
Integrated Analysis of MicroRNA and mRNA Expression Profiles in the Fat Bodies of MbMNPV-Infected Helicoverpa armigera
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects and Viruses
2.2. Bioassay and Sample Collection
2.3. Identification of H. armigera Infected with MbMNPV
2.4. RNA Extraction and Quality Control
2.5. Library Preparation and Sequencing
2.6. MiRNAs Sequencing and Analysis
2.7. Construction of DEmiRNA–mRNA Regulatory Network
2.8. Validation of miRNA and mRNA by Real-Time Quantitative PCR
2.9. Synthesis of dsRNA, RNA Interference and Viral Infection
2.10. Data Analysis
3. Results
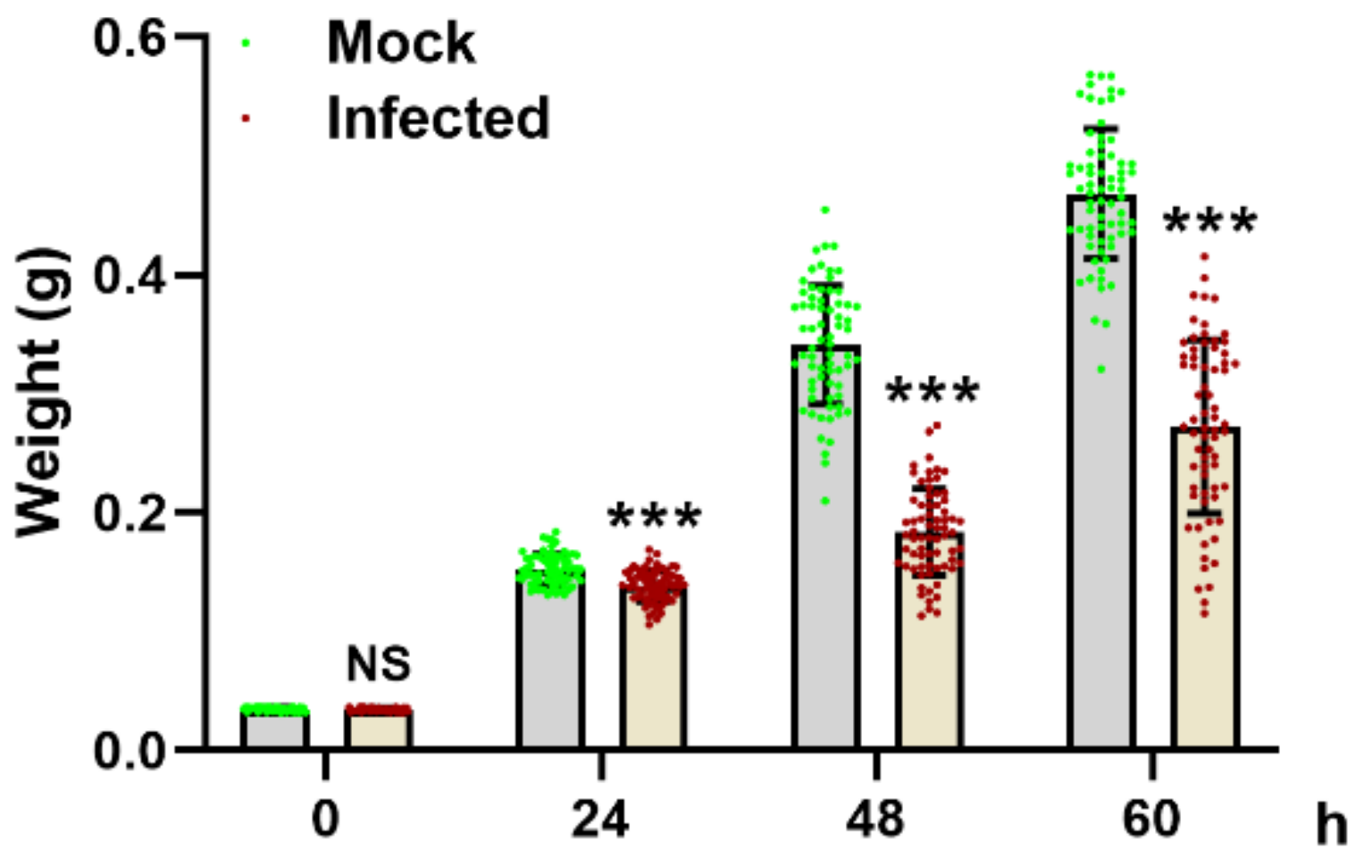
3.1. MbMNPV Infection Caused Significant Reduction in Larval Wet Weight
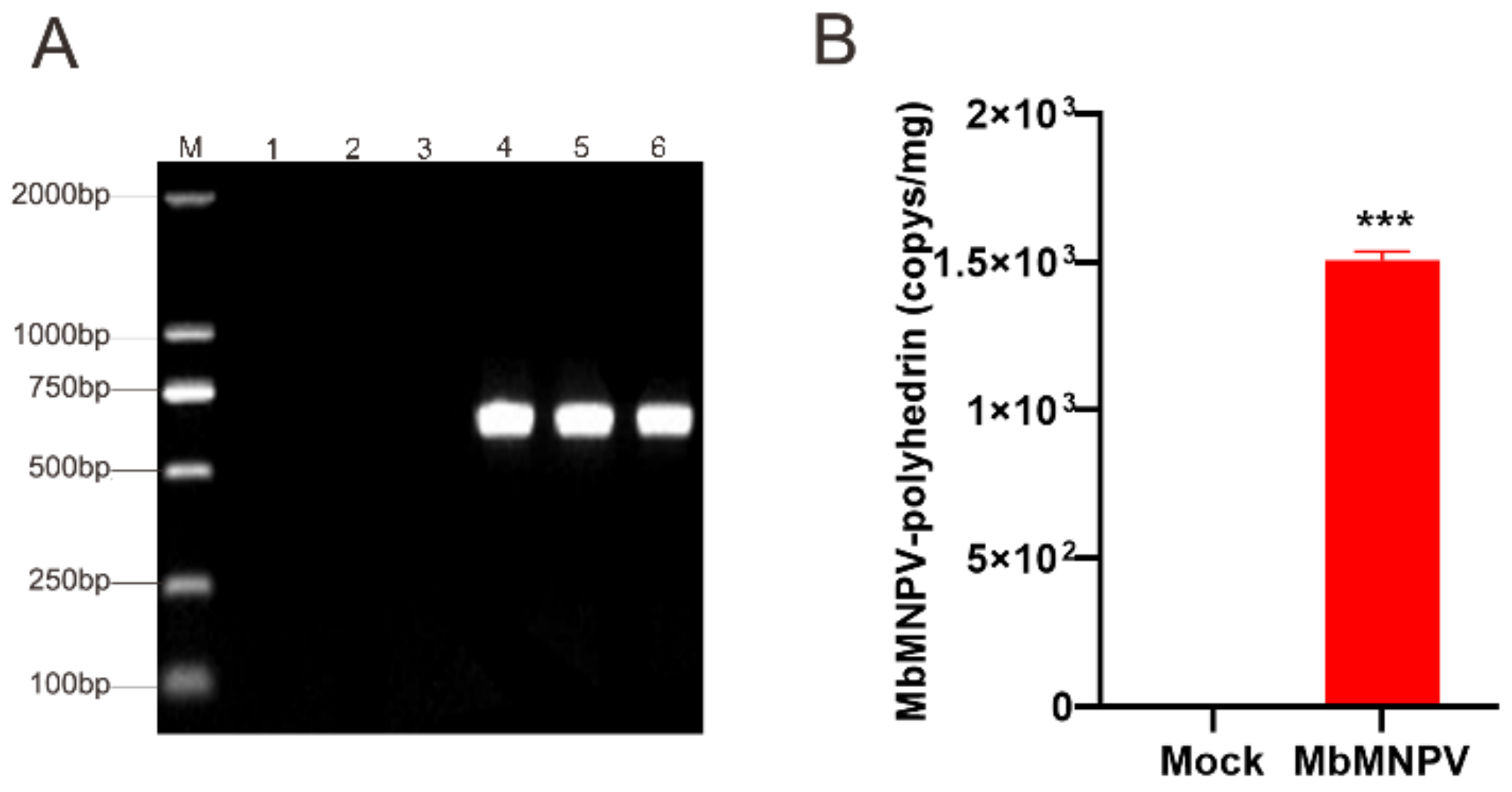
3.2. Verification of the MbMNPV-Infected H. armigera
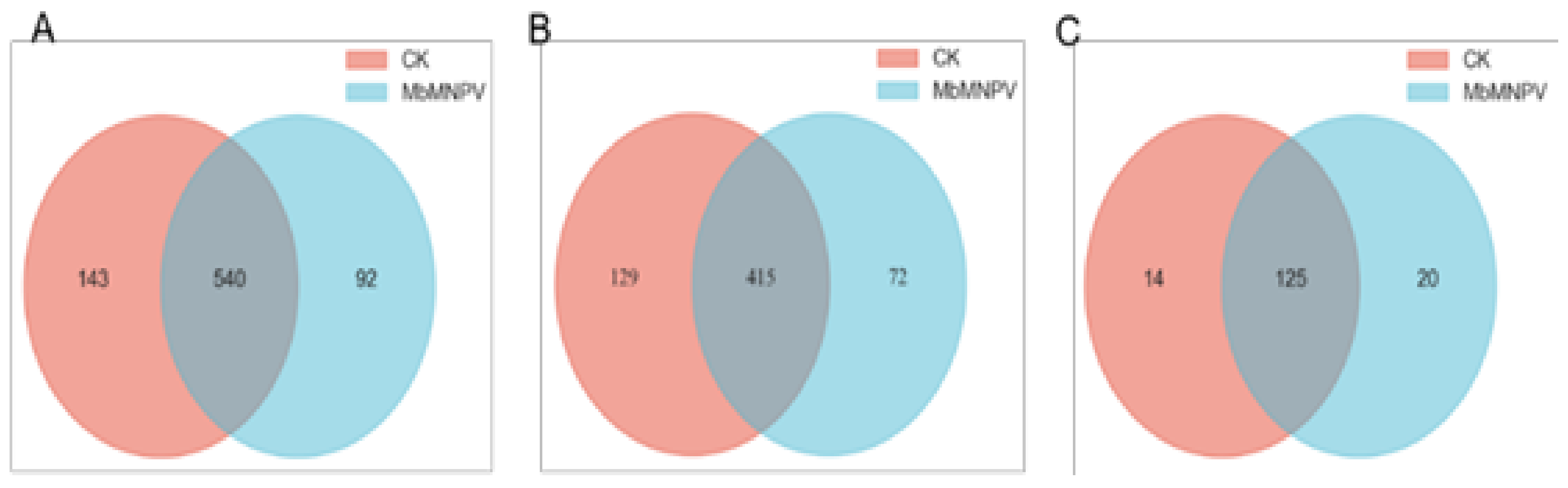
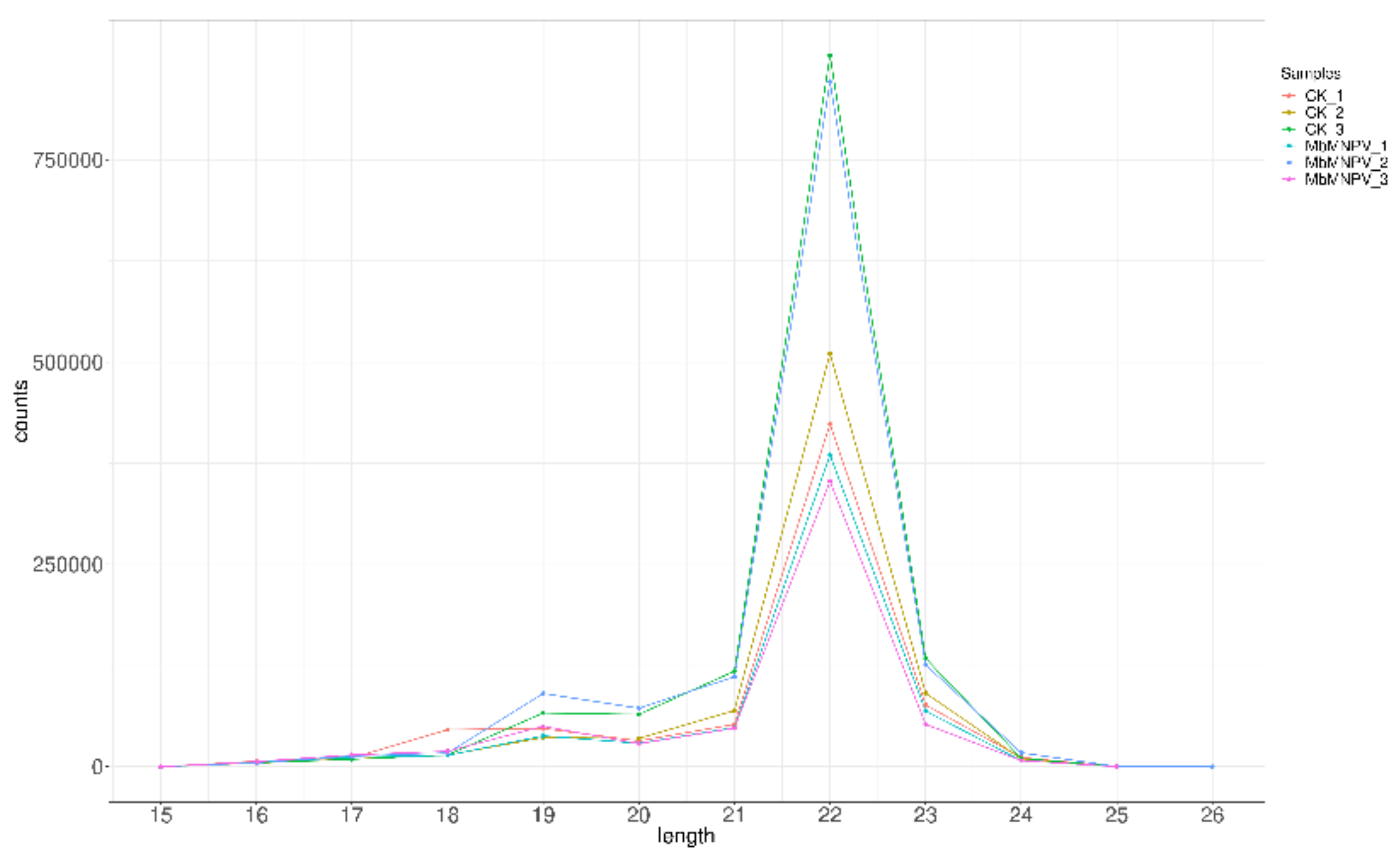
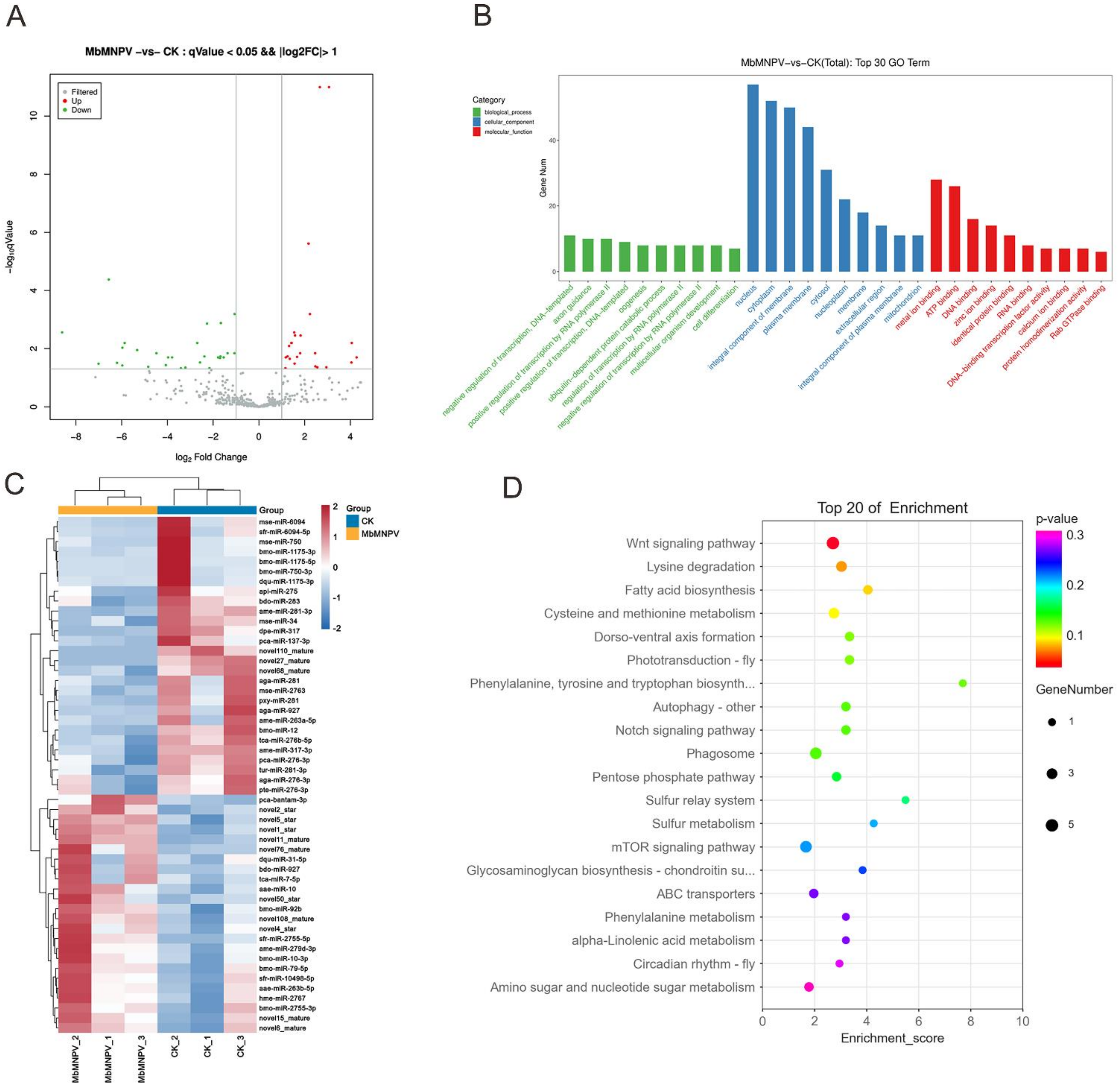
3.3. MiRNA Sequencing and Data Analysis
3.4. Functional Analysis for Targets of DEmiRNAs
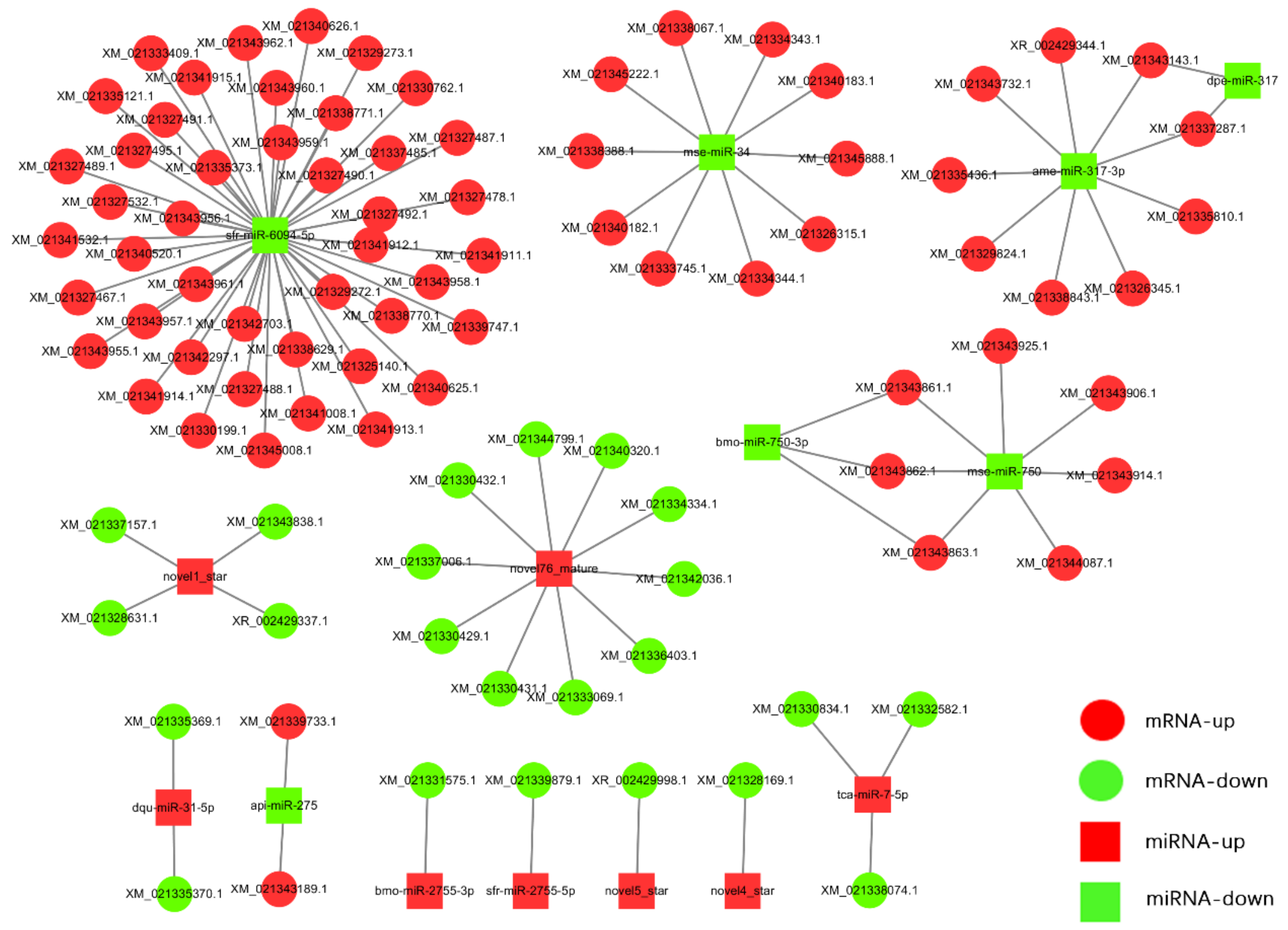
3.5. MiRNA Target Prediction and miRNA–mRNA Interaction Network
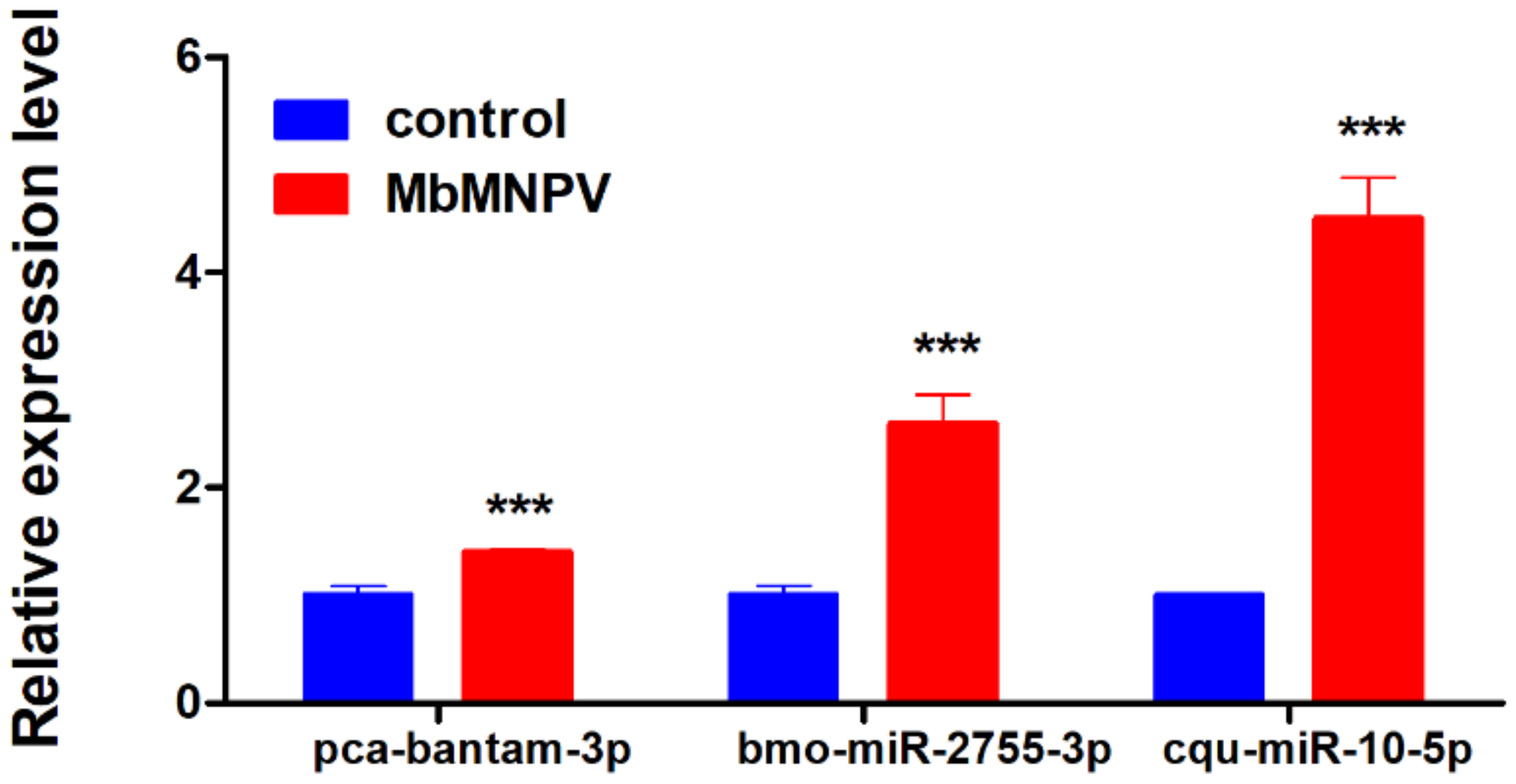
3.6. Verification of DEmiRNAs by Real-Time Quantitative PCR
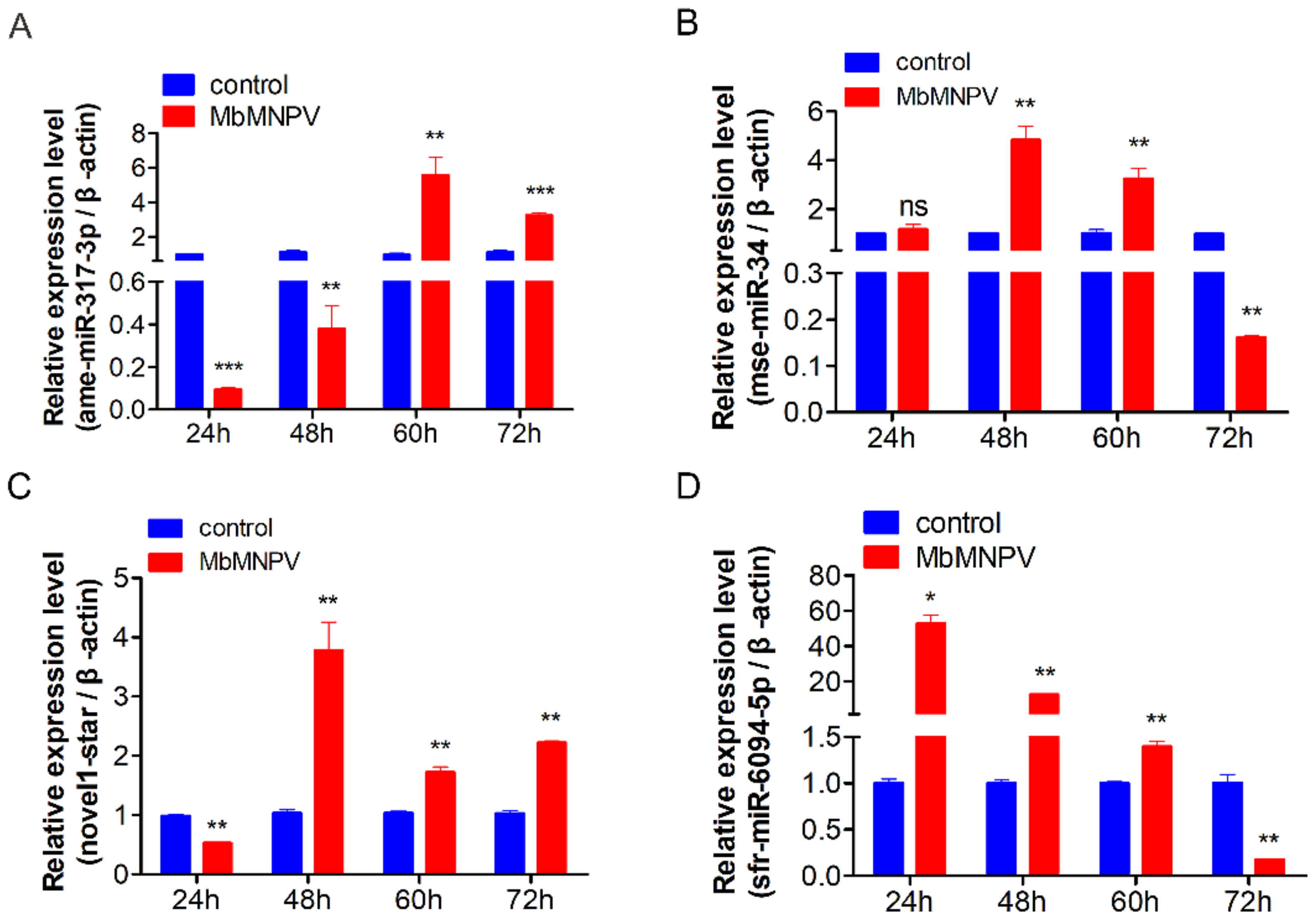
3.7. Relative Expression of miRNAs in H. armigera Fat Body
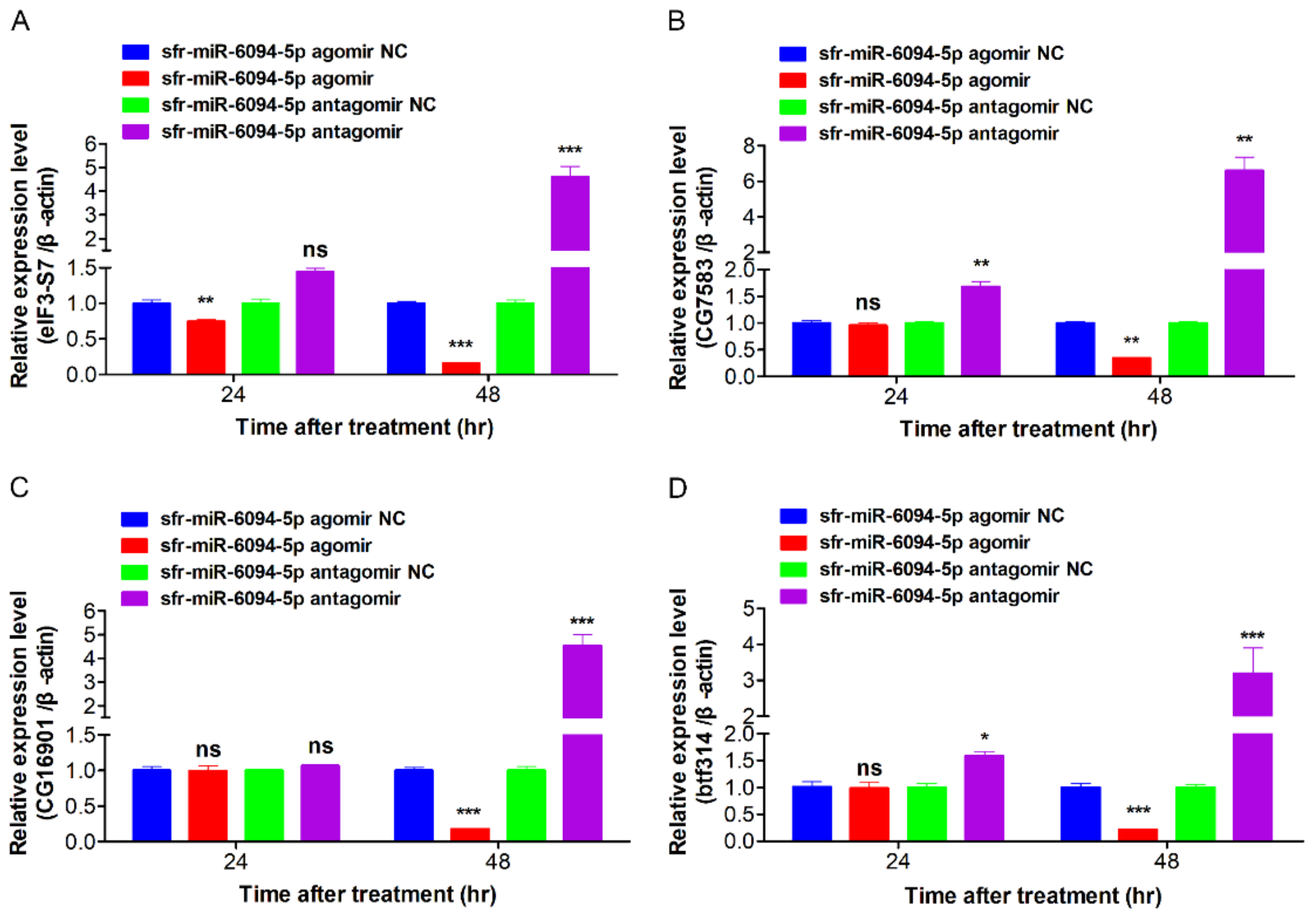
3.8. DEmiRNAs Inhibited Expression of Predicted Target Genes
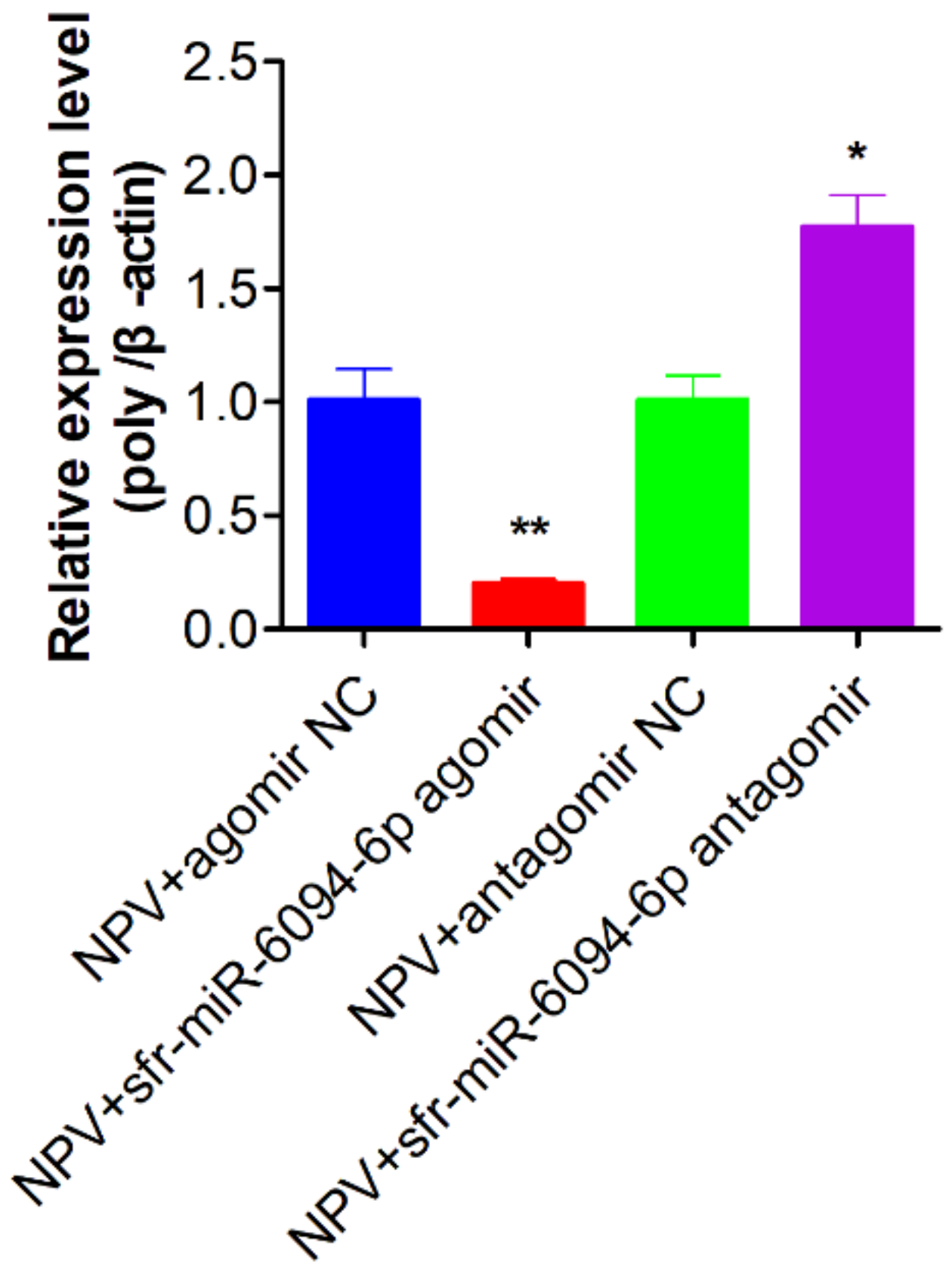
3.9. The Inhibition of sfr-miR-6094-5p on MbMNPV Infection
