
African Swine Fever Vaccine Candidate ASFV-G-ΔI177L Produced in the Swine Macrophage-Derived Cell Line IPKM Remains Genetically Stable and Protective against Homologous Virulent Challenge
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Sequencing and Analysis of the ASFV-G-∆I177L Genome
2.3. Differential Detection of ASFV-G Genome in Challenge Animals
2.4. Detection of ASFV Specific Antibodies
2.5. Evaluation of ASFV-G-ΔI177Lp10 Efficacy in Domestic Pigs
3. Results and Discussion
3.1. Successive Passages of ASFV-G-ΔI177L in IPKM Cells
3.2. Genomic Modifications of ASFV-G-ΔI177L during Passages in IPKM Cells
3.3. Assessment of Kinetic Replication of ASFV-G-ΔI177L in IPKM Cells
3.4. Assessment of ASFV-G-ΔI177Lp10 Replication in Domestic Pigs
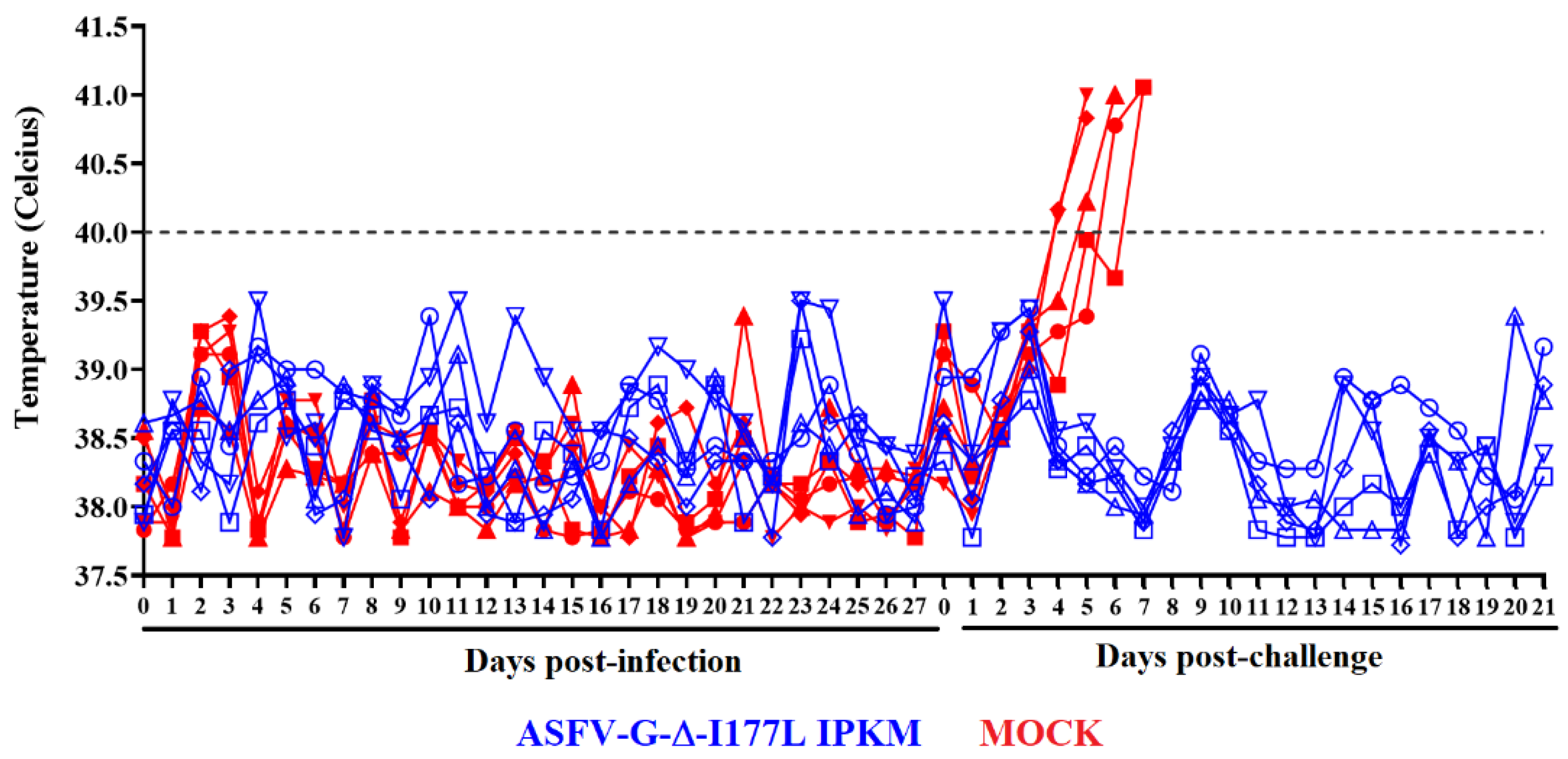
3.5. Assessment of the Efficacy of ASFV-G-ΔI177Lp10 to Protect Domestic Pigs against the Challenge with the Virulent Parental ASFV-G
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gonzales, W.; Moreno, C.; Duran, U.; Henao, N.; Bencosme, M.; Lora, P.; Reyes, R.; Nunez, R.; De Gracia, A.; Perez, A.M. African swine fever in the Dominican Republic. Transbound. Emerg. Dis. 2021, 68, 3018–3019. [Google Scholar] [CrossRef]
- Tulman, E.R.; Delhon, G.A.; Ku, B.K.; Rock, D.L. African Swine Fever Virus. In Lesser Known Large dsDNA Viruses; Etten, V., Ed.; Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2009; Volume 328, pp. 43–87. [Google Scholar]
- Spinard, E.; Azzinaro, P.; Rai, A.; Espinoza, N.; Ramirez-Medina, E.; Valladares, A.; Borca, M.V.; Gladue, D.P. Complete Structural Predictions of the Proteome of African Swine Fever Virus Strain Georgia 2007. Microbiol. Resour. Announc. 2022, 11, e00881-22. [Google Scholar] [CrossRef]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Holinka, L.G.; Velazquez-Salinas, L.; Zhu, J.; Gladue, D.P. Development of a Highly Effective African Swine Fever Virus Vaccine by Deletion of the I177L Gene Results in Sterile Immunity against the Current Epidemic Eurasia Strain. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Tran, X.H.; Le, T.T.P.; Nguyen, Q.H.; Do, T.T.; Nguyen, V.D.; Gay, C.G.; Borca, M.V.; Gladue, D.P. African swine fever virus vaccine candidate ASFV-G-DeltaI177L efficiently protects European and native pig breeds against circulating Vietnamese field strain. Transbound. Emerg. Dis. 2021, 69, e497–e504. [Google Scholar] [CrossRef]
