
Differential Replication and Cytokine Response between Vaccine and Very Virulent Marek’s Disease Viruses in Spleens and Bursas during Latency and Reactivation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Viruses, Experimental Animals, and Design
2.3. RNA and DNA Isolation from Bursa and Spleen
2.4. Absolute Quantitation of MDV Copies
2.5. Relative Quantification of Cytokine Expression Levels
2.6. Statistical Analysis
3. Results
3.1. Differences in MDV Load between Vaccine and Virulent Strain in Spleen and Bursa
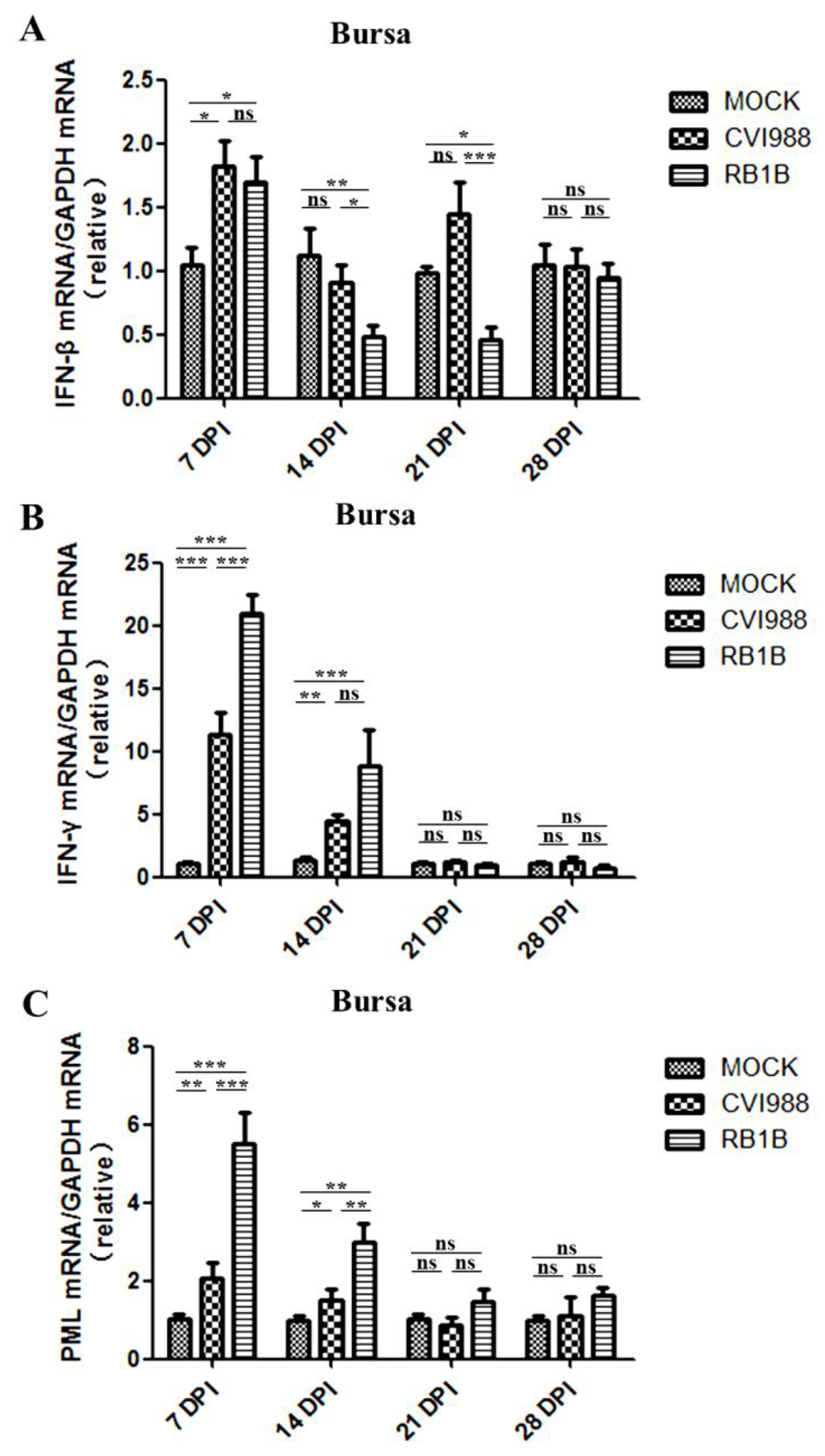
3.2. Effects of MDV Replication on Interferon and PML Expression in the Spleen and Bursa
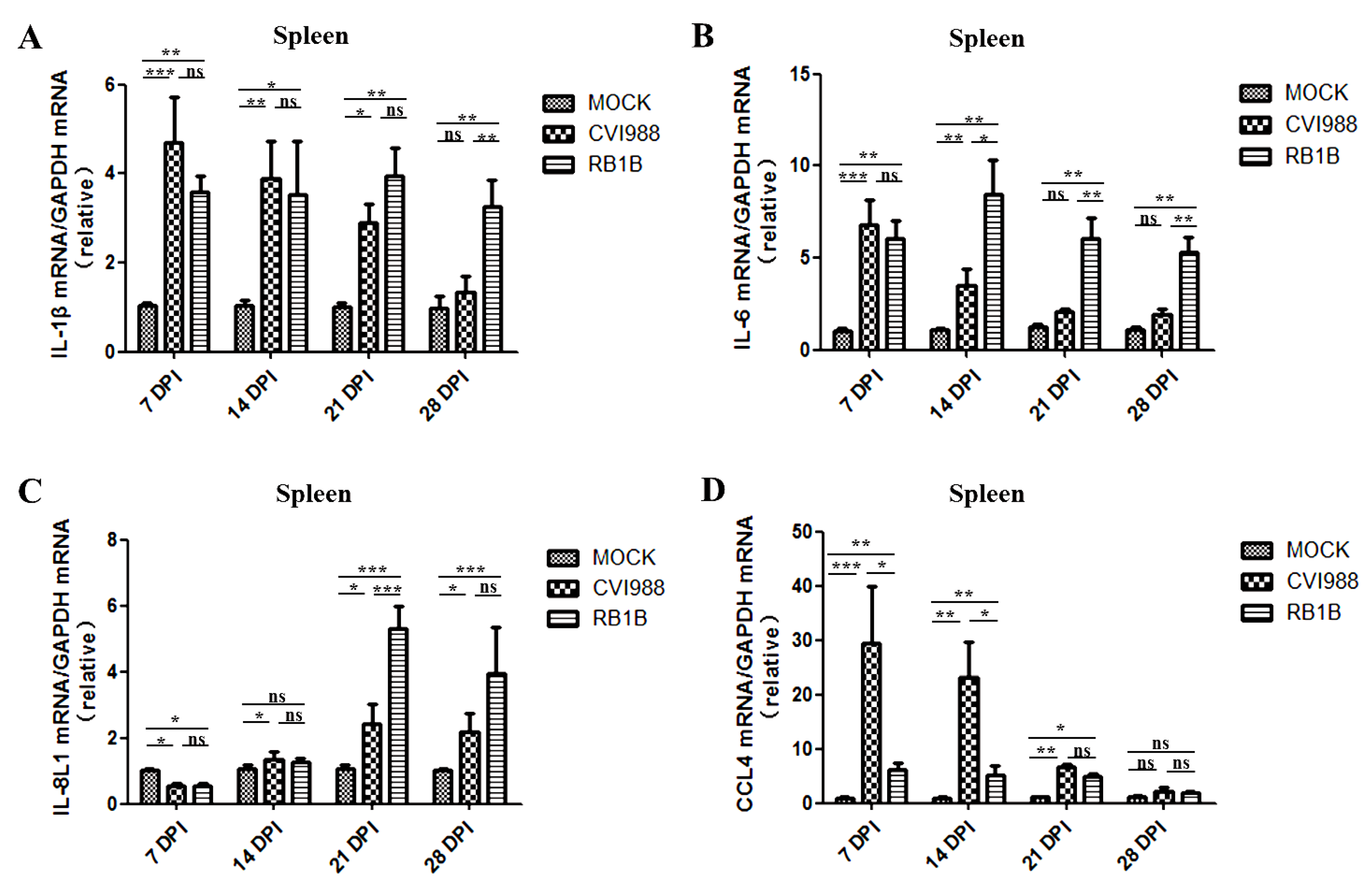
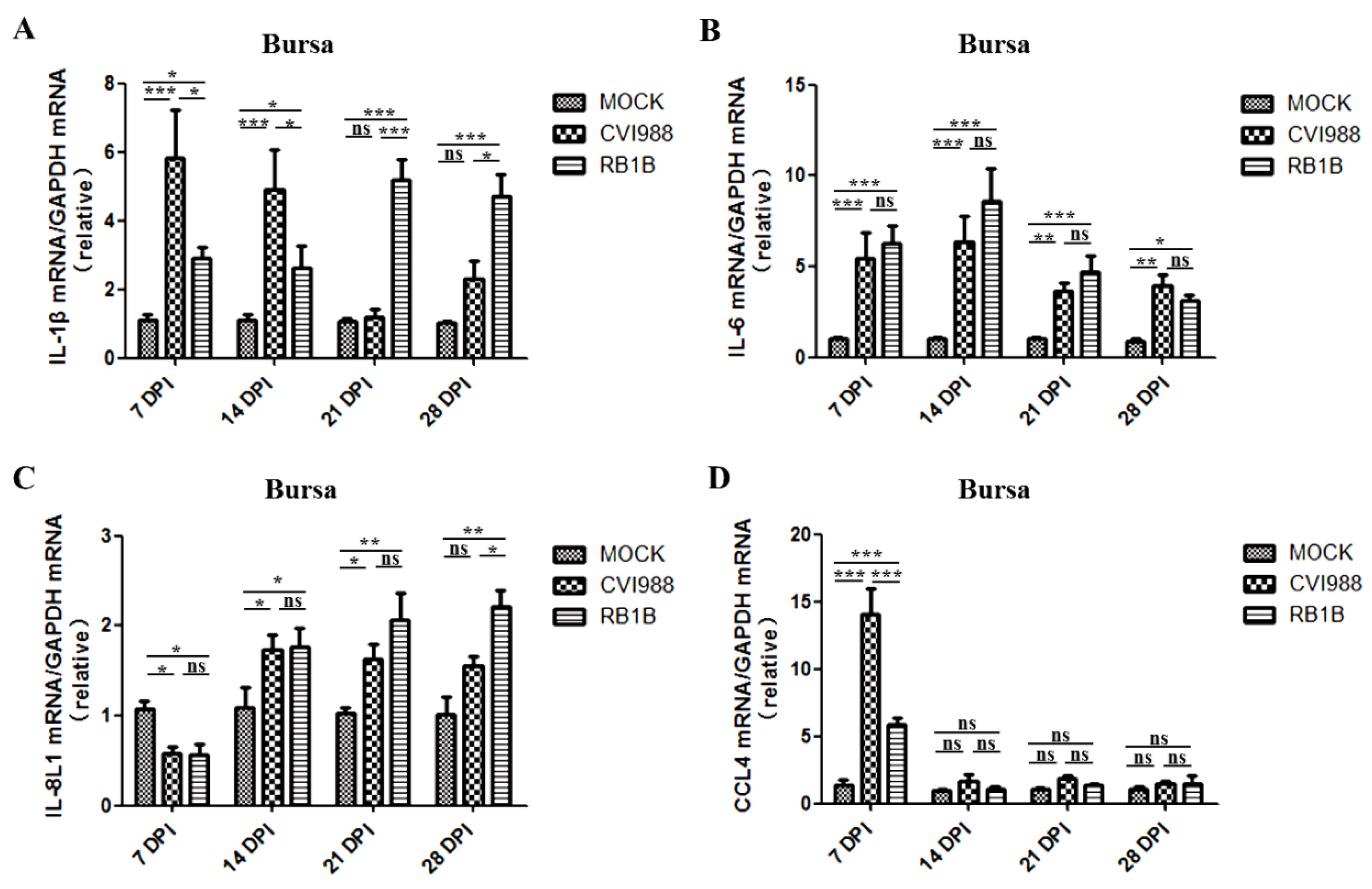
3.3. Effects of MDV Replication on and Chemokine Expression in the Spleen and Bursa
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nair, V. Spotlight on avian pathology: Marek’s disease. Avian Pathol. J. W.V.P.A 2018, 47, 440–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, B.; Zeb, J.; Hussain, S.; Aziz, M.U.; Circella, E.; Casalino, G.; Camarda, A.; Yang, G.; Buchon, N.; Sparagano, O. A Review on the Marek’s Disease Outbreak and Its Virulence-Related meq Genovariation in Asia between 2011 and 2021. Animals 2022, 12, 540. [Google Scholar] [CrossRef]
- Osterrieder, N.; Kamil, J.P.; Schumacher, D.; Tischer, B.K.; Trapp, S. Marek’s disease virus: From miasma to model. Nat. Rev. Microbiol. 2006, 4, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Careem, M.F.; Javaheri-Vayeghan, A.; Shanmuganathan, S.; Haghighi, H.R.; Read, L.R.; Haq, K.; Hunter, D.B.; Schat, K.A.; Heidari, M.; Sharif, S. Establishment of an aerosol-based Marek’s disease virus infection model. Avian Dis. 2009, 53, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Boodhoo, N.; Gurung, A.; Sharif, S.; Behboudi, S. Marek’s disease in chickens: A review with focus on immunology. Vet. Res. 2016, 47, 119. [Google Scholar] [CrossRef] [Green Version]
- Robinson, C.M.; Cheng, H.H.; Delany, M.E. Temporal kinetics of Marek’s disease herpesvirus: Integration occurs early after infection in both B and T cells. Cytogenet. Genome Res. 2014, 144, 142–154. [Google Scholar] [CrossRef]
