


Horizontal and Vertical Transmission of a Mycovirus Closely Related to the Partitivirus RhsV717 That Confers Hypovirulence in Rhizoctonia solani
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction and Detection of dsRNA
2.2. cDNA Synthesis, Molecular Cloning, and Sequencing
2.3. Sequence Analysis
2.4. Virus Particle Extraction and Detection
2.5. Horizontal and Vertical Transmission of RsPV-BS5
2.6. Virus Curing
2.7. Determination of Growth Rate
2.8. Virulence Assay
2.9. RNA-Seq and Data Analysis
3. Results
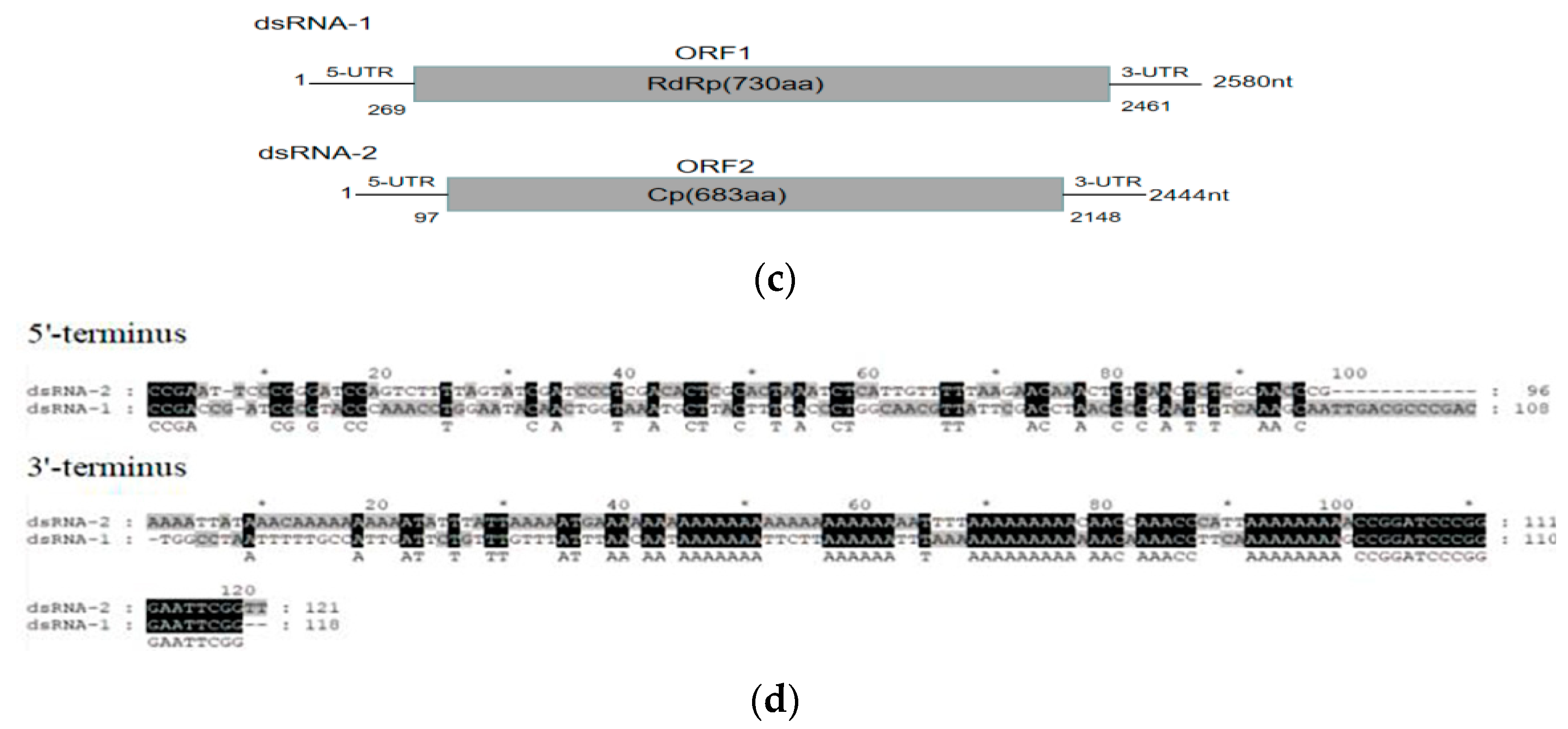
3.1. Nucleotide Sequences of RsPV-BS5 Genomic dsRNAs
3.2. Amino Acid Sequence and Phylogenetic Analysis of Partitivirus RsPV-BS5
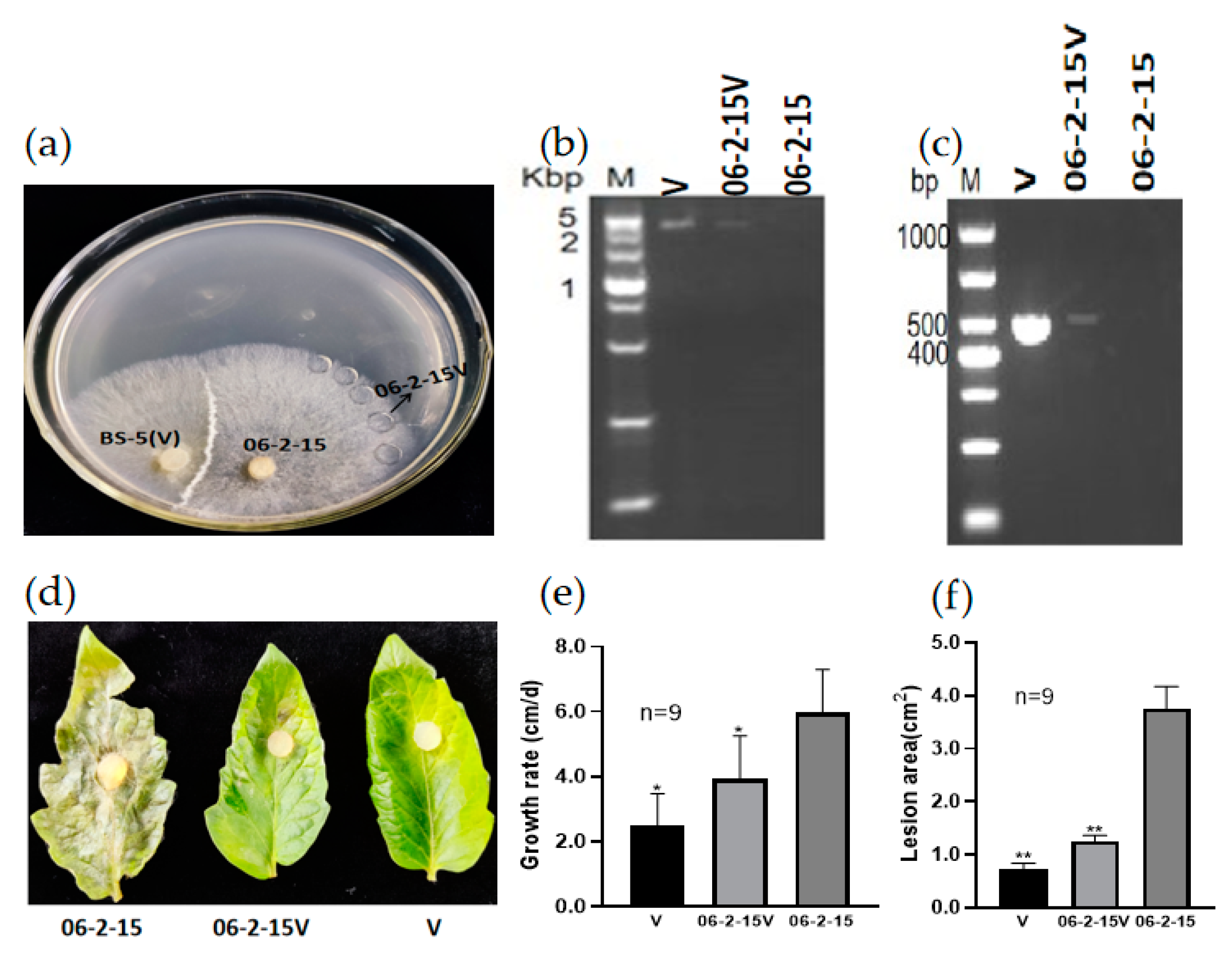
3.3. RsPV-BS5 Can Horizontally Transfer to the Virus-Free Strain 06-2-15
3.4. Vertical Transmission of RsPV-BS5
3.5. RsPV-BS5 Curing
3.6. RNA-Seq Analysis of R. solani AG-3 PT Response to RsPV-BS5 Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- García, V.G.; Onco, M.P.; Susan, V.R. Biology and systematics of the form genus Rhizoctonia. Span. J. Agric. Res. 2006, 4, 55–79. [Google Scholar] [CrossRef]
- Yang, Y.G.; Zhao, C.; Guo, Z.J.; Wu, X.H. Anastomosis group and pathogenicity of Rhizoctonia solani associated with stem canker and black scurf of potato in China. Eur. J. Plant Pathol. 2015, 143, 99–111. [Google Scholar] [CrossRef]
- Johnk, J.S.; Jones, R.; Shew, H.; Carling, D. Characterization of populations of Rhizoctonia solani AG-3 from potato and tobacco. Phytopathology 1993, 83, 854–858. [Google Scholar] [CrossRef]
- Kuninaga, S.; Carling, D.E.; Takeuchi, T.; Yokosawa, R. Comparison of rDNA-ITS sequences between potato and tobacco strains in Rhizoctonia solani AG-3. J. Gen. Plant Pathol. 2000, 66, 2–11. [Google Scholar] [CrossRef]
- Misawa, T.; Kuninaga, S. The first report of tomato foot rot caused by Rhizoctonia solani AG-3 PT and AG-2-Nt and its host range and molecular characterization. J. Gen. Plant Pathol. 2010, 76, 310–319. [Google Scholar] [CrossRef]
- Shew, H.D.; Main, C.E. Infection and development of target spot of flue-cured tobacco caused by Thanatephorus cucumeris. Plant Dis. 1991, 74, 1009–1013. [Google Scholar] [CrossRef]
- Meyer, J.C.; Wyk, R.J.V.; Phillips, A.J.R. Rhizoctonia leaf spot of tobacco in South Africa. Plant Pathol. 2010, 39, 206–207. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, Y.; Zhao, X.; An, M.; Chen, J.G. Study on the Taqman Real-Time PCR to the detection and quantification of Rhizoctonia solani AG-3 of tobacco target spot. Adv. Mater. Res. 2014, 3384, 1010–1012. [Google Scholar] [CrossRef]
