
Evidence of Circulation and Phylogenetic Analysis of Hepatitis E Virus (HEV) in Wild Boar in South-East Italy
 , , , , , ,
, , , , , ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Sample Preparation and Nucleic Acids Extraction
2.3. Real-Time Quantitative Reverse Transcription PCR (RTqPCR)
2.4. RT-PCR and Sequence Analysis
2.5. Full-Length Genome Sequencing of HEV
2.6. Phylogenetic Analysis
2.7. GenBank Sequence Submission
2.8. Statistical Analysis
3. Results
3.1. Molecular Detection and Quantification of HEV RNA
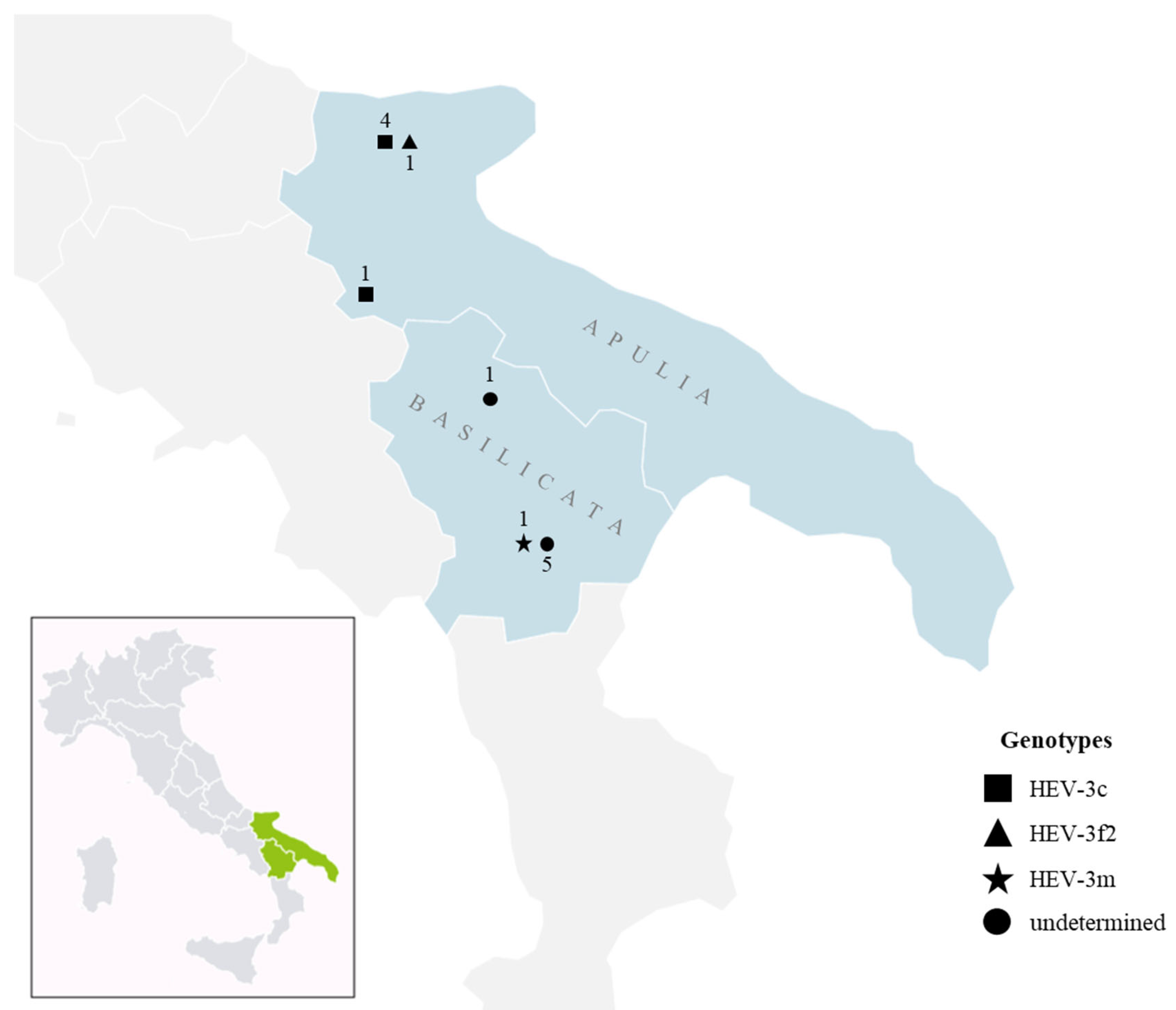
3.2. Genotyping and Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Association for the Study of the Liver (EASL). EASL Clinical Practice Guidelines on hepatitis E virus infection. J. Hepatol. 2018, 68, 1256–1271. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Hepatitis E. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-e (accessed on 1 February 2023).