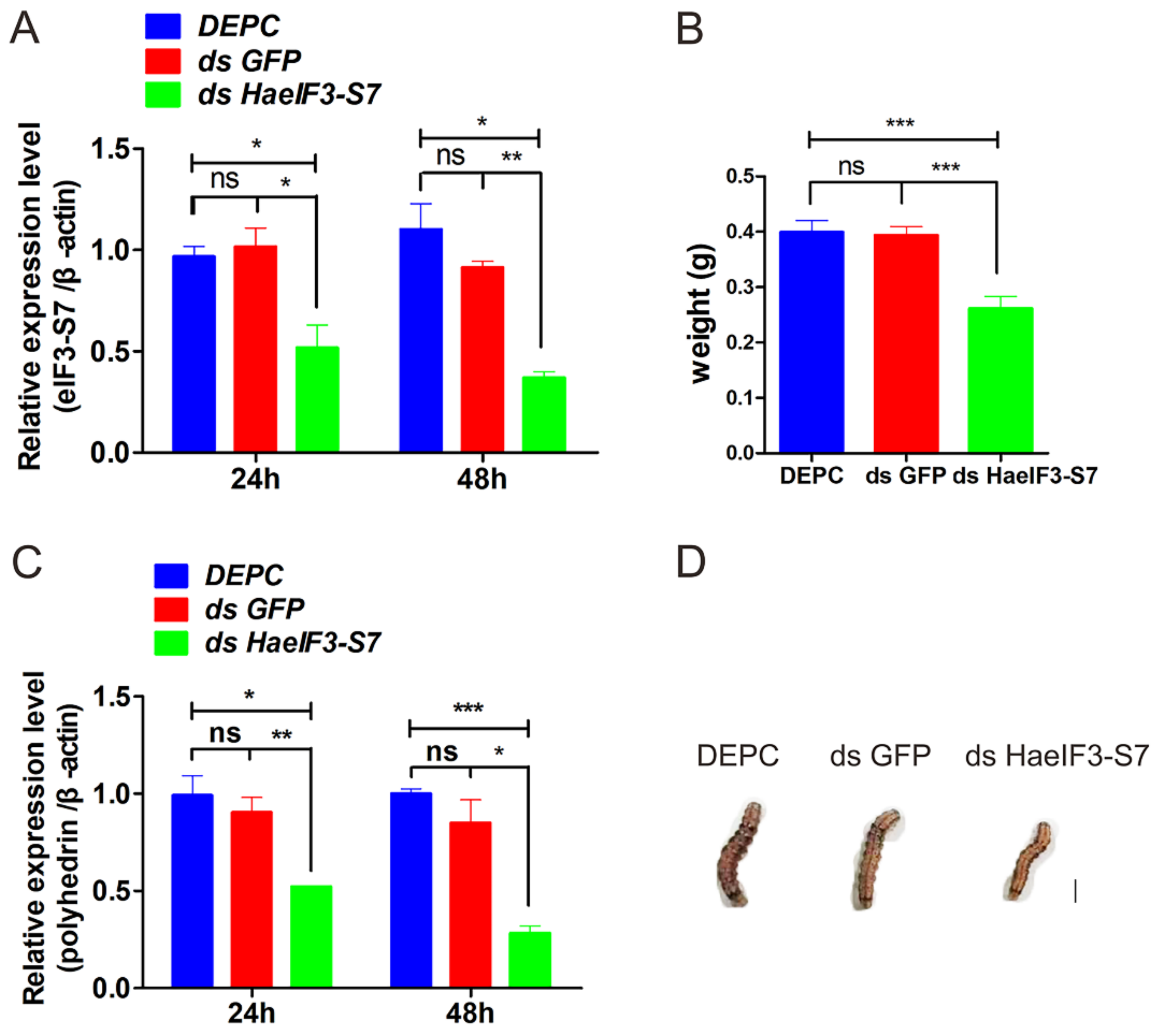
3.10. Knockdown of HaeIF3-S7 Inhibited the MbMNPV Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmad, M.; Rasool, B.; Ahmad, M.; Russell, D.A. Resistance and Synergism of Novel Insecticides in Field Populations of Cotton Bollworm Helicoverpa armigera (Lepidoptera: Noctuidae) in Pakistan. J. Econ. Entomol. 2019, 112, 859–871. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Qiu, X.; Ren, X.; Fang, N.; Wang, K. Resistance selection and biochemical characterization of spinosad resistance in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2009, 95, 90–94. [Google Scholar] [CrossRef]
- Jehle, J.A.; Lange, M.; Wang, H.; Hu, Z.; Wang, Y.; Hauschild, R. Molecular identification and phylogenetic analysis of baculoviruses from Lepidoptera. Virology 2006, 346, 180–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, C.J.; Hirst, M.L.; Cory, J.S.; Entwistle, P.F. Risk Assessment Studies: Detailed Host Range Testing of Wild-Type Cabbage Moth, Mamestra brassicae (Lepidoptera: Noctuidae), Nuclear Polyhedrosis Virus. Appl. Environ. Microbiol. 1990, 56, 2704–2710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H. RNA Interference to Knock Down Gene Expression. Methods Mol. Biol. 2018, 1706, 293–302. [Google Scholar] [CrossRef]
- Zhu, K.Y.; Palli, S.R. Mechanisms, Applications, and Challenges of Insect RNA Interference. Annu. Rev. Entomol. 2020, 65, 293–311. