ||
| 63 | 10 DPI | 22.3 | 0 | 0.75 | No growth | ||
| 20 | 14 DPI | 20.2 | 1 | 1 | No growth | ||
| 69 | 14 DPI | 34.0 | 0 | 0 | No growth | ||
| 64 | 28 DPI | ≥40 | 0 | 0.5 | No growth | ||
| 71 | 28 DPI | ≥40 | 0 | 0.5 | No growth | ||
| 1-4-4 L1C.5, Contact Pigs | 22 | 28 DPI | 36.2 | 0 | 0 | No growth | |
| 44 | 28 DPI | 34.3 | 0 | 0 | No growth | ||
| 46 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| 55 | 28 DPI | ≥40 | 0 | 0.25 | No growth | ||
| 1-4-4 L1C.1, Inoculated Pigs | 7 | 10 DPI | ≥40 | 0 | 0.25 | No growth | |
| 23 | 10 DPI | ≥40 | 0 | 0.75 | No growth | ||
| 70 | 10 DPI | 28.0 | 0 | 0.5 | No growth | ||
| 72 | 10 DPI | 28.0 | 0 | 0.25 | No growth | ||
| 12 | 20 DPI | 26.3 | 0 | 0.5 | No growth | ||
| 19 | 28 DPI | ≥40 | 0 | 0.25 | No growth | ||
| 66 | 28 DPI | 30.5 | 0 | 0 | No growth | ||
| 68 | 28 DPI | ≥40 | 0 | 0.25 | No growth | ||
| 1-4-4 L1C.1, Contact Pigs | 30 | 28 DPI | 30.3 | 0 | 0.5 | No growth | |
| 35 | 28 DPI | 30.3 | 0 | 0.25 | No growth | ||
| 43 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| 47 | 28 DPI | ≥40 | 0 | 0.25 | No growth | ||
| 1-4-4 L1A, Inoculated Pigs | 1 | 10 DPI | ≥40 | 0 | 0.5 | Low Strep suis | |
| 13 | 10 DPI | 33.3 | 0 | 0 | No growth | ||
| 53 | 10 DPI | 25.0 | 1 | 1 | No growth | ||
| 65 | 10 DPI | 33.9 | 1 | 0.5 | No growth | ||
| 5 | 12 DPI | 24.1 | 1 | 0.75 | No growth | Yes | |
| 8 | 28 DPI | 30.4 | 0 | 0 | No growth | ||
| 58 | 28 DPI | 36.7 | 0 | 0 | No growth | ||
| 59 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| 1-4-4 L1A, Contact Pigs | 38 | 17 DPI | 22.3 | 2 | 1.5 | No growth | Yes |
| 21 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| 39 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| 40 | 28 DPI | 29.8 | 0 | 0.25 | No growth | ||
| 1-4-4 L1H, Inoculated Pigs | 17 | 10 DPI | 22.9 | 2 | 1.25 | No growth | |
| 33 | 9 DPI | 21.5 | 2 | 3 | No growth | Yes | |
| 45 | 10 DPI | 34.2 | 0 | 0.25 | No growth | ||
| 62 | 9 DPI | × | × | × | × | ||
| 10 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| 18 | 28 DPI | 37.9 | 0 | 0.25 | No growth | ||
| 49 | 28 DPI | 35.3 | 0 | 0.5 | No growth | ||
| 57 | 28 DPI | ≥40 | 0 | 0.5 | No growth | ||
| 1-4-4 L1H, Contact Pigs | 37 | 12 DPI | 20.3 | 3 | 2.25 | No growth | Yes |
| 34 | 28 DPI | ≥40 | 0 | 0.25 | No growth | ||
| 41 | 28 DPI | 34.8 | 0 | 0.75 | No growth | ||
| 42 | 28 DPI | 31.9 | 0 | 0.25 | No growth | ||
| 1-7-4 L1A, Inoculated Pigs | 26 | 9 DPI | × | × | × | × | |
| 27 | 10 DPI | 24.9 | 1 | 1.5 | Few Glaesserella parasuis | ||
| 60 | 10 DPI | 35.1 | 0 | 0.5 | No growth | ||
| 61 | 10 DPI | 28.8 | 0 | 0 | No growth | ||
| 4 | 15 DPI | 20.1 | 1 | 1.5 | No growth | ||
| 2 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| 52 | 28 DPI | ≥40 | 0 | 0.25 | No growth | ||
| 67 | 28 DPI | ≥40 | 0 | 0.25 | No growth | ||
| 1-7-4 L1A, Contact Pigs | 36 | 12 DPI | 20.8 | 1 | 1.25 | No growth | |
| 50 | 15 DPI | 30.1 | 0 | 0.25 | No growth | ||
| 48 | 28 DPI | ≥40 | 0 | 0.25 | No growth | ||
| 51 | 28 DPI | 25.0 | 0 | 0.25 | No growth | ||
| Mock-inoculated Pigs | 9 | 10 DPI | ≥40 | 0 | 0 | No growth | |
| 15 | 10 DPI | ≥40 | 0 | 0 | No growth | ||
| 31 | 10 DPI | ≥40 | 0 | 0 | No growth | ||
| 56 | 10 DPI | ≥40 | 0 | 0 | No growth | ||
| 3 | 28 DPI | ≥40 | 0 | 0 | Low Strep suis | ||