- Kamar, N.; Izopet, J.; Pavio, N.; Aggarwal, R.; Labrique, A.; Wedemeyer, H.; Dalton, H.R. Hepatitis E virus infection. Nat. Rev. Dis. Primers 2017, 3, 17086. [Google Scholar] [CrossRef] [PubMed]
- Pavio, N.; Doceul, V.; Bagdassarian, E.; Johne, R. Recent knowledge on hepatitis E virus in Suidae reservoirs and transmission routes to human. Vet. Res. 2017, 48, 78. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernandez Escamez, P.S.; Herman, L.; Koutsoumanis, K.; Lindqvist, R.; Nørrung, B.; et al. Scientific Opinion on the public health risks associated with hepatitis E virus (HEV) as a food-borne pathogen. EFSA J. 2017, 15, e04886. [Google Scholar] [CrossRef] [PubMed]
- SEIEVA. Bullet No. 10—Update 2021. 2022. Available online: https://www.epicentro.iss.it/epatite/bollettino/Bollettino-n-10-marzo-2022.pdf (accessed on 1 February 2023).
- Purdy, M.A.; Drexler, J.F.; Meng, X.J.; Norder, H.; Okamoto, H.; Van der Poel, W.H.M.; Reuter, G.; de Souza, W.M.; Ulrich, R.G.; Smith, D.B. ICTV Virus Taxonomy Profile: Hepeviridae 2022. J. Gen. Virol. 2022, 103, 001778. [Google Scholar] [CrossRef] [PubMed]
- Johne, R.; Dremsek, P.; Reetz, J.; Heckel, G.; Hess, M.; Ulrich, R.G. Hepeviridae: An expanding family of vertebrate viruses. Infect. Genet. Evol. 2014, 27, 212–229. [Google Scholar] [CrossRef]
- La Rosa, G.; Muscillo, M.; Vennarucci, V.S.; Garbuglia, A.R.; La Scala, P.; Capobianchi, M.R. Hepatitis E virus in Italy: Molecular analysis of travel-related and autochthonous cases. J. Gen. Virol. 2011, 92, 1617–1626. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, J.R.; Oliveira, D.; Rivadulla, E.; Abreu-Silva, J.; Varela, M.F.; Romalde, J.L.; Nascimento, M.S. Hepatitis E virus genotype 3 in mussels (Mytilus galloprovinciallis), Spain. Food Microbiol. 2016, 58, 13–15. [Google Scholar] [CrossRef]
- Veličković, N.; Ferreira, E.; Djan, M.; Ernst, M.; Obreht Vidaković, D.; Monaco, A.; Fonseca, C. Demographic history, current expansion and future management challenges of wild boar populations in the Balkans and Europe. Heredity 2016, 117, 348–357. [Google Scholar] [CrossRef]
- Istituto Superiore per la Protezione e la Ricerca Ambientale (ISPRA). Available online: https://www.isprambiente.gov.it/files2023/area-stampa/comunicati-stampa/comunicatocinghiali-1.pdf (accessed on 1 February 2023).