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Santos, D.; Wynant, N.; Van den Brande, S.; Verdonckt, T.W.; Mingels, L.; Peeters, P.; Kolliopoulou, A.; Swevers, L.; Vanden Broeck, J. Insights into RNAi-based antiviral immunity in Lepidoptera: Acute and persistent infections in Bombyx mori and Trichoplusia ni cell lines. Sci. Rep. 2018, 8, 2423. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, T.C.; Pang, R.; Yue, X.Z.; Hu, J.; Zhang, W.Q. Genome-Wide Screening and Functional Analysis Reveal That the Specific microRNA nlu-miR-173 Regulates Molting by Targeting Ftz-F1 in Nilaparvata lugens. Front. Physiol. 2018, 9, 1854. [Google Scholar] [CrossRef] [Green Version]
- Jayachandran, B.; Hussain, M.; Asgari, S. Regulation of Helicoverpa armigera ecdysone receptor by miR-14 and its potential link to baculovirus infection. J. Invertebr. Pathol. 2013, 114, 151–157. [Google Scholar] [CrossRef]
- Rahimpour, H.; Moharramipour, S.; Asgari, S.; Mehrabadi, M. The microRNA pathway core genes are differentially expressed during the development of Helicoverpa armigera and contribute in the insect’s development. Insect Biochem. Mol. Biol. 2019, 110, 121–127. [Google Scholar] [CrossRef]
- Skowronek, P.; Wójcik, Ł.; Strachecka, A. Fat Body-Multifunctional Insect Tissue. Insects 2021, 12, 547. [Google Scholar] [CrossRef]
- Zhao, X.C.; Ma, B.W.; Berg, B.G.; Xie, G.Y.; Tang, Q.B.; Guo, X.R. A global-wide search for sexual dimorphism of glomeruli in the antennal lobe of female and male Helicoverpa armigera. Sci. Rep. 2016, 6, 35204. [Google Scholar] [CrossRef]