- Xu, M.; Hao, K.; Yang, J.; Wang, F.; Xiao, Z.; Li, W. First report of Rhizoctonia solani AG-3 causing tobacco target spot in Yunnan, China. Plant Dis. 2018, 102, 2038. [Google Scholar] [CrossRef]
- Castano, R.; Borrero, C.; Trillas, M.; Avilés, M. Selection of biological control agents against tomato Fusarium wilt and evaluation in greenhouse conditions of two selected agents in three growing media. BioControl 2013, 58, 105–116. [Google Scholar] [CrossRef]
- Hollings, M. Viruses associated with a die-back disease of cultivated mushroom. Nature 1962, 196, 962–965. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, C.; Zhang, M.; Yang, M.; Zhou, E. Diversity of dsRNA viruses infecting rice sheath blight fungus Rhizoctonia solani AG-1 IA. Rice Sci. 2018, 25, 57–60. [Google Scholar] [CrossRef]
- Li, Y.; Li, S.; Zhao, Y.; Zhou, T.; Wu, X.; Zhao, C. Six novel mycoviruses containing positive single-stranded RNA and double-stranded RNA genomes co-Infect a single strain of the Rhizoctonia solani AG-3 PT. Viruses 2022, 14, 813. [Google Scholar] [CrossRef]
- Abdoulaye, A.H.; Foda, M.F.; Kotta-Loizou, I. Viruses infecting the plant pathogenic fungus Rhizoctonia solani. Viruses 2019, 11, 1113. [Google Scholar] [CrossRef]
- Picarelli, M.A.S.C.; Forgia, M.; Rivas, E.B.; Nerva, L.; Chiapello, M.; Turina, M. Extreme diversity of mycoviruses present in isolates of Rhizoctonia solani AG2-2 LP from Zoysia japonica from Brazil. Front. Cell Infect. Microbiol. 2019, 9, 244. [Google Scholar] [CrossRef]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef]
- Nibert, M.L.; Woods, K.M.; Upton, S.J.; Ghabrial, S.A. Cryspovirus: A new genus of protozoan viruses in the family Partitiviridae. Arch. Virol. 2009, 154, 1959–1965. [Google Scholar] [CrossRef]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M. ICTV virus taxonomy profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef]
- Petrzik, K. Evolutionary forces at work in partitiviruses. Virus Genes 2019, 55, 563–573. [Google Scholar] [CrossRef]
- Nuss, D.L. Biological control of chestnut blight: An example of virus-mediated attenuation of fungal pathogenesis. Microbiol. Rev. 1992, 56, 561–576. [Google Scholar] [CrossRef]
- Yu, X.; Li, B.; Fu, Y.; Jiang, D.; Ghabrial, S.A.; Li, G. A geminivirus-related DNA mycovirus that confers hypovirulence to a plant pathogenic fungus. Proc. Natl. Acad. Sci. USA 2010, 107, 8387–8392. [Google Scholar] [CrossRef]
- Yang, D.; Wu, M.; Zhang, J.; Chen, W.; Li, G.; Yang, L. Sclerotinia minor endornavirus 1, a novel pathogenicity debilitation-associated mycovirus with a wide spectrum of horizontal transmissibility. Viruses 2018, 10, 589. [Google Scholar] [CrossRef]