- Teng, M.; Zheng, L.P.; Li, H.Z.; Ma, S.M.; Zhu, Z.J.; Chai, S.J.; Yao, Y.; Nair, V.; Zhang, G.P.; Luo, J. Pathogenicity and Pathotype Analysis of Henan Isolates of Marek’s Disease Virus Reveal Long-Term Circulation of Highly Virulent MDV Variant in China. Viruses 2022, 14, 1651. [Google Scholar] [CrossRef]
- Biggs, P.M.; Nair, V. The long view: 40 years of Marek’s disease research and Avian Pathology. Avian Pathol. J. W.V.P.A 2012, 41, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Baigent, S.J.; Nair, V.K.; Le Galludec, H. Real-time PCR for differential quantification of CVI988 vaccine virus and virulent strains of Marek’s disease virus. J. Virol. Methods 2016, 233, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Petherbridge, L.; Howes, K.; Baigent, S.J.; Sacco, M.A.; Evans, S.; Osterrieder, N.; Nair, V. Replication-competent bacterial artificial chromosomes of Marek’s disease virus: Novel tools for generation of molecularly defined herpesvirus vaccines. J. Virol. 2003, 77, 8712–8718. [Google Scholar] [CrossRef]
- Boodhoo, N.; Behboudi, S. Differential Virus-Specific IFN-Gamma Producing T Cell Responses to Marek’s Disease Virus in Chickens With B19 and B21 MHC Haplotypes. Front. Immunol. 2021, 12, 784359. [Google Scholar] [CrossRef]
- Bacon, L.D.; Witter, R.L. Influence of B-haplotype on the relative efficacy of Marek’s disease vaccines of different serotypes. Avian Dis. 1993, 37, 53–59. [Google Scholar] [CrossRef]
- Conradie, A.M.; Bertzbach, L.D.; Trimpert, J.; Patria, J.N.; Murata, S.; Parcells, M.S.; Kaufer, B.B. Distinct polymorphisms in a single herpesvirus gene are capable of enhancing virulence and mediating vaccinal resistance. PLoS Pathog. 2020, 16, e1009104. [Google Scholar] [CrossRef]
- Read, A.F.; Baigent, S.J.; Powers, C.; Kgosana, L.B.; Blackwell, L.; Smith, L.P.; Kennedy, D.A.; Walkden-Brown, S.W.; Nair, V.K. Imperfect Vaccination Can Enhance the Transmission of Highly Virulent Pathogens. PLoS Biol. 2015, 13, e1002198. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Kong, Z.; Mehboob, A.; Jiang, B.; Xu, J.; Cai, Y.; Liu, W.; Hong, J.; Li, Y. Transcriptional Profiles Associated with Marek’s Disease Virus in Bursa and Spleen Lymphocytes Reveal Contrasting Immune Responses during Early Cytolytic Infection. Viruses 2020, 12, 354. [Google Scholar] [CrossRef] [Green Version]