- Kyeiccoku, G.K.; Kunimi, Y. Effect of entomopoxvirus infection of Pseudaletia separata larvae on the oviposition behavior of Cotesia kariyai. Entomol. Exp. Appl. 2010, 83, 93–97. [Google Scholar] [CrossRef]
- Langmead, B. Aligning short sequencing reads with Bowtie. Curr. Protoc. Bioinform. 2010, 32, 11–17. [Google Scholar] [CrossRef]
- Griffiths-Jones, S.; Saini, H.K.; van Dongen, S.; Enright, A.J. miRBase: Tools for microRNA genomics. Nucleic Acids Res. 2007, 36, D154–D158. [Google Scholar] [CrossRef] [Green Version]
- Friedländer, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2011, 40, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Enright, A.; John, B.; Gaul, U.; Tuschl, T.; Sander, C.; Marks, D. MicroRNA targets in Drosophila. Genome Biol. 2003, 4, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Ma, L.; Xiao, H.; Xie, B.; Liang, G. Molecular Characterization and Function Analysis of the Vitellogenin Receptor from the Cotton Bollworm, Helicoverpa armigera (Hübner) (Lepidoptera, Noctuidae). PLoS ONE 2016, 11, e0155785. [Google Scholar] [CrossRef] [Green Version]
- Singh, C.P. Role of microRNAs in insect-baculovirus interactions. Insect Biochem. Mol. Biol. 2020, 127, 103459. [Google Scholar] [CrossRef]
- Sun, X.; Liu, Q.H.; Yang, B.; Huang, J. Differential expression of microRNAs of Litopenaeus vannamei in response to different virulence WSSV infection. Fish Shellfish Immunol. 