
ADP-Ribosylation and Antiviral Resistance in Plants

 ,
,  ,
, {kind=link}
{kind=link}
Abstract
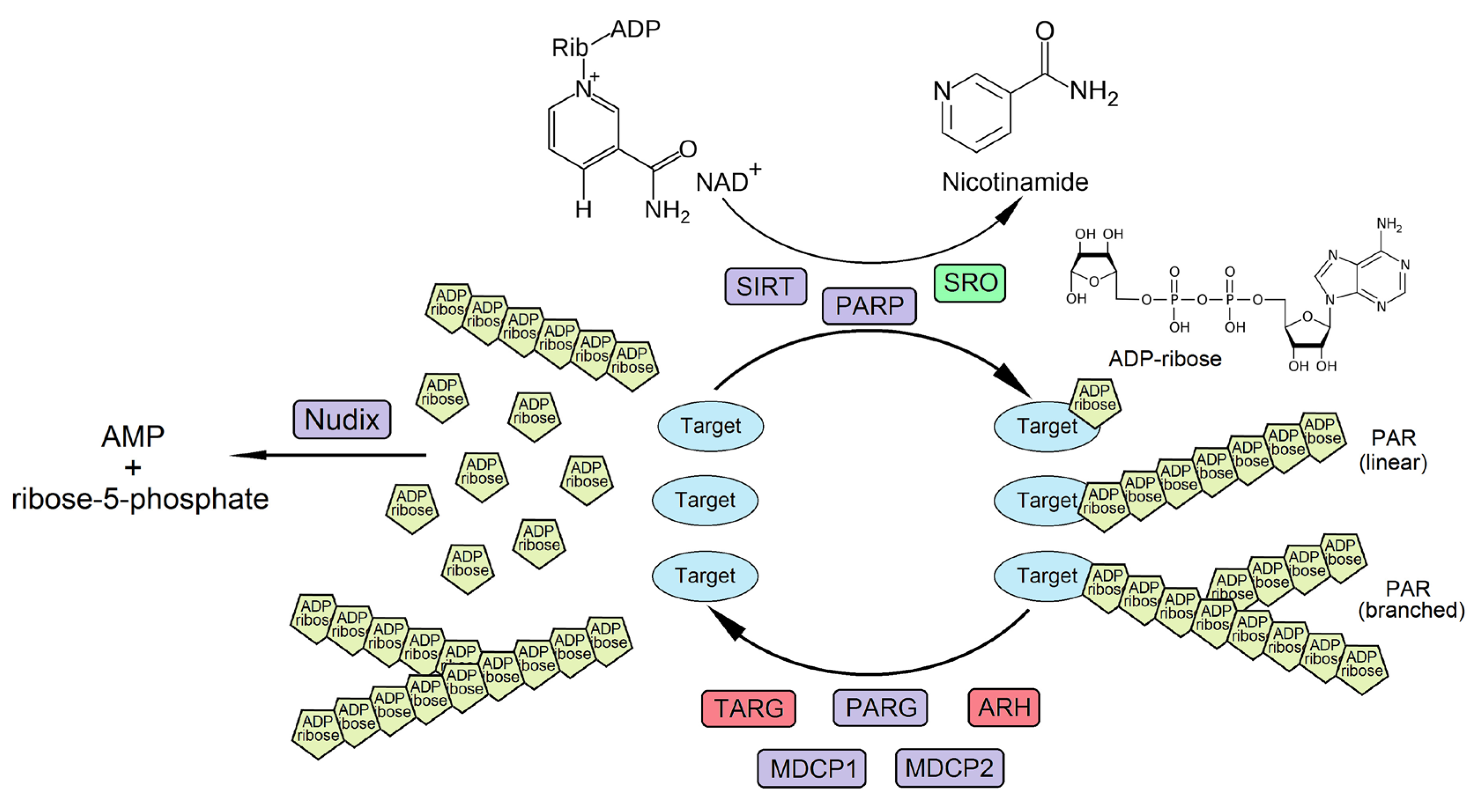
:1. Introduction to ADP-Ribose Metabolism
2. ADP-Ribosylation in Plants
2.1. Major Enzymes
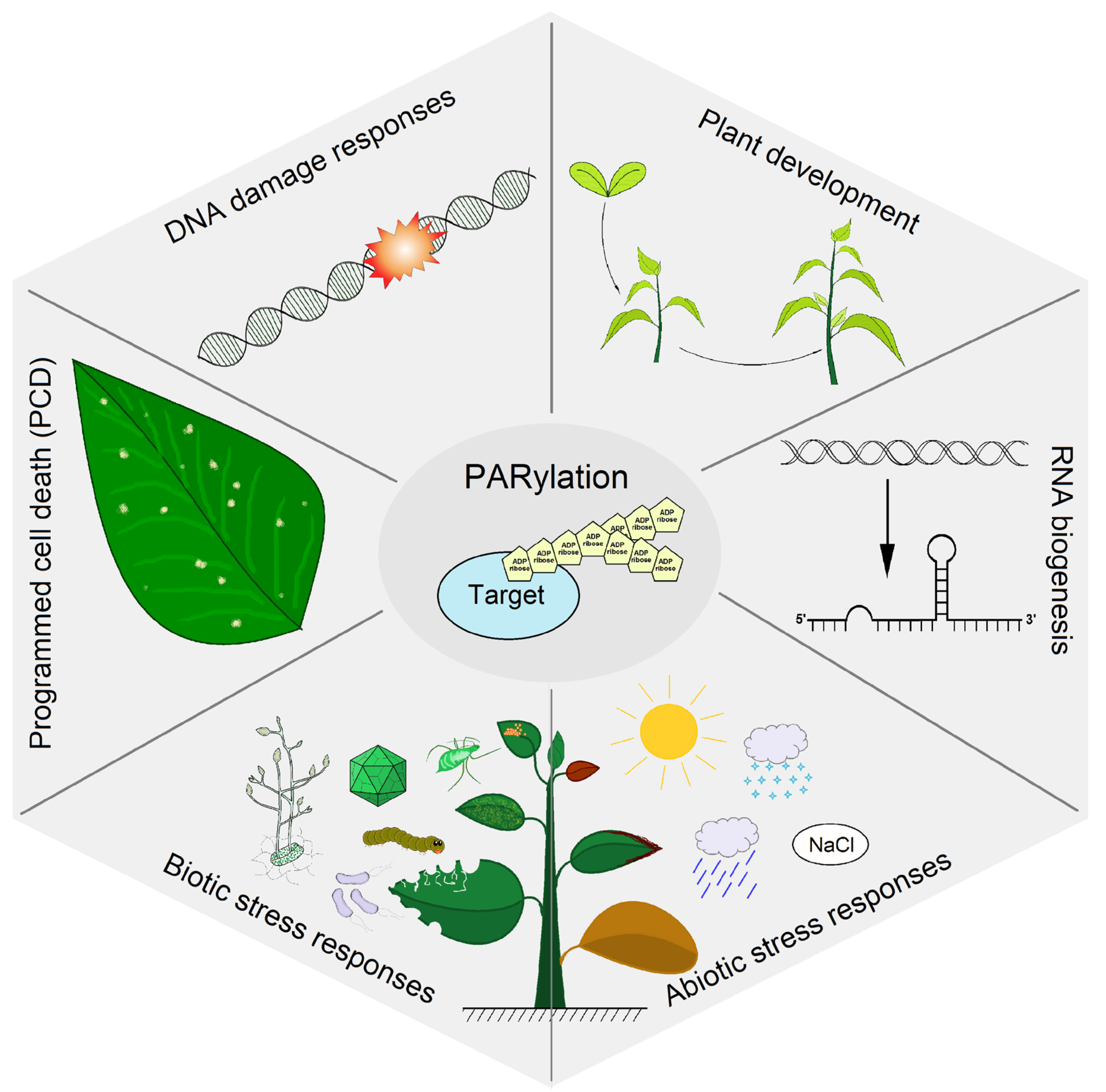
2.2. Biological Functions
2.2.1. DNA Damage Responses
2.2.2. Plant Development
2.2.3. RNA Biogenesis
2.2.4. Abiotic Stress Responses
2.2.5. Plant Immunity
2.2.6. Possible Off-Target Effects of PARP Inhibitors
3. PARylation and Plant–Virus Interactions: What Is Already Known?
4. PARylation and Plant–Virus Interactions: What Is Next?
- CBs and their signature protein, coilin, have been shown to affect a variety of interactions between host plants and viruses that have RNA or DNA genomes. Moreover, the effects of coilin on these interactions are manifested differently: coilin contributes to plant defence against some viruses, but in contrast may serve to increase virus pathogenicity in other viruses. These findings show that interactions with coilin (or CBs) may involve diverse mechanisms with different viruses, and that these mechanisms act at different phases of virus infection [44,45,46]. Taking into account that CBs/coilin may trigger responses to virus infection via interaction with PARP and modulation of its activity, it would be conceivable to select a wide range of DNA and RNA viruses to address their ability to multiply in parp, parg or nudix knockout or knockdown plants.