- Spatz, S.J.; Petherbridge, L.; Zhao, Y.; Nair, V. Comparative full-length sequence analysis of oncogenic and vaccine (Rispens) strains of Marek’s disease virus. J. Gen. Virol. 2007, 88, 1080–1096. [Google Scholar] [CrossRef]
- Berthault, C.; Larcher, T.; Hartle, S.; Vautherot, J.F.; Trapp-Fragnet, L.; Denesvre, C. Atrophy of primary lymphoid organs induced by Marek’s disease virus during early infection is associated with increased apoptosis, inhibition of cell proliferation and a severe B-lymphopenia. Vet. Res. 2018, 49, 31. [Google Scholar] [CrossRef] [Green Version]
- Baaten, B.J.; Staines, K.A.; Smith, L.P.; Skinner, H.; Davison, T.F.; Butter, C. Early replication in pulmonary B cells after infection with Marek’s disease herpesvirus by the respiratory route. Viral Immunol. 2009, 22, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Bavananthasivam, J.; Alkie, T.N.; Astill, J.; Abdul-Careem, M.F.; Wootton, S.K.; Behboudi, S.; Yitbarek, A.; Sharif, S. In ovo administration of Toll-like receptor ligands encapsulated in PLGA nanoparticles impede tumor development in chickens infected with Marek’s disease virus. Vaccine 2018, 36, 4070–4076. [Google Scholar] [CrossRef]
- Abdul-Careem, M.F.; Read, L.R.; Parvizi, P.; Thanthrige-Don, N.; Sharif, S. Marek’s disease virus-induced expression of cytokine genes in feathers of genetically defined chickens. Dev. Comp. Immunol. 2009, 33, 618–623. [Google Scholar] [CrossRef]
- Calnek, B.W. Marek’s disease—A model for herpesvirus oncology. Crit. Rev. Microbiol. 1986, 12, 293–320. [Google Scholar] [CrossRef]
- Zhang, Q.; Sun, X.; Wang, T.; Chen, B.; Huang, Y.; Chen, H.; Chen, Q. The Postembryonic Development of the Immunological Barrier in the Chicken Spleens. J. Immunol. Res. 2019, 2019, 6279360. [Google Scholar] [CrossRef] [Green Version]
- Ifrah, M.E.; Perelman, B.; Finger, A.; Uni, Z. The role of the bursa of Fabricius in the immune response to vaccinal antigens and the development of immune tolerance in chicks (Gallus domesticus) vaccinated at a very young age. Poultry science 2017, 96, 51–57. [Google Scholar] [CrossRef]