2016, 58, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yao, Y.; Zhao, Y.; Smith, L.P.; Baigent, S.J.; Nair, V. Analysis of the expression profiles of Marek’s disease virus-encoded microRNAs by real-time quantitative PCR. J. Virol. Methods 2008, 149, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhuo, H.Z.; Wu, J.Y.; Lin, L.Y.; Huang, Z.L.; Lu, J.X.; Cheng, K.L. MiR-92b inhibits proliferation and invasion of lung cancer by targeting EZH2. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3166–3173. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Zhang, Z.; Pan, L.; Zhang, Y. MicroRNA-34/449 family and viral infections. Virus Res. 2018, 260, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Avila-Bonilla, R.G.; Yocupicio-Monroy, M.; Marchat, L.A.; Pérez-Ishiwara, D.G.; Cerecedo-Mercado, D.A.; Del Ángel, R.M.; Salas-Benito, J.S. miR-927 has pro-viral effects during acute and persistent infection with dengue virus type 2 in C6/36 mosquito cells. J. Gen. Virol. 2020, 101, 825–839. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, Y.; Yan, H.; Li, Y.; Zhang, H.; Xu, J.; Puthiyakunnon, S.; Chen, X. miR-281, an abundant midgut-specific miRNA of the vector mosquito Aedes albopictus enhances dengue virus replication. Parasites Vectors 2014, 7, 488. [Google Scholar] [CrossRef]
- Zhou, H.; Wu, S.; Liu, L.; Liu, X.; Lan, S.; Jiang, J.; Yang, W.; Jin, P.; Xia, X.; Ma, F. Drosophila Relish-mediated miR-317 expression facilitates immune homeostasis restoration via inhibiting PGRP-LC. Eur. J. Immunol. 2022, 52, 1934–1945. [Google Scholar] [CrossRef]
- Liu, F.; Du, Y.; Feng, W.H. New perspective of host microRNAs in the control of PRRSV infection. Vet. Microbiol. 2017, 209, 48–56. [Google Scholar] [CrossRef]