- Lewis, T.; Zsak, L.; Burrage, T.G.; Lu, Z.; Kutish, G.F.; Neilan, J.G.; Rock, D.L. An African swine fever virus ERV1-ALR homologue, 9GL, affects virion maturation and viral growth in macrophages and viral virulence in swine. J. Virol. 2000, 74, 1275–1285. [Google Scholar]
- Zhang, Y.; Ke, J.; Zhang, J.; Yang, J.; Yue, H.; Zhou, X.; Qi, Y.; Zhu, R.; Miao, F.; Li, Q.; et al. African Swine Fever Virus Bearing an I226R Gene Deletion Elicits Robust Immunity in Pigs to African Swine Fever. J. Virol. 2021, 95, e0119921. [Google Scholar] [CrossRef]
- Gladue, D.P.; Ramirez-Medina, E.; Vuono, E.; Silva, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Borca, M.V. Deletion of the A137R Gene from the Pandemic Strain of African Swine Fever Virus Attenuates the Strain and Offers Protection against the Virulent Pandemic Virus. J. Virol. 2021, 95, e0113921. [Google Scholar] [CrossRef] [PubMed]
- Ran, Y.; Li, D.; Xiong, M.G.; Liu, H.N.; Feng, T.; Shi, Z.W.; Li, Y.H.; Wu, H.N.; Wang, S.Y.; Zheng, H.X.; et al. African swine fever virus I267L acts as an important virulence factor by inhibiting RNA polymerase III-RIG-I-mediated innate immunity. PLoS Pathog. 2022, 18, e1010270. [Google Scholar] [CrossRef]
- Li, D.; Zhang, J.; Yang, W.; Li, P.; Ru, Y.; Kang, W.; Li, L.; Ran, Y.; Zheng, H. African swine fever virus protein MGF-505-7R promotes virulence and pathogenesis by inhibiting JAK1- and JAK2-mediated signaling. J. Biol. Chem. 2021, 297, 101190. [Google Scholar] [CrossRef]
- Li, D.; Liu, Y.; Qi, X.; Wen, Y.; Li, P.; Ma, Z.; Liu, Y.; Zheng, H.; Liu, Z. African Swine Fever Virus MGF-110-9L-deficient Mutant Has Attenuated Virulence in Pigs. Virol. Sin. 2021, 36, 187–195. [Google Scholar] [CrossRef]