- Islam, A.F.; Wong, C.W.; Walkden-Brown, S.W.; Colditz, I.G.; Arzey, K.E.; Groves, P.J. Immunosuppressive effects of Marek’s disease virus (MDV) and herpesvirus of turkeys (HVT) in broiler chickens and the protective effect of HVT vaccination against MDV challenge. Avian Pathol. J. W.V.P.A 2002, 31, 449–461. [Google Scholar] [CrossRef]
- Jarosinski, K.W.; Njaa, B.L.; O’Connell, P.H.; Schat, K.A. Pro-inflammatory responses in chicken spleen and brain tissues after infection with very virulent plus Marek’s disease virus. Viral Immunol. 2005, 18, 148–161. [Google Scholar] [CrossRef]
- Paramithiotis, E.; Ratcliffe, M.J. Bursa-dependent subpopulations of peripheral B lymphocytes in chicken blood. Eur. J. Immunol. 1993, 23, 96–102. [Google Scholar] [CrossRef]
- Baigent, S.J.; Smith, L.P.; Currie, R.J.W.; Nair, V.K. Replication kinetics of Marek’s disease vaccine virus in feathers and lymphoid tissues using PCR and virus isolation. J. Gen. Virol. 2005, 86, 2989–2998. [Google Scholar] [CrossRef]
- Calnek, B.W. Pathogenesis of Marek’s disease virus infection. Curr. Top. Microbiol. Immunol. 2001, 255, 25–55. [Google Scholar] [CrossRef]
- Sadigh, Y.; Tahiri-Alaoui, A.; Spatz, S.; Nair, V.; Ribeca, P. Pervasive Differential Splicing in Marek’s Disease Virus can Discriminate CVI-988 Vaccine Strain from RB-1B Very Virulent Strain in Chicken Embryonic Fibroblasts. Viruses 2020, 12, 329. [Google Scholar] [CrossRef] [Green Version]
- Bertzbach, L.D.; Pfaff, F.; Pauker, V.I.; Kheimar, A.M.; Hoper, D.; Hartle, S.; Karger, A.; Kaufer, B.B. The Transcriptional Landscape of Marek’s Disease Virus in Primary Chicken B Cells Reveals Novel Splice Variants and Genes. Viruses 2019, 11, 264. [Google Scholar] [CrossRef]
- Haq, K.; Fear, T.; Ibraheem, A.; Abdul-Careem, M.F.; Sharif, S. Influence of vaccination with CVI988/Rispens on load and replication of a very virulent Marek’s disease virus strain in feathers of chickens. Avian Pathol. J. W.V.P.A 2012, 41, 69–75. [Google Scholar] [CrossRef]
- Bertzbach, L.D.; Harlin, O.; Hartle, S.; Fehler, F.; Vychodil, T.; Kaufer, B.B.; Kaspers, B. IFNalpha and IFNgamma Impede Marek’s Disease Progression. Viruses 2019, 11, 1103. [Google Scholar] [CrossRef] [Green Version]