- Tavantzis, S.M.; Bandy, B.P. Properties of a Mycovirus from Rhizoctonia solani and Its Virion-associated RNA Polymerase. J. Gen. Virol. 1988, 69, 1465–1477. [Google Scholar] [CrossRef]
- Strauss, E.E.; Lakshman, D.K.; Tavantzis, S.M. Molecular characterization of the genome of a partitivirus from the basidiomycete Rhizoctonia solani. J. Gen. Virol. 2000, 81, 549–555. [Google Scholar] [CrossRef]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal-plant interface. Nat. Rev. Microbiol 2005, 3, 632–642. [Google Scholar] [CrossRef]
- Zheng, L.; Zhang, M.; Chen, Q.; Zhu, M.; Zhou, E. A novel mycovirus closely related to viruses in the genus Alphapartitivirus confers hypovirulence in the phytopathogenic fungus Rhizoctonia solani. Virology 2014, 456–457, 220–226. [Google Scholar] [CrossRef]
- Xiao, X.; Cheng, J.; Tang, J.; Fu, Y.; Jiang, D.; Baker, T.S. A novel partitivirus that confers hypovirulence on plant pathogenic fungi. J. Virol. 2014, 88, 10120–10133. [Google Scholar] [CrossRef]
- Zhang, M.; He, Z.; Huang, X.; Shu, C.; Zhou, E. Genome Organizations and Functional Analyses of a Novel Gammapartitivirus from Rhizoctonia solani AG-1 IA Strain D122. Viruses 2021, 13, 2254. [Google Scholar] [CrossRef]
- Valverde, R.A.; Gutierrez, D.L. Transmission of a dsRNA in bell pepper and evidence that it consists of the genome of an endornavirus. Virus Genes 2007, 35, 399–403. [Google Scholar] [CrossRef]
- Gonzalez, M.; Pujol, M.; Metraux, J.P. leaf spot and root rot caused by Rhizoctonia solani Kühn. Mol. Plant Pathol. 2011, 12, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.; Pujol, M.; Metraux, J.P.; Gonzalez-Garcia, V.; Bolton, M.D.; Borrás-Hidalgo, O. Characterization of south African Cryphonectria cubensis isolates infected with a C. parasitica Hypovirus. Phytopathology 2001, 91, 628–632. [Google Scholar]
- Sneh, B.J.; Abaji-Hare, S.; Neate, S. Rhizoctonia species: Taxonomy, molecular biology, ecology, pathology and disease control. In Book Meeting; Springer: Berlin/Heidelberg, Germany, 1996; pp. 341–349. [Google Scholar]
- Nerva, L.; Ciuffo, M.; Vallino, M.; Margaria, P.; Varese, G.C.; Gnavi, G.; Turina, M. Multiple approaches for the detection and characterization of viral and plasmid symbionts from a collection of marine fungi. Virus Res. 2016, 219, 22–38. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, Y.; Xu, P.; Zhang, L.; Chen, Z.; Niu, Y.; Yang, G.; Mo, X. A novel alphapartitivirus from binucleate Rhizoctonia fumigata AG-Ba isolate C-314 Baishi. Arch. Virol. 2022, 167, 255–259. [Google Scholar] [CrossRef]