| 14 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| 32 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| 54 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| Mock-contact Pigs | 24 | 28 DPI | ≥40 | 0 | 0 | No growth | |
| 25 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| 28 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| 29 | 28 DPI | ≥40 | 0 | 0 | No growth | ||
| ORF | Protein | Nucleotide Length (Protein Length) | |||||
|---|---|---|---|---|---|---|---|
| 2-5-2 L5A (VR-2332) | 1-4-4 L1C.5 (USA/MN/01775 GA/2021) | 1-4-4 L1C.1 (USA/NE/05828-3/2020) | 1-4-4 L1A (USA/85099/2018) | 1-4-4 L1H (USA/81793-6/2019 | 1-7-4 L1A (USA/IN/65239 GA/2014 | ||
| 5′ UTR | N.A. | 189 nt | 187 nt | 190 nt | 188 nt | 190 nt | 188 nt |
| ORF1a | pp1a | 7512 nt (2503 aa) | 7212 nt (2403 aa) | 7119 nt (2372 aa) | 7116 nt (2371 aa) | 7098 nt (2365 aa) | 7212 nt (2403 aa) |
| ORF1b | N.A. | all 4377 nt | |||||
| ORF1a/b | pp1ab | 11,883 nt (3960 aa) | 11,583 nt (3860 aa) | 11,490 nt (3829 aa) | 11,487 nt (3828 aa) | 11,469 nt (3822 aa) | 11,583 nt (3860 aa) |
| nsp1 | all 1149 nt (383 aa) | ||||||
| nsp2 | 3588 nt (1196 aa) | 3288 nt (1096 aa) | 3195 nt (1065 aa) | 3192 nt (1064 aa) | 3174 nt (1058 aa) | 3288 nt (1096 aa) | |
| nsp3 | all 690 nt (230 aa) | ||||||
| nsp4 | all 612 nt (204 aa) | ||||||
| nsp5 | all 510 nt (170 aa) | ||||||
| nsp6 | all 48 nt (16 aa) | ||||||
| nsp7 | all 777 nt (259 aa) | ||||||
| nsp8 | all 135 nt (45 aa) | ||||||
| nsp9 | all 2055 nt (685 aa) | ||||||
| nsp10 | all 1323 nt (441 aa) | ||||||
| nsp11 | all 669 nt (223 aa) | ||||||
| nsp12 | all 459 nt (153 aa) | ||||||
| ORF2a | GP2 | all 771 nt (256 aa) | |||||
| ORF2b | E | all 222 nt (73 aa) | |||||
| ORF3 | GP3 | all 765 nt (254 aa) | |||||
| ORF4 | GP4 | all 537 nt (178 aa) | |||||
| ORF5a | ORF5a | 156 nt (51 aa) | 141 nt (46 aa) | 141 nt (46 aa) | 156 nt (51 aa) | 141 nt (46 aa) | 141 nt (46 aa) |
| ORF5 | GP5 | all 603 nt (200 aa) | |||||
| ORF6 | M | all 525 nt (174 aa) | |||||
| ORF7 | N | all 372 nt (123 aa) | |||||
| 3′ UTR | N.A. | 151 nt | 151 nt | 141 nt | 151 nt | 151 nt | 151 nt |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rawal, G.; Almeida, M.N.; Gauger, P.C.; Zimmerman, J.J.; Ye, F.; Rademacher, C.J.; Armenta Leyva, B.; Munguia-Ramirez, B.; Tarasiuk, G.; Schumacher, L.L.; et al. In Vivo and In Vitro Characterization of the Recently Emergent PRRSV 1-4-4 L1C Variant (L1C.5) in Comparison with Other PRRSV-2 Lineage 1 Isolates. Viruses 2023, 15, 2233. https://doi.org/10.3390/v15112233
Rawal G, Almeida MN, Gauger PC, Zimmerman JJ, Ye F, Rademacher CJ, Armenta Leyva B, Munguia-Ramirez B, Tarasiuk G, Schumacher LL, et al. In Vivo and In Vitro Characterization of the Recently Emergent PRRSV 1-4-4 L1C Variant (L1C.5) in Comparison with Other PRRSV-2 Lineage 1 Isolates. Viruses. 2023; 15(11):2233. https://doi.org/10.3390/v15112233
Chicago/Turabian StyleRawal, Gaurav, Marcelo N. Almeida, Phillip C. Gauger, Jeffrey J. Zimmerman, Fangshu Ye, Christopher J. Rademacher, Betsy Armenta Leyva, Berenice Munguia-Ramirez, Grzegorz Tarasiuk, Loni L. Schumacher, and et al. 2023. "In Vivo and In Vitro Characterization of the Recently Emergent PRRSV 1-4-4 L1C Variant (L1C.5) in Comparison with Other PRRSV-2 Lineage 1 Isolates" Viruses 15, no. 11: 2233. https://doi.org/10.3390/v15112233