- O’Donnell, V.; Holinka, L.G.; Krug, P.W.; Gladue, D.P.; Carlson, J.; Sanford, B.; Alfano, M.; Kramer, E.; Lu, Z.; Arzt, J.; et al. African swine fever virus Georgia 2007 with a deletion of virulence-associated gene 9GL (B119L), when administered at low doses, leads to virus attenuation in swine and induces an effective protection against homologous challenge. J. Virol. 2015, 89, 8556–8566. [Google Scholar] [CrossRef]
- O’Donnell, V.; Holinka, L.G.; Gladue, D.P.; Sanford, B.; Krug, P.W.; Lu, X.; Arzt, J.; Reese, B.; Carrillo, C.; Risatti, G.R.; et al. African Swine Fever Virus Georgia Isolate Harboring Deletions of MGF360 and MGF505 Genes Is Attenuated in Swine and Confers Protection against Challenge with Virulent Parental Virus. J. Virol. 2015, 89, 6048–6056. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Rai, A.; Espinoza, N.; Velazquez-Salinas, L.; Gladue, D.P. ASF vaccine candidate ASFV-G-∆I177L does not exhibit residual virulence in long term clinical studies. Pathogens 2023, 12, 805. [Google Scholar] [CrossRef] [PubMed]
- Xuan, T.H.; Phuong, L.T.T.; Huy, N.Q.; Thuy, D.T.T.; Dung, N.V.; Quang, P.H.; Ngôn, Q.V.; Rai, A.; Gay, C.G.; Gladue, D.P.; et al. Evaluation of the safety profile of the ASFV vaccine candidate ASFV-G-∆I177L. Viruses 2022, 14, 896. [Google Scholar] [CrossRef]
- Krug, P.; Holinka, L.; O’Donnell, V.; Rees, B.; Sanford, B.; Fernandez Sainz, I.; Gladue, D.P.; Arzt, J.; Rodriguez, R.L.; Risatti, G.R.; et al. The progressive adaptation of a Georgian isolate of African swine fever virus to Vero cells leads to a gradual attenuation of virulence in swine corresponding with major modifications of the viral genome. J. Virol. 2015, 89, 2324–2332. [Google Scholar] [CrossRef] [PubMed]
- Takenouchi, T.; Masujin, K.; Suzuki, S.; Haraguchi, S.; Hiramatsu, K.; Kokuho, T.; Uenishi, H. Establishment and characterization of the immortalized porcine lung-derived mononuclear phagocyte cell line. Front. Vet. Sci. 2022, 9, 1058124. [Google Scholar] [CrossRef]
- World Organisation for Animal Health. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 12th ed; World Organisation for Animal Health: Paris, French, 2023; Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/A_summry.htm (accessed on 4 September 2023).