- Brubaker, S.W.; Bonham, K.S.; Zanoni, I.; Kagan, J.C. Innate immune pattern recognition: A cell biological perspective. Annu. Rev. Immunol. 2015, 33, 257–290. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Liu, Y.; Xu, Z.; Zhang, Y.; Luo, D.; Gao, Y.; Qian, Y.; Bao, C.; Liu, C.; Zhang, Y.; et al. Avian oncogenic herpesvirus antagonizes the cGAS-STING DNA-sensing pathway to mediate immune evasion. PLoS Pathog. 2019, 15, e1007999. [Google Scholar] [CrossRef] [Green Version]
- Gurung, A.; Kamble, N.; Kaufer, B.B.; Pathan, A.; Behboudi, S. Association of Marek’s Disease induced immunosuppression with activation of a novel regulatory T cells in chickens. PLoS Pathog. 2017, 13, e1006745. [Google Scholar] [CrossRef] [Green Version]
- Xing, Z.; Schat, K.A. Inhibitory effects of nitric oxide and gamma interferon on in vitro and in vivo replication of Marek’s disease virus. J. Virol. 2000, 74, 3605–3612. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.; Wang, J.; Liu, W.; Cheng, J.; Xu, J.; Cao, M.; Li, Y. Comparative transcriptome analysis of MDBK cells reveals that BoIFN-gamma augmented host immune responses to bovine herpesvirus 1 infection. Front. Microbiol. 2022, 13, 973278. [Google Scholar] [CrossRef]
- Jud, A.; Kotur, M.; Berger, C.; Gysin, C.; Nadal, D.; Lunemann, A. Tonsillar CD56brightNKG2A+ NK cells restrict primary Epstein-Barr virus infection in B cells via IFN-gamma. Oncotarget 2017, 8, 6130–6141. [Google Scholar] [CrossRef] [Green Version]
- Bavananthasivam, J.; Astill, J.; Matsuyama-Kato, A.; Taha-Abdelaziz, K.; Shojadoost, B.; Sharif, S. Gut microbiota is associated with protection against Marek’s disease virus infection in chickens. Virology 2021, 553, 122–130. [Google Scholar] [CrossRef]
- Bertzbach, L.D.; van Haarlem, D.A.; Hartle, S.; Kaufer, B.B.; Jansen, C.A. Marek’s Disease Virus Infection of Natural Killer Cells. Microorganisms 2019, 7, 588. [Google Scholar] [CrossRef]
- Djeraba, A.; Musset, E.; Bernardet, N.; Le Vern, Y.; Quere, P. Similar pattern of iNOS expression, NO production and cytokine response in genetic and vaccination-acquired resistance to Marek’s disease. Vet. Immunol. Immunopathol. 2002, 85, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Cohen, C.; Corpet, A.; Roubille, S.; Maroui, M.A.; Poccardi, N.; Rousseau, A.; Kleijwegt, C.; Binda, O.; Texier, P.; Sawtell, N.; et al. Promyelocytic leukemia (PML) nuclear bodies (NBs) induce latent/quiescent HSV-1 genomes chromatinization through a PML NB/Histone H3.3/H3.3 Chaperone Axis. PLoS Pathog. 