- Bar-Joseph, M.; Rosner, A.; Moscovitz, M.; Hull, R. A simple procedure for the extraction of double-stranded RNA from virus-infected plants. J. Virol. Methods 1983, 6, 1–8. [Google Scholar] [CrossRef]
- Sun, A.; Luo, L.; Zhao, Z.; Kang, L.; Li, C.; Dong, W.; Yang, G. Complete genome sequence of a novel fusarivirus from Rhizoctonia solani AG-3 PT strain 3P-2-2. Arch. Virol. 2023, 168, 75. [Google Scholar] [CrossRef]
- Gong, W.; Liu, H.; Zhu, X.; Zhao, S.; Cheng, J.; Zhu, H.; Zhong, J.; Zhou, Q. Molecular characterization of a novel fusarivirus infecting the plant-pathogenic fungus Alternaria solani. Arch. Virol. 2021, 166, 2063–2067. [Google Scholar] [CrossRef]
- He, Y.; Zou, Q.; Li, S.; Zhu, H.; Hong, N.; Wang, G.; Wang, L. Molecular characterization of a new fusarivirus infecting Botryosphaeria dothidea, the causal agent of pear ring rot disease. Arch. Virol. 2022, 167, 1893–1897. [Google Scholar] [CrossRef]
- Potgieter, A.C.; Page, N.A.; Liebenberg, J.; Wright, I.M.; Landt, O.; van Dijk, A.A. Improved strategies for sequence- independent amplification and sequencing of viral double-stranded RNA genomes. J. Gen. Virol. 2009, 90, 1423–1432. [Google Scholar] [CrossRef]
- Li, Y.; Xu, P.; Zhang, L.; Xia, Z.; Qin, X.; Yang, G.; Mo, X. Molecular characterization of a novel mycovirus from Rhizoctonia fumigata AG-Ba isolate C-314 Baishi. Arch. Virol. 2015, 160, 2371–2374. [Google Scholar] [CrossRef]
- Sanderlin, R.S.; Ghabrial, S.A. Physicochemical properties of two distinct types of virus-like particles from Helminthosporium victoriae. Virology 1978, 87, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; De Micco, P.; de Lamballerie, X. Complete nucleotide sequence of Colorado tick fever virus segments M6, S1 and S2. J. Gen. Virol. 1997, 78, 2895–2899. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gai, X.; Chen, R.; Li, C.; Zhao, G.; Xia, Z. Characterization of three novel betapartitiviruses co-infecting the phytopathogenic fungus Rhizoctonia solani. Virus Res. 2019, 270, 197649. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.S.; Jeong, J.H.; Jeong, R.D.; Yoo, Y.B.; Yie, S.W.; Kim, K.H. Complete nucleotide sequence and genome organization of a dsRNA partitivirus infecting Pleurotus ostreatus. Virus Res. 2005, 108, 111–119. [Google Scholar] [CrossRef]
- Ghabrial, S.A. Origin, adaptation and evolutionary pathways of fungal viruses. Virus Genes 1998, 16, 119–131. [Google Scholar] [CrossRef]