- Khanizadeh, S.; Hasanvand, B.; Lashgarian, H.E.; Almasian, M.; Goudarzi, G. Interaction of viral oncogenic proteins with the Wnt signaling pathway. Iran. J. Basic Med. Sci. 2018, 21, 651–659. [Google Scholar] [CrossRef]
- Wang, X.; Wei, Z.; Jiang, Y.; Meng, Z.; Lu, M. mTOR Signaling: The Interface Linking Cellular Metabolism and Hepatitis B Virus Replication. Virol. Sin. 2021, 36, 1303–1314. [Google Scholar] [CrossRef]
- Jaumouillé, V.; Grinstein, S. Molecular Mechanisms of Phagosome Formation. Myeloid Cells in Health and Disease: A Synthesis 2017, 507–526. [Google Scholar] [CrossRef]
- Hoffman, R.M.; Han, Q. Oral Methioninase for Covid-19 Methionine-restriction Therapy. In Vivo 2020, 34, 1593–1596. [Google Scholar] [CrossRef]
- Feili, X.; Wu, S.; Ye, W.; Tu, J.; Lou, L. MicroRNA-34a-5p inhibits liver fibrosis by regulating TGF-β1/Smad3 pathway in hepatic stellate cells. Cell Biol. Int. 2018, 42, 1370–1376. [Google Scholar] [CrossRef]
- Li, R.; Huang, Y.; Zhang, Q.; Zhou, H.; Jin, P.; Ma, F. The miR-317 functions as a negative regulator of Toll immune response and influences Drosophila survival. Dev. Comp. Immunol. 2019, 95, 19–27. [Google Scholar] [CrossRef]
- Gomes-Duarte, A.; Lacerda, R.; Menezes, J.; Romão, L. eIF3: A factor for human health and disease. RNA Biol. 2018, 15, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Morais, A.; Terzian, A.C.; Duarte, D.V.; Bronzoni, R.V.; Madrid, M.C.; Gavioli, A.F.; Gil, L.H.; Oliveira, A.G.; Zanelli, C.F.; Valentini, S.R.; et al. The eukaryotic translation initiation factor 3 subunit L protein interacts with Flavivirus NS5 and may modulate yellow fever virus replication. Virol. J. 2013, 10, 205. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, X.; Wang, Q.; Luo, M.; Guo, H.; Gong, W.; Tu, C.; Sun, J. The eukaryotic translation initiation factor 3 subunit E binds to classical swine fever virus NS5A and facilitates viral replication. Virology 2018, 515, 11–20. [Google Scholar] [CrossRef]
- Maeda, K.; Kobayashi, Y.; Koide, M.; Uehara, S.; Okamoto, M.; Ishihara, A.; Kayama, T.; Saito, M.; Marumo, K. The Regulation of Bone Metabolism and Disorders by Wnt Signaling. Int. J. Mol. Sci. 2019, 20, 5525. [Google Scholar] [CrossRef] [Green Version]
- Bruton, R.K.; Rasti, M.; Mapp, K.L.; Young, N.; Carter, R.Z.; Abramowicz, I.A.; Sedgwick, G.G.; Onion, D.F.; Shuen, M.; Mymryk, J.S.; et al. C-terminal-binding protein interacting protein binds directly to adenovirus early region 1A through its N-terminal region and conserved region 3. Oncogene 2007, 26, 7467–7479. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Lupiani, B.; Izumiya, Y.; Reddy, S.M. Marek’s disease virus Meq oncoprotein interacts with chicken HDAC 1 and 2 and mediates their degradation via proteasome dependent pathway. Sci. Rep. 2021, 11, 637. [Google Scholar] [CrossRef]