- In nature, plants are usually exposed to diverse environmental stresses, which may modulate plant–virus interactions [48]. It is suggested that responses to abiotic stresses and virus infections may be integrated in a specific consolidated network which controls plant sensitivity to multiple stresses. Given that PARylation process may differentially affect various biotic and abiotic stresses, investigation of effects of combined stress caused by viruses and major environmental cues on parp, parg or nudix knockout or knockdown plants must be carried out. In addition to using genetic approaches, PARP inhibitors may also be exploited to explore interactions of these different molecular switches, but with the precautions discussed above.
- To elucidate the mechanisms underlying the cross-talk between PARylation and virus infection outcomes, analysis looking at expression level (transcript levels and protein levels) and activity of PARylation-related enzymes (PARP, SRO, PARG etc) and PAR itself should be performed during the infection process. It would also be important to look at possible relocalisation of PARylation related proteins potentially induced by various viruses, as significantly altered distribution (including perhaps sequestration to defined cellular regions) may greatly affect PARylation efficiency. It is well known, for example, that the nucleolus can act as a sequestration centre which may capture and detain various cellular factors [49].
- In mammalian model systems, it appears that MARylation is required for host cells to respond to viral infection. It is, therefore, conceivable to examine the distinctive role of MARylation (compared with PARylation) in plant–virus interactions.
- It has recently been shown that nucleic acids (DNA and RNA) may be directly ADPRylated. Therefore, future studies should examine whether such modifications can occur with plant viral RNAs or DNAs and investigate the biological relevance of such process.
- It would also be critically important to identify PARylation targets among virus components (proteins, nucleic acids) and host signalling molecules involved in plant responses to virus attack or to combined stresses. Application of systems biology approaches to analyse the data obtained will allow the identification of target genes and regulatory hubs which could be exploited to develop improved plant resistance to biotic and environmental stresses.