- Mrzljak, A.; Balen, I.; Barbic, L.; Ilic, M.; Vilibic-Cavlek, T. Hepatitis E virus in professionally exposed: A reason for concern? World J. Hepatol. 2021, 13, 723–730. [Google Scholar] [CrossRef]
- Fanelli, A.; Tizzani, P.; Buonavoglia, D. A systematic review and meta-analysis of hepatitis E virus (HEV) in wild boars. Res. Vet. Sci. 2022, 142, 54–69. [Google Scholar] [CrossRef] [PubMed]
- Lucarelli, C.; Spada, E.; Taliani, G.; Chionne, P.; Madonna, E.; Marcantonio, C.; Pezzotti, P.; Bruni, R.; La Rosa, G.; Pisani, G.; et al. High prevalence of anti-hepatitis E virus antibodies among blood donors in central Italy, February to March 2014. Eurosurveillance 2016, 21, 30299. [Google Scholar] [CrossRef]
- La Bella, G.; Basanisi, M.G.; Nobili, G.; Terio, V.; Suffredini, E.; La Salandra, G. First Report of Hepatitis E Virus in Shellfish in Southeast Italy. Appl. Sci. 2021, 11, 43. [Google Scholar] [CrossRef]
- Di Pasquale, S.; De Santis, P.; La Rosa, G.; Di Domenico, K.; Iaconelli, M.; Micarelli, G.; Martini, E.; Bilei, S.; De Medici, D.; Suffredini, E. Quantification and genetic diversity of Hepatitis E virus in wild boar (Sus scrofa) hunted for domestic consumption in Central Italy. Food Microbiol. 2019, 82, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Iso/ts 15216-2; Microbiology of Food Chain: Horizontal Method for Determination of Hepatitis a Virus and Norovirus Using Real-Time rt-pcr. Part 2: Method for Detection. International Organization for Standardization: Geneva, Switzerland, 2019.
- Szabo, K.; Trojnar, E.; Anheyer-Behmenburg, H.; Binder, A.; Schotte, U.; Ellerbroek, L.; Klein, G.; Johne, R. Detection of hepatitis E virus RNA in raw sausages and liver sausages from retail in Germany using an optimized method. Int. J. Food Microbiol. 2015, 215, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of Hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef]
- Garson, J.A.; Ferns, R.B.; Grant, P.R.; Ijaz, S.; Nastouli, E.; Szypulska, R.; Tedder, R.S. Minor groove binder modification of widely used TaqMan probe for hepatitis E virus reduces risk of false negative real-time PCR results. J. Virol. Methods 2012, 186, 157–160. [Google Scholar] [CrossRef]
- Iso/ts 15216-1; Microbiology of the Food Chain: Horizontal Method for Determination of Hepatitis a Virus and Norovirus Using Real-Time rt-pcr. Part 1: Method for Quantification. International Organization for Standardization: Geneva, Switzerland, 2017.
- Fogeda, M.; Avellón, A.; Cilla, C.G.; Echevarría, J.M. Imported and autochthonous hepatitis E virus strains in Spain. J. Med. Virol. 2009, 81, 1743–1749. [Google Scholar] [CrossRef]
- Mulder, A.C.; Kroneman, A.; Franz, E.; Vennema, H.; Tulen, A.D.; Takkinen, J.; Hofhuis, A.; Adlhoch, C. HEVnet: A One Health, collaborative, interdisciplinary network and sequence data repository for enhanced hepatitis E virus molecular typing, characterisation and epidemiological investigations. Eurosurveillance 2019, 24, 1800407. [Google Scholar] [CrossRef]
- Drexler, J.F.; Seelen, A.; Corman, V.M.; Fumie Tateno, A.; Cottontail, V.; Melim Zerbinati, R.; Gloza-Rausch, F.; Klose, S.M.; Adu-Sarkodie, Y.; Oppong, S.K.; et al. Bats worldwide carry hepatitis E virus-related viruses that form a putative novel genus within the family Hepeviridae. J. Virol. 2012, 86, 9134–9147. [Google Scholar] [CrossRef]