- Ohashi, M.; Hayes, M.; McChesney, K.; Johannsen, E. Epstein-Barr virus nuclear antigen 3C (EBNA3C) interacts with the metabolism sensing C-terminal binding protein (CtBP) repressor to upregulate host genes. PLoS Pathog. 2021, 17, e1009419. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | CK_1 | CK_2 | CK_3 | MbMNPV_1 | MbMNPV_2 | MbMNPV_3 |
|---|---|---|---|---|---|---|
| Raw reads | 29.55 M | 29.17 M | 31.7 M | 27.16 M | 27.2 M | 27.34 M |
| Sequences with Q20 Greater than 80% | 23.92 M | 24.13 M | 23.32 M | 23.83 M | 23.97 M | 24.16 M |
| Clean reads | 23.88 M | 24.08 M | 23.28 M | 23.79 M | 23.93 M | 24.12 M |
| Q30 | 95.94% | 96.84% | 96.71% | 96.31% | 96.79% | 96.98% |
| GC content | 55.31% | 56.90% | 56.94% | 57.62% | 57.20% | 59.02% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Z.; Yang, Y.; Sun, X.; Du, J.; Wang, Q.; Zhang, G.; Zhang, J.; Yin, X.; Singh, D.; Su, P.; et al. Integrated Analysis of MicroRNA and mRNA Expression Profiles in the Fat Bodies of MbMNPV-Infected Helicoverpa armigera. Viruses 2023, 15, 19. https://doi.org/10.3390/v15010019
Liang Z, Yang Y, Sun X, Du J, Wang Q, Zhang G, Zhang J, Yin X, Singh D, Su P, et al. Integrated Analysis of MicroRNA and mRNA Expression Profiles in the Fat Bodies of MbMNPV-Infected Helicoverpa armigera. Viruses. 2023; 15(1):19. https://doi.org/10.3390/v15010019
Chicago/Turabian StyleLiang, Zhenpu, Yanqing Yang, Xiaoyan Sun, Junyang Du, Qiuyun Wang, Guozhi Zhang, Jiran Zhang, Xinming Yin, Deepali Singh, Ping Su, and et al. 2023. "Integrated Analysis of MicroRNA and mRNA Expression Profiles in the Fat Bodies of MbMNPV-Infected Helicoverpa armigera" Viruses 15, no. 1: 19. https://doi.org/10.3390/v15010019