2018, 14, e1007313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, T.S.; Glass, M.; Cole, J.J.; Rather, M.I.; Marsden, M.; Neilson, M.; Brock, C.; Humphreys, I.R.; Everett, R.D.; Adams, P.D. Histone chaperone HIRA deposits histone H3.3 onto foreign viral DNA and contributes to anti-viral intrinsic immunity. Nucleic Acids Res. 2017, 45, 11673–11683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.R.; Munkhjargal, A.; Kim, M.J.; Park, S.Y.; Jung, E.; Ryu, J.H.; Yang, Y.; Lim, J.S.; Kim, Y. The functional roles of PML nuclear bodies in genome maintenance. Mutat. Res. 2018, 809, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Zheng, Y. Role of ND10 nuclear bodies in the chromatin repression of HSV-1. Virol. J. 2016, 13, 62. [Google Scholar] [CrossRef] [Green Version]
- Catez, F.; Picard, C.; Held, K.; Gross, S.; Rousseau, A.; Theil, D.; Sawtell, N.; Labetoulle, M.; Lomonte, P. HSV-1 genome subnuclear positioning and associations with host-cell PML-NBs and centromeres regulate LAT locus transcription during latency in neurons. PLoS Pathog. 2012, 8, e1002852. [Google Scholar] [CrossRef]
- Scherer, M.; Stamminger, T. Emerging Role of PML Nuclear Bodies in Innate Immune Signaling. J. Virol. 2016, 90, 5850–5854. [Google Scholar] [CrossRef] [Green Version]
- Chan, D.; Bennett, P.R.; Lee, Y.S.; Kundu, S.; Teoh, T.G.; Adan, M.; Ahmed, S.; Brown, R.G.; David, A.L.; Lewis, H.V.; et al. Microbial-driven preterm labour involves crosstalk between the innate and adaptive immune response. Nat. Commun. 2022, 13, 975. [Google Scholar] [CrossRef]
- Cardoso Dal Pont, G.; Lee, A.; Bortoluzzi, C.; Farnell, Y.Z.; Gougoulias, C.; Kogut, M.H. Novel model for chronic intestinal inflammation in chickens: (2) Immunologic mechanism behind the inflammatory response. Dev. Comp. Immunol. 2022, 138, 104524. [Google Scholar] [CrossRef]
- Schilling, M.A.; Katani, R.; Memari, S.; Cavanaugh, M.; Buza, J.; Radzio-Basu, J.; Mpenda, F.N.; Deist, M.S.; Lamont, S.J.; Kapur, V. Transcriptional Innate Immune Response of the Developing Chicken Embryo to Newcastle Disease Virus Infection. Front. Genet. 2018, 9, 61. [Google Scholar] [CrossRef]