- Scotto-Lavino, E.; Du, G.; Frohman, M.A. 3’ End cDNA amplification using classic RACE. Nat. Protoc. 2006, 1, 2742–2745. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Iaconelli, M.; Bonanno Ferraro, G.; Mancini, P.; Suffredini, E.; Veneri, C.; Ciccaglione, A.R.; Bruni, R.; Della Libera, S.; Bignami, F.; Brambilla, M.; et al. Nine-Year Nationwide Environmental Surveillance of Hepatitis E Virus in Urban Wastewaters in Italy (2011–2019). Int. J. Environ. Res. Public Health 2020, 17, 2059. [Google Scholar] [CrossRef]
- Martelli, F.; Caprioli, A.; Zengarini, M.; Marata, A.; Fiegna, C.; Di Bartolo, I.; Ruggeri, F.M.; Delogu, M.; Ostanello, F. Detection of hepatitis E virus (HEV) in a demographic managed wild boar (Sus scrofa scrofa) population in Italy. Vet. Microbiol. 2008, 126, 74–81. [Google Scholar] [CrossRef]
- Caruso, C.; Modesto, P.; Bertolini, S.; Peletto, S.; Acutis, P.L.; Dondo, A.; Robetto, S.; Mignone, W.; Orusa, R.; Ru, G.; et al. Serological and virological survey of hepatitis E virus in wild boar populations in northwestern Italy: Detection of HEV subtypes 3e and 3f. Arch. Virol. 2015, 160, 153–160. [Google Scholar] [CrossRef]
- Mazzei, M.; Nardini, R.; Verin, R.; Forzan, M.; Poli, A.; Tolari, F. Serologic and molecular survey for hepatitis E virus in wild boar (Sus scrofa) in Central Italy. New Microbes New Infect. 2015, 7, 41–47. [Google Scholar] [CrossRef]
- Montagnaro, S.; De Martinis, C.; Sasso, S.; Ciarcia, R.; Damiano, S.; Auletta, L.; Iovane, V.; Zottola, T.; Pagnini, U. Viral and antibody prevalence of hepatitis E in European wild boars (Sus scrofa) and hunters at zoonotic risk in the Latium region. J. Comp. Pathol. 2015, 153, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Serracca, L.; Battistini, R.; Rossini, I.; Mignone, W.; Peletto, S.; Boin, C.; Pistone, G.; Ercolini, R.; Ercolini, C. Molecular investigation on the presence of hepatitis E virus (HEV) in wild game in North-western Italy. Food Environ. Virol. 2015, 7, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Di Profio, F.; Melegari, I.; Sarchese, V.; Robetto, S.; Marruchella, G.; Bona, M.C.; Orusa, R.; Martella, V.; Marsilio, F.; Di Martino, B. Detection and genetic characterization of hepatitis E virus (HEV) genotype 3 subtype c in wild boars in Italy. Arch. Virol. 2016, 161, 2829–2834. [Google Scholar] [CrossRef]
- Aprea, G.; Amoroso, M.G.; Di Bartolo, I.; D’Alessio, N.; Di Sabatino, D.; Boni, A.; Cioffi, B.; D’Angelantonio, D.; Scattolini, S.; De Sabato, L.; et al. Molecular detection and phylogenetic analysis of hepatitis E virus strains circulating in wild boars in south-central Italy. Transbound. Emerg. Dis. 2018, 65, e25–e31. [Google Scholar] [CrossRef] [PubMed]
- De Sabato, L.; Ostanello, F.; De Grossi, L.; Marcario, A.; Franzetti, B.; Monini, M.; Di Bartolo, I. Molecular survey of HEV infection in wild boar population in Italy. Transbound. Emerg. Dis. 2018, 65, 1749–1756. [Google Scholar] [CrossRef] [PubMed]