- Wang, Q.; Mu, F.; Xie, J.; Cheng, J.S.; Fu, Y.; Jiang, D. A single ssRNA segment encoding RdRp is sufficient for replication, infection, and transmission of Ourmia-Like virus in fungi. Front. Microbiol. 2020, 11, 379. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, J.; Li, P.; Qiu, D.; Guo, L. Transcriptome-based discovery of Fusarium graminearum stress responses to FgHV1 Infection. Int. J. Mol. Sci. 2016, 17, 1922. [Google Scholar] [CrossRef]
- Romaine, C.P.; Ulhrich, P.; Schlagnhaufer, B. Transmission of La France isometric virus during basidiosporogenesis in Agaricus bisporus. Mycologia 1993, 85, 175–179. [Google Scholar] [CrossRef]
- Chu, Y.M.; Lim, W.S.; Yea, S.J.; Cho, J.D.; Lee, Y.W.; Kim, K.H. Complexity of dsRNA mycovirus isolated from Fusarium graminearum. Virus Genes 2004, 28, 135–143. [Google Scholar] [CrossRef]
- Won, H.K.; Park, S.J.; Kim, D.K.; Shin, M.J.; Kim, N.; Lee, S.H.; Kwon, Y.C.; Ko, H.K.; Ro, H.S.; Lee, H.S. Isolation and characterization of a mycovirus in Lentinula edodes. J. Microbiol. 2013, 51, 118–122. [Google Scholar] [CrossRef]
- Kwon, Y.C.; Jeong, D.W.; Gim, S.I.; Ro, H.S.; Lee, H.S. Curing viruses in Pleurotus ostreatus by growth on a limited nutrient medium containing cAMP and rifamycin. J. Virol. Methods 2012, 185, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Herrero, N.; Zabalgogeazcoa, I. Mycoviruses infecting the endophytic and entomopathogenic fungus Tolypocladium cylindrosporum. Virus Res. 2011, 160, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Romo, M.; Leuchtmann, A.; García, B.; Zabalgogeazcoa, I. A totivirus infecting the mutualistic fungal endophyte Epichloë festucae. Virus Res. 2007, 124, 38–43. [Google Scholar] [CrossRef]
- Van Diepeningen, A.D.; Debets, A.J.; Hoekstra, R.F. Dynamics of dsRNA mycoviruses in black Aspergillus populations. Fungal Genet. Biol. 2006, 43, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, M.F.; Jamal, A.; Petrou, M.A.; Cairns, T.C.; Bignell, E.M.; Coutts, R.H. The effects of dsRNA mycoviruses on growth and murine virulence of Aspergillus fumigatus. Fungal Genet. Biol. 2011, 48, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Nakamura, H.; Suzuki, N.; Kanematsu, S. Characterization of a new megabirnavirus that confers hypovirulence with the aid of a co-infecting partitivirus to the host fungus Rosellinia necatrix. Virus Res. 2016, 219, 73–82. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, J.; Wang, Y.; Hong, N.; Zhang, F.; Xu, W. Hypovirulence of the phytopathogenic fungus Botryosphaeria dothidea: Association with a coinfecting chrysovirus and a partitivirus. J. Virol. 2014, 88, 7517–7527. [Google Scholar] [CrossRef]