- Ni Gabhann-Dromgoole, J.; de Chaumont, C.; Shahnazaryan, D.; Smith, S.; Malone, C.; Hassan, J.; De Gascun, C.F.; Jefferies, C.A.; Murphy, C.C. Systemic IL-1beta production as a consequence of corneal HSV-1 infection-contribution to the development of herpes simplex keratitis. Int. J. Ophthalmol. 2019, 12, 1493–1497. [Google Scholar] [CrossRef]
- Dinarello, C.A. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef] [Green Version]
- Rebe, C.; Ghiringhelli, F. Interleukin-1beta and Cancer. Cancers 2020, 12, 1791. [Google Scholar] [CrossRef]
- Karaba, A.H.; Figueroa, A.; Massaccesi, G.; Botto, S.; DeFilippis, V.R.; Cox, A.L. Herpes simplex virus type 1 inflammasome activation in proinflammatory human macrophages is dependent on NLRP3, ASC, and caspase-1. PLoS ONE 2020, 15, e0229570. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, P.; Underwood, G.; Davison, F. Differential cytokine responses following Marek’s disease virus infection of chickens differing in resistance to Marek’s disease. J. Virol. 2003, 77, 762–768. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Careem, M.F.; Hunter, B.D.; Parvizi, P.; Haghighi, H.R.; Thanthrige-Don, N.; Sharif, S. Cytokine gene expression patterns associated with immunization against Marek’s disease in chickens. Vaccine 2007, 25, 424–432. [Google Scholar] [CrossRef]
- Jones, D.; Neff, C.P.; Palmer, B.E.; Stenmark, K.; Nagel, M.A. Varicella zoster virus-infected cerebrovascular cells produce a proinflammatory environment. Neurol. Neuroimmunol. Neuroinflamm. 2017, 4, e382. [Google Scholar] [CrossRef] [Green Version]
- Costa, H.; Nascimento, R.; Sinclair, J.; Parkhouse, R.M. Human cytomegalovirus gene UL76 induces IL-8 expression through activation of the DNA damage response. PLoS Pathog. 2013, 9, e1003609. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhang, J.; Kumar, A.; Zheng, M.; Atherton, S.S.; Yu, F.S. Herpes simplex virus 1 infection induces the expression of proinflammatory cytokines, interferons and TLR7 in human corneal epithelial cells. Immunology 2006, 117, 167–176. [Google Scholar] [CrossRef]