- Zecchin, B.; Schivo, A.; Milani, A.; Fusaro, A.; Zamperin, G.; Bellinati, L.; Ceglie, L.; Natale, A.; Bonfanti, L.; Cunial, G.; et al. Identification of a zoonotic genotype 3 hepatitis E subtype in wildlife in north-eastern Italy. Infect. Genet. Evol. 2019, 71, 16–20. [Google Scholar] [CrossRef]
- De Sabato, L.; Amoroso, M.G.; Ianiro, G.; Esposito, C.; De Grossi, L.; Fusco, G.; Barone, A.; Martini, E.; Ostanello, F.; Di Bartolo, I. Detection of Hepatitis E Virus in Livers and Muscle Tissues of Wild Boars in Italy. Food Environ. Virol. 2020, 12, 1–8. [Google Scholar] [CrossRef]
- Bonardi, S.; Filipello, V.; Pavoni, E.; Carta, V.; Bolzoni, L.; Corradi, M.; Gilioli, S.; Losio, M.N. Geographical restriction of Hepatitis E virus circulation in wild boars (Sus scrofa) in Emilia-Romagna region, Northern Italy. Ital. J. Food Saf. 2020, 9, 8463. [Google Scholar] [CrossRef]
- Arnaboldi, S.; Righi, F.; Carta, V.; Bonardi, S.; Pavoni, E.; Bianchi, A.; Losio, M.N.; Filipello, V. Hepatitis E Virus (HEV) Spread and Genetic Diversity in Game Animals in Northern Italy. Food Environ. Virol. 2021, 13, 146–153. [Google Scholar] [CrossRef]
- Forzan, M.; Pacini, M.I.; Periccioli, M.; Mazzei, M. Hepatitis E Virus RNA Presence in Wild Boar Carcasses at Slaughterhouses in Italy. Animals 2021, 11, 1624. [Google Scholar] [CrossRef]
- Pierini, I.; Di Bartolo, I.; Manuali, E.; Pirani, S.; Bazzucchi, M.; Moscati, L.; De Mia, G.M.; Giammarioli, M. Hepatitis E virus (HEV) genotype 3 diversity: Identification of a novel HEV subtype in wild boar in Central Italy. Transbound. Emerg. Dis. 2021, 68, 2121–2129. [Google Scholar] [CrossRef]
- Lorusso, P.; Bonerba, E.; Pandiscia, A.; Mottola, A.; Di Pinto, A.; Piredda, R.; Terio, V. Occurrence of hepatitis E virus (HEV) in Calabrian wild boars. Int. J. Food Microbiol. 2022, 371, 109671. [Google Scholar] [CrossRef] [PubMed]
- Ferri, G.; Piccinini, A.; Olivastri, A.; Vergara, A. Hepatitis E virus detection in hunted wild boar (Sus scrofa) livers in Central Italy. Ital. J. Food Saf. 2022, 21, 9979. [Google Scholar] [CrossRef]
- Battistini, R.; Serracca, L.; Razzuoli, E.; Listorti, V.; Guardone, L.; Dellepiane, M.; Pussini, N.; Brunelli, F.; Peletto, S.; Beltramo, C.; et al. Hepatitis E Virus (HEV): Identification of Subtypes 3b and 3m in Wild Boar Population in Liguria Region, Italy. Pathogens 2022, 11, 1236. [Google Scholar] [CrossRef] [PubMed]
- De Massis, F.; Aprea, G.; Scattolini, S.; D’Angelantonio, D.; Chiaverini, A.; Mangone, I.; Perilli, M.; Colacicco, G.; Olivieri, S.; Pomilio, F.; et al. Detection of Hepatitis E Virus (HEV) in Pigs and in the Wild Boar (Sus scrofa) Population of Chieti Province, Abruzzo Region, Italy. Appl. Microbiol. 2022, 2, 818–826. [Google Scholar] [CrossRef]
- Lhomme, S.; Top, S.; Bertagnoli, S.; Dubois, M.; Guerin, J.L.; Izopet, J. Wildlife reservoir for hepatitis E virus, southwestern France. Emerg. Infect. Dis. 2015, 21, 1224–1226. [Google Scholar] [CrossRef] [PubMed]
- Kubankova, M.; Kralik, P.; Lamka, J.; Zakovcik, V.; Dolanský, M.; Vasickova, P. Prevalence of hepatitis E virus in populations of wild animals in comparison with animals bred in game enclosures. Food Environ. Virol. 2015, 7, 159–163. [Google Scholar] [CrossRef]