- On a practical level, genetic alteration of the PARylation (and possibly MARylation) process (by affecting PARP and PARG proteins) in crops may be achieved either by CRISPR-Cas technology (gene knock out or knock in or by alteration of a specific gene sequence) [50] or by RNA interference (RNAi)-based methods [51]. It is worth noting that for modification of genes critical to plant survival, CRISPR-Cas may not be ideal, as it would permanently affect those genes, possibly interfering with plant growth and development. Alternatively, spray induced gene silencing (SIGS) RNAi techniques which employs external treatment of plants with dsRNA as a trigger for RNAi, may be preferable for those genes since it would allow flexible reversible modification of gene expression in real time without compromising their essential function over the lifecycle of the plant. In animals MARylation often mediates host defence responses. Inhibitors of hydrolases (such as PARG) are therefore being pursued as a potential antiviral therapeutic strategy. A similar strategy could be applied for crop protection. Despite the certain off-target effects of known PARP/PARG inhibitors, development of the next generation inhibitors is required to enhance plant stress tolerance resulting in improved growth and yield.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rissel, D.; Peiter, E. Poly(ADP-Ribose) Polymerases in Plants and Their Human Counterparts: Parallels and Peculiarities. Int. J. Mol. Sci. 2019, 20, 1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challa, S.; Stokes, M.S.; Kraus, W.L. MARTs and MARylation in the Cytosol: Biological Functions, Mechanisms of Action, and Therapeutic Potential. Cells 2021, 10, 313. [Google Scholar] [CrossRef] [PubMed]
- Maluchenko, N.V.; Koshkina, D.O.; Feofanov, A.V.; Studitsky, V.M.; Kirpichnikov, M.P. Poly(ADP-Ribosyl) Code Functions. Acta Nat. 2021, 13, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Briggs, A.G.; Bent, A.F. Poly(ADP-Ribosyl)Ation in Plants. Trends Plant Sci. 2011, 16, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Kraus, W.L. On PAR with PARP: Cellular Stress Signaling through Poly(ADP-Ribose) and PARP-1. Genes Dev. 2012, 26, 417–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, A.J.; Yu, C.; Petukhova, N.V.; Kalinina, N.O.; Chen, J.; Taliansky, M.E. Cajal Bodies and Their Role in Plant Stress and Disease Responses. RNA Biol. 2017, 14, 779–790. [Google Scholar] [CrossRef] [Green Version]
- Kalinina, N.O.; Makarova, S.; Makhotenko, A.; Love, A.J.; Taliansky, M. The Multiple Functions of the Nucleolus in Plant Development, Disease and Stress Responses. Front. Plant Sci. 2018, 9, 132. [Google Scholar] [CrossRef]
- Kotova, E.; Jarnik, M.; Tulin, A.V. Poly (ADP-Ribose) Polymerase 1 Is Required for Protein Localization to Cajal Body. PLoS Genet. 2009, 5, e1000387. [Google Scholar] [CrossRef] [Green Version]
- Suskiewicz, M.J.; Palazzo, L.; Hughes, R.; Ahel, I. Progress and Outlook in Studying the Substrate Specificities of PARPs and Related Enzymes. FEBS J. 2021, 288, 2131–2142. [Google Scholar] [CrossRef]
- Weixler, L.; Schäringer, K.; Momoh, J.; Lüscher, B.; Feijs, K.L.H.; Žaja, R. ADP-Ribosylation of RNA and DNA: From in Vitro Characterization to in Vivo Function. Nucleic Acids Res. 2021, 49, 3634–3650. [Google Scholar] [CrossRef]