- Reed, L.J. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Velazquez-Salinas, L.; Ramirez-Medina, E.; Rai, A.; Pruitt, S.; Vuono, E.; Espinoza, N.; Gay, C.; Witte, S.; Gladue, D.P.; Borca, M.V. Confirming the absence of parental African swine fever virus as a potential contaminant of recombinant live attenuated ASF vaccines. Biologicals 2023, 83, 101685. [Google Scholar] [CrossRef]
- Carlson, J.; O’Donnell, V.; Alfano, M.; Velazquez Salinas, L.; Holinka, L.G.; Krug, P.W.; Gladue, D.P.; Higgs, S.; Borca, M.V. Association of the host immune response with protection using a live attenuated African swine fever virus model. Viruses 2016, 8, 291. [Google Scholar] [CrossRef]
- Masujin, K.; Kitamura, T.; Kameyama, K.; Okadera, K.; Nishi, T.; Takenouchi, T.; Kitani, H.; Kokuho, T. An immortalized porcine macrophage cell line competent for the isolation of African swine fever virus. Sci. Rep. 2021, 11, 4759. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, X.; Nie, Y.; Li, H.; Chen, W.; Lin, W.; Chen, F.; Xie, Q. African Swine Fever Virus Protein E199L Promotes Cell Autophagy through the Interaction of PYCR2. Virol. Sin. 2021, 36, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.M.; Hernáez, B.; Guerra, M.; Fraile-Ramos, A.; Andrés, G. African Swine Fever Virus Protein pE199L Mediates Virus Entry by Enabling Membrane Fusion and Core Penetration. J. Virol. 2020, 11, e00789-20. [Google Scholar] [CrossRef]
- Onisk, D.V.; Borca, M.V.; Kutish, S.; Kramer, E.; Irusta, P.; Rock, D.L. Passively Transferred African Swine Fever Virus Antibodies Protect Swine against Lethal Infection. Virology 1994, 198, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Oura, C.A.L.; Denyer, M.S.; Takamatsu, H.; Parkhouse, R.M.E. In vivo depletion of CD8+ T lymphocytes abrogates protective immunity to African swine fever virus Free. J. Gen. Virol. 2005, 86, 2445–2450. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borca, M.V.; Rai, A.; Espinoza, N.; Ramirez-Medina, E.; Spinard, E.; Velazquez-Salinas, L.; Valladares, A.; Silva, E.; Burton, L.; Meyers, A.; et al. African Swine Fever Vaccine Candidate ASFV-G-ΔI177L Produced in the Swine Macrophage-Derived Cell Line IPKM Remains Genetically Stable and Protective against Homologous Virulent Challenge. Viruses 2023, 15, 2064. https://doi.org/10.3390/v15102064
Borca MV, Rai A, Espinoza N, Ramirez-Medina E, Spinard E, Velazquez-Salinas L, Valladares A, Silva E, Burton L, Meyers A, et al. African Swine Fever Vaccine Candidate ASFV-G-ΔI177L Produced in the Swine Macrophage-Derived Cell Line IPKM Remains Genetically Stable and Protective against Homologous Virulent Challenge. Viruses. 2023; 15(10):2064. https://doi.org/10.3390/v15102064
Chicago/Turabian StyleBorca, Manuel V., Ayushi Rai, Nallely Espinoza, Elizabeth Ramirez-Medina, Edward Spinard, Lauro Velazquez-Salinas, Alyssa Valladares, Ediane Silva, Leeanna Burton, Amanda Meyers, and et al. 2023. "African Swine Fever Vaccine Candidate ASFV-G-ΔI177L Produced in the Swine Macrophage-Derived Cell Line IPKM Remains Genetically Stable and Protective against Homologous Virulent Challenge" Viruses 15, no. 10: 2064. https://doi.org/10.3390/v15102064