- Anheyer-Behmenburg, H.E.; Szabo, K.; Schotte, U.; Binder, A.; Klein, G.; Johne, R. Hepatitis E virus in wild boars and spillover infection in red and roe deer, Germany, 2013-2015. Emerg. Infect. Dis. 2017, 23, 130–133. [Google Scholar] [CrossRef]
- Adlhoch, C.; Avellon, A.; Baylis, S.A.; Ciccaglione, A.R.; Couturier, E.; de Sousa, R.; Epštein, J.; Ethelberg, S.; Faber, M.; Fehér, Á.; et al. Hepatitis E virus: Assessment of the epidemiological situation in humans in Europe, 2014/15. J. Clin. Virol. 2016, 82, 9–16. [Google Scholar] [CrossRef]
- Bouwknegt, M.; Frankena, K.; Rutjes, S.A.; Wellenberg, G.J.; de Roda Husman, A.M.; van der Poel, W.H.; de Jong, M.C. Estimation of hepatitis E virus transmission among pigs due to contact-exposure. Vet. Res. 2008, 39, 40. [Google Scholar] [CrossRef]
- Risalde, M.A.; Rivero-Juárez, A.; Romero-Palomo, F.; Frías, M.; López-López, P.; Cano-Terriza, D.; García-Bocanegra, I.; Jiménez-Ruíz, S.; Camacho, Á.; Machuca, I.; et al. Persistence of hepatitis E virus in the liver of nonviremic naturally infected wild boar. PLoS ONE 2017, 12, e0186858. [Google Scholar] [CrossRef]
- Leblanc, D.; Poitras, E.; Gagné, M.J.; Ward, P.; Houde, A. Hepatitis E virus load in swine organs and tissues at slaughterhouse determined by real-time RT-PCR. Int. J. Food Microbiol. 2010, 139, 206–209. [Google Scholar] [CrossRef]
- Feurer, C.; Le Roux, A.; Rossel, R.; Barnaud, E.; Dumarest, M.; Garry, P.; Pavio, N. High load of hepatitis E viral RNA in pork livers but absence in pork muscle at French slaughterhouses. Int. J. Food Microbiol. 2018, 264, 25–30. [Google Scholar] [CrossRef]
- Salines, M.; Demange, A.; Stéphant, G.; Renson, P.; Bourry, O.; Andraud, M.; Rose, N.; Pavio, N. Persistent viremia and presence of hepatitis E virus RNA in pig muscle meat after experimental coinfection with porcine reproductive and respiratory syndrome virus. Int. J. Food Microbiol. 2019, 292, 144–149. [Google Scholar] [CrossRef]
- Lo Presti, A.; Bruni, R.; Villano, U.; Marcantonio, C.; Equestre, M.; Ciuffetelli, M.; Grimaldi, A.; Suffredini, E.; Di Pasquale, S.; De Medici, D.; et al. Phylogenetic analysis and epidemiological history of Hepatitis E virus 3f and 3c in swine and wild boar, Italy. Heliyon 2020, 6, e05110. [Google Scholar] [CrossRef]
- De Sabato, L.; Di Bartolo, I.; Lapa, D.; Capobianchi, M.R.; Garbuglia, A.R. Molecular Characterization of HEV Genotype 3 in Italy at Human/Animal Interface. Front. Microbiol. 2020, 11, 137. [Google Scholar] [CrossRef] [PubMed]
- Suin, V.; Klamer, S.E.; Hutse, V.; Wautier, M.; Jacques, M.; Abady, M.; Lamoral, S.; Verburgh, V.; Thomas, I.; Brochier, B.; et al. Epidemiology and genotype 3 subtype dynamics of hepatitis E virus in Belgium, 2010 to 2017. Eurosurveillance 2019, 24, 33–41. [Google Scholar] [CrossRef]
- Nicot, F.; Jeanne, N.; Roulet, A.; Lefebvre, C.; Carcenac, R.; Manno, M.; Dubois, M.; Kamar, N.; Lhomme, S.; Abravanel, F.; et al. Diversity of hepatitis E virus genotype 3. Rev. Med. Virol. 2018, 28, e1987. [Google Scholar] [CrossRef]