- Kim, D.-S.; Challa, S.; Jones, A.; Kraus, W.L. PARPs and ADP-Ribosylation in RNA Biology: From RNA Expression and Processing to Protein Translation and Proteostasis. Genes Dev. 2020, 34, 302–320. [Google Scholar] [CrossRef] [PubMed]
- Mikolčević, P.; Hloušek-Kasun, A.; Ahel, I.; Mikoč, A. ADP-Ribosylation Systems in Bacteria and Viruses. Comput. Struct. Biotechnol. J. 2021, 19, 2366–2383. [Google Scholar] [CrossRef] [PubMed]
- Malgras, M.; Garcia, M.; Jousselin, C.; Bodet, C.; Lévêque, N. The Antiviral Activities of Poly-ADP-Ribose Polymerases. Viruses 2021, 13, 582. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Wolf, J.J.; Sun, C.; Xu, M.; Studstill, C.J.; Chen, J.; Ngo, H.; Zhu, H.; Hahm, B. PARP1 Enhances Influenza A Virus Propagation by Facilitating Degradation of Host Type I Interferon Receptor. J. Virol. 2020, 94, e01572-19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Guo, J.; Chen, Q. Role of PARP-1 in Human Cytomegalovirus Infection and Functional Partners Encoded by This Virus. Viruses 2022, 14, 2049. [Google Scholar] [CrossRef]
- Vainonen, J.P.; Shapiguzov, A.; Vaattovaara, A.; Kangasjärvi, J. Plant PARPs, PARGs and PARP-like Proteins. Curr. Protein Pept. Sci. 2016, 17, 713–723. [Google Scholar] [CrossRef]
- Gu, Z.; Pan, W.; Chen, W.; Lian, Q.; Wu, Q.; Lv, Z.; Cheng, X.; Ge, X. New Perspectives on the Plant PARP Family: Arabidopsis PARP3 Is Inactive, and PARP1 Exhibits Predominant Poly (ADP-Ribose) Polymerase Activity in Response to DNA Damage. BMC Plant Biol. 2019, 19, 364. [Google Scholar] [CrossRef] [Green Version]
- Szućko, I. Sirtuins: Not Only Animal Proteins. Acta Physiol. Plant. 2016, 38, 237. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Feng, B.; Yan, Y.; Zhang, C.; Kim, J.H.; Xu, L.; Rack, J.G.M.; Wang, Y.; Jang, J.-C.; Ahel, I.; et al. Noncanonical Mono(ADP-Ribosyl)Ation of Zinc Finger SZF Proteins Counteracts Ubiquitination for Protein Homeostasis in Plant Immunity. Mol. Cell 2021, 81, 4591–4604. [Google Scholar] [CrossRef]
- Vogt, S.; Feijs, K.; Hosch, S.; De Masi, R.; Lintermann, R.; Loll, B.; Wirthmueller, L. The Superior Salinity Tolerance of Bread Wheat Cultivar Shanrong No. 3 Is Unlikely to Be Caused by Elevated Ta-Sro1 Poly-(ADP-Ribose) Polymerase Activity. Plant Cell 2022, 34, 4130–4137. [Google Scholar] [CrossRef]
- Liu, S.-W.; Liu, S.-P.; Wang, W.-L.; Wang, M.; Wang, M.; Xia, G.-M. Whether the ADP-Ribosyltransferase Activity of Ta-Sro1, a Noncanonical PARP Protein, Contributes to Its Function in Salinity-Stress Tolerance? BioRxiv 2022, 24, 505095. [Google Scholar] [CrossRef]
- Yoshimura, K.; Shigeoka, S. Versatile Physiological Functions of the Nudix Hydrolase Family in Arabidopsis. Biosci. Biotechnol. Biochem. 2015, 79, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Yu, X. Functions of PARylation in DNA Damage Repair Pathways. Genom. Proteom. Bioinform. 2016, 14, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talhaoui, I.; Lebedeva, N.A.; Zarkovic, G.; Saint-Pierre, C.; Kutuzov, M.M.; Sukhanova, M.V.; Matkarimov, B.T.; Gasparutto, D.; Saparbaev, M.K.; Lavrik, O.I.; et al. Poly(ADP-Ribose) Polymerases Covalently Modify Strand Break Termini in DNA Fragments in Vitro. Nucleic Acids Res. 2016, 44, 9279–9295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eleazer, R.; Fondufe-Mittendorf, Y.N. The Multifaceted Role of PARP1 in RNA Biogenesis. WIREs RNA 2021, 12, e1617. [Google Scholar] [CrossRef]