- Engel, A.T.; Selvaraj, R.K.; Kamil, J.P.; Osterrieder, N.; Kaufer, B.B. Marek’s disease viral interleukin-8 promotes lymphoma formation through targeted recruitment of B cells and CD4+ CD25+ T cells. J. Virol. 2012, 86, 8536–8545. [Google Scholar] [CrossRef]
- Nakayama, T.; Shirane, J.; Hieshima, K.; Shibano, M.; Watanabe, M.; Jin, Z.; Nagakubo, D.; Saito, T.; Shimomura, Y.; Yoshie, O. Novel antiviral activity of chemokines. Virology 2006, 350, 484–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bystry, R.S.; Aluvihare, V.; Welch, K.A.; Kallikourdis, M.; Betz, A.G. B cells and professional APCs recruit regulatory T cells via CCL4. Nat. Immunol. 2001, 2, 1126–1132. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | Name | Sequence (5′-3′) |
|---|---|---|---|
| Meq-F | CCCAACAGCCCCTCCAAACAC | Meq-R | CTTCATGGAGTTTGTCTACA |
| ChIFN-β-F | CCTCAACCAGATCCAGCATTAC | ChIFN-β-R | CCCAGGTACAAGCACTGTAGTT |
| ChIFN-γ-F | GCCGCACATCAAACACATATCTG | ChIFN-γ-R | GCGCTGGATTCTCAAGTCGTTC |
| ChPML-F | TCTACCGGCGCATTGTCAG | ChPML-R | TGGGTCTCGAAGCACTTTGT |
| IL-1β-F | CCCGCTTCATCTTCTACCGC | IL-1β-R | GCTTGTAGGTGGCGATGTTG |
| IL-6-F | AAATCCCTCCTCGCCAATCT | IL-6-R | CCCTCACGGTCTTCTCCATAAA |
| IL8L1-F | CCTTCAGCTTTGTGGCAGAC | IL8L1-R | GGAGGAGGTAGGACGTTTTTG |
| CCL4-F | CCAGAATGCTGGTAATCGACG | CCL4-R | GGTGATGAACACAACACCAGC |
| GAPDH-F | AGAACATCATCCCAGCGT | GAPDH-R | AGCCTTCACTACCCTCTTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, B.; Wang, J.; Cao, M.; Jin, H.; Liu, W.; Cheng, J.; Zhou, L.; Xu, J.; Li, Y. Differential Replication and Cytokine Response between Vaccine and Very Virulent Marek’s Disease Viruses in Spleens and Bursas during Latency and Reactivation. Viruses 2023, 15, 6. https://doi.org/10.3390/v15010006
Jiang B, Wang J, Cao M, Jin H, Liu W, Cheng J, Zhou L, Xu J, Li Y. Differential Replication and Cytokine Response between Vaccine and Very Virulent Marek’s Disease Viruses in Spleens and Bursas during Latency and Reactivation. Viruses. 2023; 15(1):6. https://doi.org/10.3390/v15010006
Chicago/Turabian StyleJiang, Bo, Jing Wang, Mengyao Cao, Huan Jin, Wenxiao Liu, Jing Cheng, Linyi Zhou, Jian Xu, and Yongqing Li. 2023. "Differential Replication and Cytokine Response between Vaccine and Very Virulent Marek’s Disease Viruses in Spleens and Bursas during Latency and Reactivation" Viruses 15, no. 1: 6. https://doi.org/10.3390/v15010006