- Oeser, C.; Vaughan, A.; Said, B.; Ijaz, S.; Tedder, R.; Haywood, B.; Warburton, F.; Charlett, A.; Elson, R.; Morgan, D. Epidemiology of hepatitis E in England and Wales: A 10-uear retrospective surveillance study, 2008–2017. J. Infect. Dis. 2019, 202, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Lhomme, S.; Gallian, P.; Dimeglio, C.; Assal, A.; Abravanel, F.; Tiberghien, P.; Izopet, J. Viral load and clinical manifestations of hepatitis E virus genotype 3 infections. J. Viral Hepat. 2019, 26, 1139–1142. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Chimeno, M.; Forero, J.E.; Echevarría, J.M.; Muñoz-Bellido, J.L.; Vázquez-López, L.; Morago, L.; García-Galera, M.C.; Avellón, A. Full coding hepatitis E virus genotype 3 genome amplification method. J. Virol. Methods 2016, 230, 18–23. [Google Scholar] [CrossRef]
- Caballero-Gómez, J.; Jiménez-Ruiz, S.; Lopez-Lopez, P.; Vicente, J.; Risalde, M.A.; Cano-Terriza, D.; Frias, M.; Barasona, J.A.; Rivero, A.; García-Bocanegra, I.; et al. Emergent subtype of hepatitis E virus genotype 3 in wild boar in Spain. Transbound. Emerg. Dis. 2019, 66, 1803–1808. [Google Scholar] [CrossRef]
- Muñoz-Chimeno, M.; Bartúren, S.; García-Lugo, M.A.; Morago, L.; Rodríguez, Á.; Galán, J.C.; Pérez-Rivilla, A.; Rodríguez, M.; Millán, R.; Del Álamo, M.; et al. Hepatitis E virus genotype 3 microbiological surveillance by the Spanish Reference Laboratory: Geographic distribution and phylogenetic analysis of subtypes from 2009 to 2019. Eurosurveillance 2022, 27, 2100542. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, S.; Said, B.; Boxall, E.; Smit, E.; Morgan, D.; Tedder, R.S. Indigenous hepatitis E in England and wales from 2003 to 2012: Evidence of an emerging novel phylotype of viruses. J. Infect. Dis. 2014, 209, 1212–1218. [Google Scholar] [CrossRef]
- Wang, H.; Castillo-Contreras, R.; Saguti, F.; López-Olvera, J.R.; Karlsson, M.; Mentaberre, G.; Lindh, M.; Serra-Cobo, J.; Norder, H. Genetically similar hepatitis E virus strains infect both humans and wild boars in the Barcelona area, Spain, and Sweden. Transbound. Emerg. Dis. 2019, 66, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Nicot, F.; Dimeglio, C.; Migueres, M.; Jeanne, N.; Latour, J.; Abravanel, F.; Ranger, N.; Harter, A.; Dubois, M.; Lameiras, S.; et al. Classification of the Zoonotic Hepatitis E Virus Genotype 3 Into Distinct Subgenotypes. Front. Microbiol. 2021, 11, 634430. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Ijaz, S.; Tedder, R.S.; Hogema, B.; Zaaijer, H.L.; Izopet, J.; Bradley-Stewart, A.; Gunson, R.; Harvala, H.; Kokki, I.; et al. Variability and pathogenicity of hepatitis E virus genotype 3 variants. J. Gen. Virol. 2015, 96, 3255–3264. [Google Scholar] [CrossRef]
- Subissi, L.; Peeters, M.; Lamoral, S.; Klamer, S.; Suin, V.; Van Gucht, S. Subtype-specific differences in the risk of hospitalisation among patients infected with hepatitis E virus genotype 3 in Belgium, 2010-2018. Epidemiol. Infect. 2019, 147, e224. [Google Scholar] [CrossRef] [PubMed]
- Abravanel, F.; Dimeglio, C.; Castanier, M.; Péron, J.M.; Kamar, N.; Lhomme, S.; Izopet, J. Does HEV-3 subtype play a role in the severity of acute hepatitis E? Liver Int. 2020, 40, 333–337. [Google Scholar] [CrossRef]