- Thomas, C.; Ji, Y.; Wu, C.; Datz, H.; Boyle, C.; MacLeod, B.; Patel, S.; Ampofo, M.; Currie, M.; Harbin, J.; et al. Hit and Run versus Long-Term Activation of PARP-1 by Its Different Domains Fine-Tunes Nuclear Processes. Proc. Natl. Acad. Sci. USA 2019, 116, 9941–9946. [Google Scholar] [CrossRef] [Green Version]
- Singhal, P.; Jan, A.T.; Azam, M.; Haq, Q.M.R. Plant Abiotic Stress: A Prospective Strategy of Exploiting Promoters as Alternative to Overcome the Escalating Burden. Front. Life Sci. 2016, 9, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Block, M.D.; Verduyn, C.; Brouwer, D.D.; Cornelissen, M. Poly(ADP-Ribose) Polymerase in Plants Affects Energy Homeostasis, Cell Death and Stress Tolerance. Plant J. 2005, 41, 95–106. [Google Scholar] [CrossRef]
- Gomez-Cadenas, A.; Vives, V.; Zandalinas, S.I.; Manzi, M.; Sanchez-Perez, A.M.; Perez-Clemente, R.M.; Arbona, V. Abscisic Acid: A Versatile Phytohormone in Plant Signaling and Beyond. Curr. Protein Pept. Sci. 2015, 16, 413–434. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, J.-M. Plant Immunity Triggered by Microbial Molecular Signatures. Mol. Plant 2010, 3, 783–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, D.; Arguez, M.A.; He, P.; Bent, A.F.; Song, J. Coordinated Regulation of Plant Immunity by Poly(ADP-Ribosyl)Ation and K63-Linked Ubiquitination. Mol. Plant 2021, 14, 2088–2103. [Google Scholar] [CrossRef] [PubMed]
- Adams-Phillips, L.; Wan, J.; Tan, X.; Dunning, F.M.; Meyers, B.C.; Michelmore, R.W.; Bent, A.F. Discovery of ADP-Ribosylation and Other Plant Defense Pathway Elements Through Expression Profiling of Four Different Arabidopsis–Pseudomonas R-Avr Interactions. Mol. Plant-Microbe Interact. 2008, 21, 646–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams-Phillips, L.; Briggs, A.G.; Bent, A.F. Disruption of Poly(ADP-Ribosyl)Ation Mechanisms Alters Responses of Arabidopsis to Biotic Stress. Plant Physiol. 2010, 152, 267–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briggs, A.G.; Adams-Phillips, L.C.; Keppler, B.D.; Zebell, S.G.; Arend, K.C.; Apfelbaum, A.A.; Smith, J.A.; Bent, A.F. A Transcriptomics Approach Uncovers Novel Roles for Poly(ADP-Ribosyl)Ation in the Basal Defense Response in Arabidopsis Thaliana. PLoS ONE 2017, 12, e0190268. [Google Scholar] [CrossRef] [Green Version]
- Vatta, M.; Lyons, B.; Heney, K.A.; Lidster, T.; Merrill, A.R. Mapping the DNA-Binding Motif of Scabin Toxin, a Guanine Modifying Enzyme from Streptomyces Scabies. Toxins 2021, 13, 55. [Google Scholar] [CrossRef]
- Keppler, B.D.; Song, J.; Nyman, J.; Voigt, C.A.; Bent, A.F. 3-Aminobenzamide Blocks MAMP-Induced Callose Deposition Independently of Its Poly(ADPribosyl)Ation Inhibiting Activity. Front. Plant Sci. 2018, 9, 1907. [Google Scholar] [CrossRef]
- Sekine, K.-T.; Kawakami, S.; Hase, S.; Kubota, M.; Ichinose, Y.; Shah, J.; Kang, H.-G.; Klessig, D.F.; Takahashi, H. High Level Expression of a Virus Resistance Gene, RCY1, Confers Extreme Resistance to Cucumber Mosaic Virus in Arabidopsis Thaliana. Mol. Plant-Microbe Interact. 2008, 21, 1398–1407. [Google Scholar] [CrossRef] [Green Version]
- Glushkevich, A.; Spechenkova, N.; Fesenko, I.; Knyazev, A.; Samarskaya, V.; Kalinina, N.O.; Taliansky, M.; Love, A.J. Transcriptomic Reprogramming, Alternative Splicing and RNA Methylation in Potato (Solanum Tuberosum L.) Plants in Response to Potato Virus Y Infection. Plants 2022, 11, 635. [Google Scholar] [CrossRef]