- Chun, J.; Ko, Y.H.; Kim, D.H. Transcriptome analysis of Cryphonectria parasitica infected with Cryphonectria hypovirus 1 (CHV1) reveals distinct genes related to fungal metabolites, Virulence, antiviral RNA-silencing, and their regulation. Front. Microbiol. 2020, 11, 1711. [Google Scholar] [CrossRef]
- Ding, F.; Cheng, J.; Fu, Y.; Chen, T.; Li, B.; Jiang, D.; Xie, J. Early transcriptional response to DNA virus infection in Sclerotinia sclerotiorum. Viruses 2019, 11, 278. [Google Scholar] [CrossRef]
- Qu, Z.; Fu, Y.; Lin, Y.; Zhao, Z.; Zhang, X.; Cheng, J.; Xie, J.; Chen, T.; Li, B.; Jiang, D. Transcriptional responses of Sclerotinia sclerotiorum to the Infection by SsHADV-1. J. Fungi 2021, 7, 493. [Google Scholar] [CrossRef]
- Lee, K.; Cho, W.; Yu, J.; Son, M.; Choi, H.; Min, K.; Lee, Y.-W.; Kim, K.-H. A comparison of transcriptional patterns and mycological phenotypes following infection of Fusarium graminearum by four mycoviruses. PLoS ONE 2014, 9, e100989. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Liu, X.; Benny, U.; Kistler, H.; VanEtten, H. Genes determining pathogenicity to pea are clustered on a supernumerary chromosome in the fungal plant pathogen Nectria haematococca. Plant J. 2001, 25, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Valette-Collet, O.; Cimerman, A.; Reignault, P.; Levis, C.; Boccara, M. Disruption of Botrytis cinerea pectin methylesterase gene Bcpme1 reduces virulence on several host plants. Mol. Plant Microbe Interact. 2003, 16, 360–367. [Google Scholar] [CrossRef]
- Kars, I.; Krooshof, G.H.; Wagemakers, L.; Joosten, R.; Benen, J.A.; van Kan, J.A. Necrotizing activity of five Botrytis cinerea endopolygalacturonases produced in Pichia pastoris. Plant J. 2005, 43, 213–225. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, A.; Zhao, L.; Sun, Y.; Chen, Y.; Li, C.; Dong, W.; Yang, G. Horizontal and Vertical Transmission of a Mycovirus Closely Related to the Partitivirus RhsV717 That Confers Hypovirulence in Rhizoctonia solani. Viruses 2023, 15, 2088. https://doi.org/10.3390/v15102088
Sun A, Zhao L, Sun Y, Chen Y, Li C, Dong W, Yang G. Horizontal and Vertical Transmission of a Mycovirus Closely Related to the Partitivirus RhsV717 That Confers Hypovirulence in Rhizoctonia solani. Viruses. 2023; 15(10):2088. https://doi.org/10.3390/v15102088
Chicago/Turabian StyleSun, Aili, Lianjing Zhao, Yang Sun, Yingrui Chen, Chengyun Li, Wenhan Dong, and Genhua Yang. 2023. "Horizontal and Vertical Transmission of a Mycovirus Closely Related to the Partitivirus RhsV717 That Confers Hypovirulence in Rhizoctonia solani" Viruses 15, no. 10: 2088. https://doi.org/10.3390/v15102088