- Chelli, E.; Suffredini, E.; De Santis, P.; De Medici, D.; Di Bella, S.; D’Amato, S.; Gucciardi, F.; Guercio, A.; Ostanello, F.; Perrone, V.; et al. Hepatitis E Virus Occurrence in Pigs Slaughtered in Italy. Animals 2021, 11, 277. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, P.; Bonerba, E.; Pandiscia, A.; Bozzo, G.; Piredda, R.; Manfredi, A.; Terio, V. Epidemiological and genetic evaluation of HEV in swine slaughtered in Sicily region (Italy). Int. J. Food Microbiol. 2023, 388, 110068. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| N. of Tested Samples | N. of Positive Samples (%) | Viral Load | Geometric Mean of Quantifiable Samples (g.c./g) | ||
|---|---|---|---|---|---|
| <LOQ | ≥LOQ | ||||
| Region | |||||
| Apulia | 50 | 6 (12.0%) | - | 6 | 1.50 × 105 |
| Basilicata | 75 | 7 (9.3%) | 4 | 3 | 2.81 × 104 |
| Sample Type | |||||
| Muscle | 47 | 6 (12.8%) | - | 6 | 1.5 × 105 |
| Liver | 78 | 7 (9.0%) | 4 | 3 | 2.81 × 104 |
| Gender | |||||
| Male | 48 | 3 (6.2%) | 2 | 1 | 4.82 × 104 |
| Female | 77 | 10 (12.9%) | 2 | 8 | 9.21 × 104 |
| Age (months) | |||||
| <12 | 10 | - | - | - | - |
| 13–24 | 43 | 6 (13.3%) | 1 | 5 | 2.06 × 105 |
| >24 | 72 | 7 (9.7%) | 3 | 4 | 2.86 × 104 |
| Total | 125 | 13 (10.4%) | 4 | 9 | 6.49 × 104 |
| Sample | Hevnet Assignment | Identity in the NCBI Database | ||||||
|---|---|---|---|---|---|---|---|---|
| ORF1 | ORF2 | |||||||
| Genotype | Subtype | Accession nr | Reference Strain | nt Identity % | Accession nr | Reference Strain | nt Identity % | |
| ITA/2021/wild boar/C1 | 3 | f | MZ289122 | isolate 179535MM66 | 94.2 | MZ289122 | isolate 179535MM66 | 95.1 |
| ITA/2021/wild boar/287 | 3 | c | FJ705359 | isolate wbGER27 | 98.8 | MT497761 | isolate S03_Germany | 95.9 |
| ITA/2021/wild boar/37155-1 | 3 | c | MH450021 | isolate HEVCU634 | 96.5 | MH450021 | isolate HEVCU634 | 94.9 |
| ITA/2021/wild boar/37155-2 | 3 | c | MF444063 | isolate HEPAC-11 | 97.1 | MH450021 | isolate HEVCU634 | 95.7 |
| ITA/2021/wild boar/37155-3 | 3 | c | MH504129 | isolate HEV 35_Poole_UK_050115 | 98.8 | MH450021 | isolate HEVCU634 | 96.2 |
| ITA/2021/wild boar/37155-4 | 3 | c | MH504129 | isolate HEV 35_Poole_UK_050115 | 96.5 | MH450021 | isolate HEVCU634 | 95.7 |
| ITA/2021/wild boar/337 * | 3 | m | MW355399 | isolate HESQL050 | 98.3 | MW355399 | isolate HESQL050 | 90.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
La Bella, G.; Basanisi, M.G.; Nobili, G.; Coppola, R.; Damato, A.M.; Donatiello, A.; Occhiochiuso, G.; Romano, A.C.; Toce, M.; Palazzo, L.; et al. Evidence of Circulation and Phylogenetic Analysis of Hepatitis E Virus (HEV) in Wild Boar in South-East Italy. Viruses 2023, 15, 2021. https://doi.org/10.3390/v15102021
La Bella G, Basanisi MG, Nobili G, Coppola R, Damato AM, Donatiello A, Occhiochiuso G, Romano AC, Toce M, Palazzo L, et al. Evidence of Circulation and Phylogenetic Analysis of Hepatitis E Virus (HEV) in Wild Boar in South-East Italy. Viruses. 2023; 15(10):2021. https://doi.org/10.3390/v15102021
Chicago/Turabian StyleLa Bella, Gianfranco, Maria Grazia Basanisi, Gaia Nobili, Rosa Coppola, Annita Maria Damato, Adelia Donatiello, Gilda Occhiochiuso, Antonella Cristina Romano, Mariateresa Toce, Lucia Palazzo, and et al. 2023. "Evidence of Circulation and Phylogenetic Analysis of Hepatitis E Virus (HEV) in Wild Boar in South-East Italy" Viruses 15, no. 10: 2021. https://doi.org/10.3390/v15102021