- Samarskaya, V.O.; Spechenkova, N.; Markin, N.; Suprunova, T.P.; Zavriev, S.K.; Love, A.J.; Kalinina, N.O.; Taliansky, M. Impact of Exogenous Application of Potato Virus Y-Specific DsRNA on RNA Interference, Pattern-Triggered Immunity and Poly(ADP-Ribose) Metabolism. Int. J. Mol. Sci. 2022, 23, 7915. [Google Scholar] [CrossRef]
- Niehl, A.; Wyrsch, I.; Boller, T.; Heinlein, M. Double-Stranded RNAs Induce a Pattern-Triggered Immune Signaling Pathway in Plants. New Phytol. 2016, 211, 1008–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Ryabov, E.V.; Kalinina, N.O.; Rakitina, D.V.; Gillespie, T.; MacFarlane, S.; Haupt, S.; Brown, J.W.; Taliansky, M. Cajal Bodies and the Nucleolus Are Required for a Plant Virus Systemic Infection. EMBO J. 2007, 26, 2169–2179. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; MacFarlane, S.; Kalinina, N.O.; Rakitina, D.V.; Ryabov, E.V.; Gillespie, T.; Haupt, S.; Brown, J.W.S.; Taliansky, M. Interaction of a Plant Virus-Encoded Protein with the Major Nucleolar Protein Fibrillarin Is Required for Systemic Virus Infection. Proc. Natl. Acad. Sci. USA 2007, 104, 11115–11120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, J.; Love, A.J.; Makarova, S.S.; Kalinina, N.O.; Harrison, B.D.; Taliansky, M.E. Coilin, the Signature Protein of Cajal Bodies, Differentially Modulates the Interactions of Plants with Viruses in Widely Different Taxa. Nucleus 2014, 5, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J.; Yu, C.; Makhotenko, A.V.; Makarova, S.S.; Love, A.J.; Kalinina, N.O.; MacFarlane, S.; Chen, J.; Taliansky, M.E. Interaction of a Plant Virus Protein with the Signature Cajal Body Protein Coilin Facilitates Salicylic Acid-Mediated Plant Defence Responses. New Phytol. 2019, 224, 439–453. [Google Scholar] [CrossRef]
- Ding, Y.; Lozano-Durán, R. The Cajal Body in Plant-Virus Interactions. Viruses 2020, 12, 250. [Google Scholar] [CrossRef] [Green Version]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging Infectious Diseases of Plants: Pathogen Pollution, Climate Change and Agrotechnology Drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef]
- Makarova, S.; Makhotenko, A.; Spechenkova, N.; Love, A.J.; Kalinina, N.O.; Taliansky, M. Interactive Responses of Potato (Solanum Tuberosum L.) Plants to Heat Stress and Infection With Potato Virus Y. Front. Microbiol. 2018, 9, 2582. [Google Scholar] [CrossRef] [Green Version]
- Audas, T.E.; Jacob, M.D.; Lee, S. The Nucleolar Detention Pathway. Cell Cycle 2012, 11, 2059–2062. [Google Scholar] [CrossRef] [Green Version]
- Kalinina, N.O.; Khromov, A.; Love, A.J.; Taliansky, M.E. CRISPR Applications in Plant Virology: Virus Resistance and Beyond. Phytopathology 2020, 110, 18–28. [Google Scholar] [CrossRef]
- Taliansky, M.; Samarskaya, V.; Zavriev, S.K.; Fesenko, I.; Kalinina, N.O.; Love, A.J. RNA-Based Technologies for Engineering Plant Virus Resistance. Plants 2021, 10, 82. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spechenkova, N.; Kalinina, N.O.; Zavriev, S.K.; Love, A.J.; Taliansky, M. ADP-Ribosylation and Antiviral Resistance in Plants. Viruses 2023, 15, 241. https://doi.org/10.3390/v15010241
Spechenkova N, Kalinina NO, Zavriev SK, Love AJ, Taliansky M. ADP-Ribosylation and Antiviral Resistance in Plants. Viruses. 2023; 15(1):241. https://doi.org/10.3390/v15010241
Chicago/Turabian StyleSpechenkova, Nadezhda, Natalya O. Kalinina, Sergey K. Zavriev, Andrew J. Love, and Michael Taliansky. 2023. "ADP-Ribosylation and Antiviral Resistance in Plants" Viruses 15, no. 1: 241. https://doi.org/10.3390/v15010241