
Enterococcus faecium Bacteriophage vB_EfaH_163, a New Member of the Herelleviridae Family, Reduces the Mortality Associated with an E. faecium vanR Clinical Isolate in a Galleria mellonella Animal Model
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample, Strains and Culture Conditions
2.2. Phage Titre Determination
2.3. Phage Isolation and Propagation
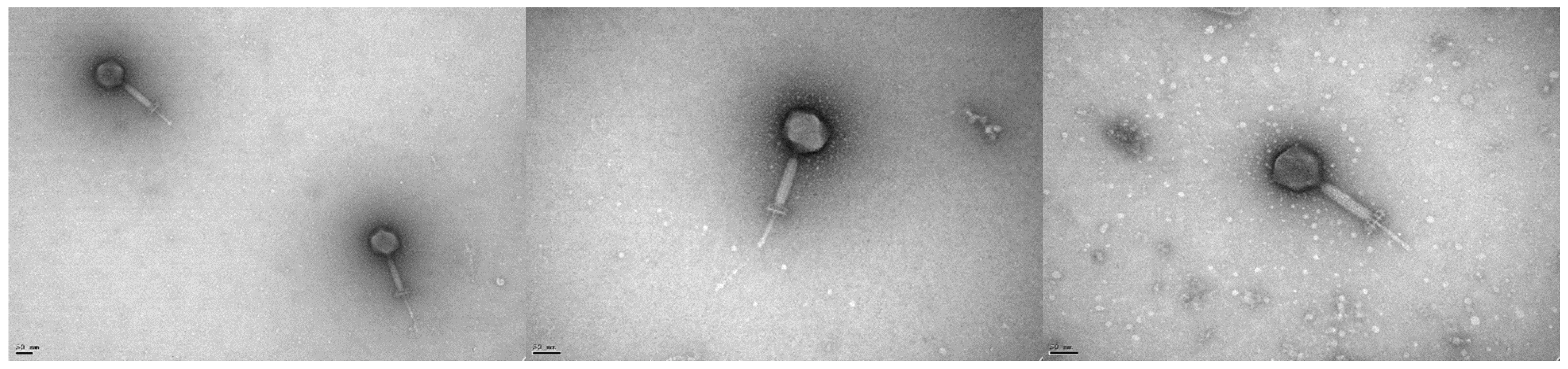
2.4. Electron Microscopy
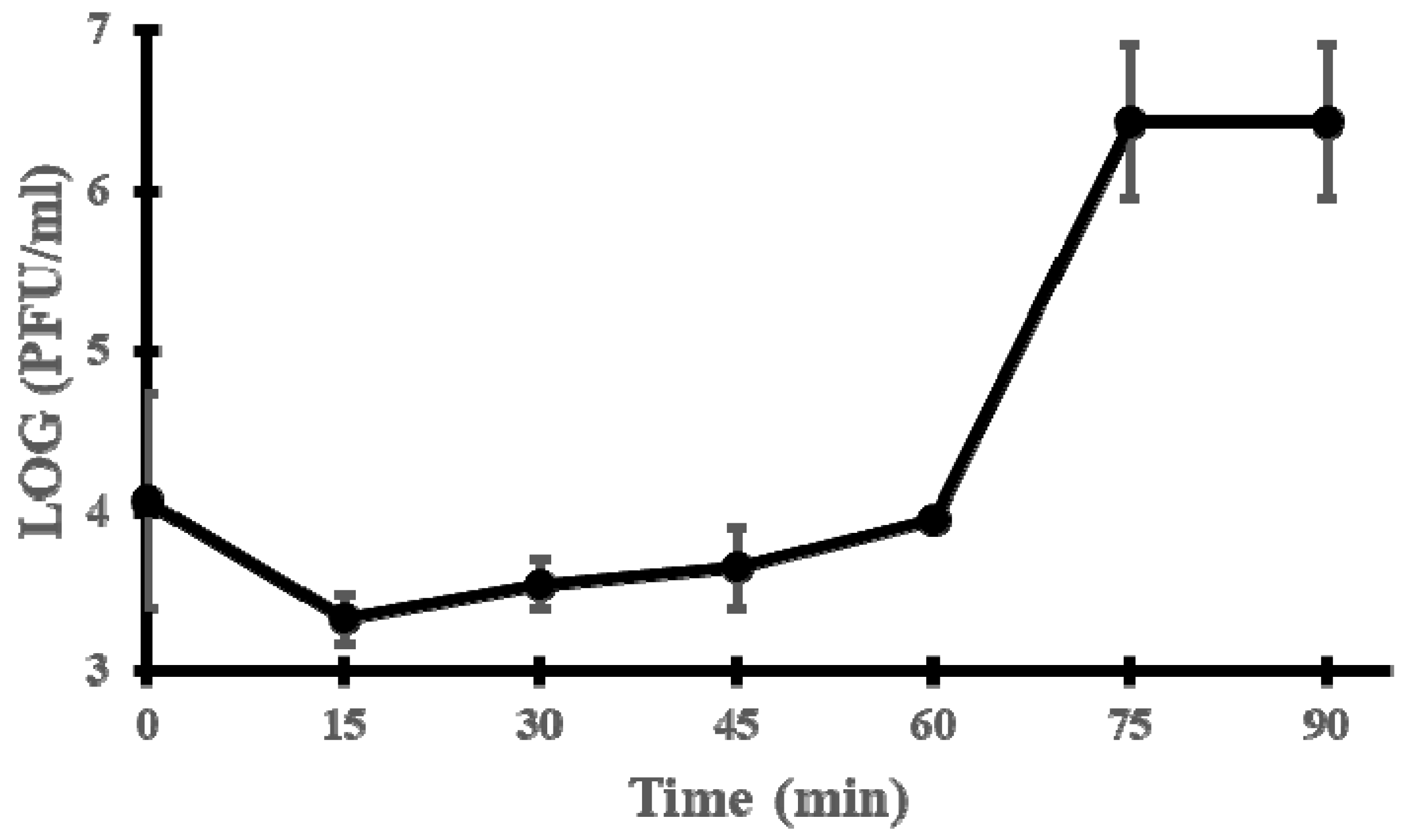
2.5. One-Step Growth Curve
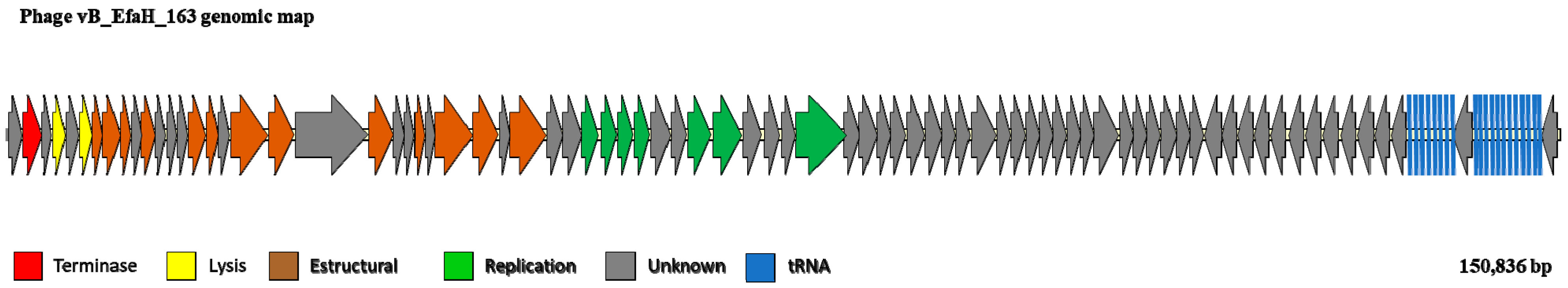
2.6. Phage Genome Sequencing and Analysis
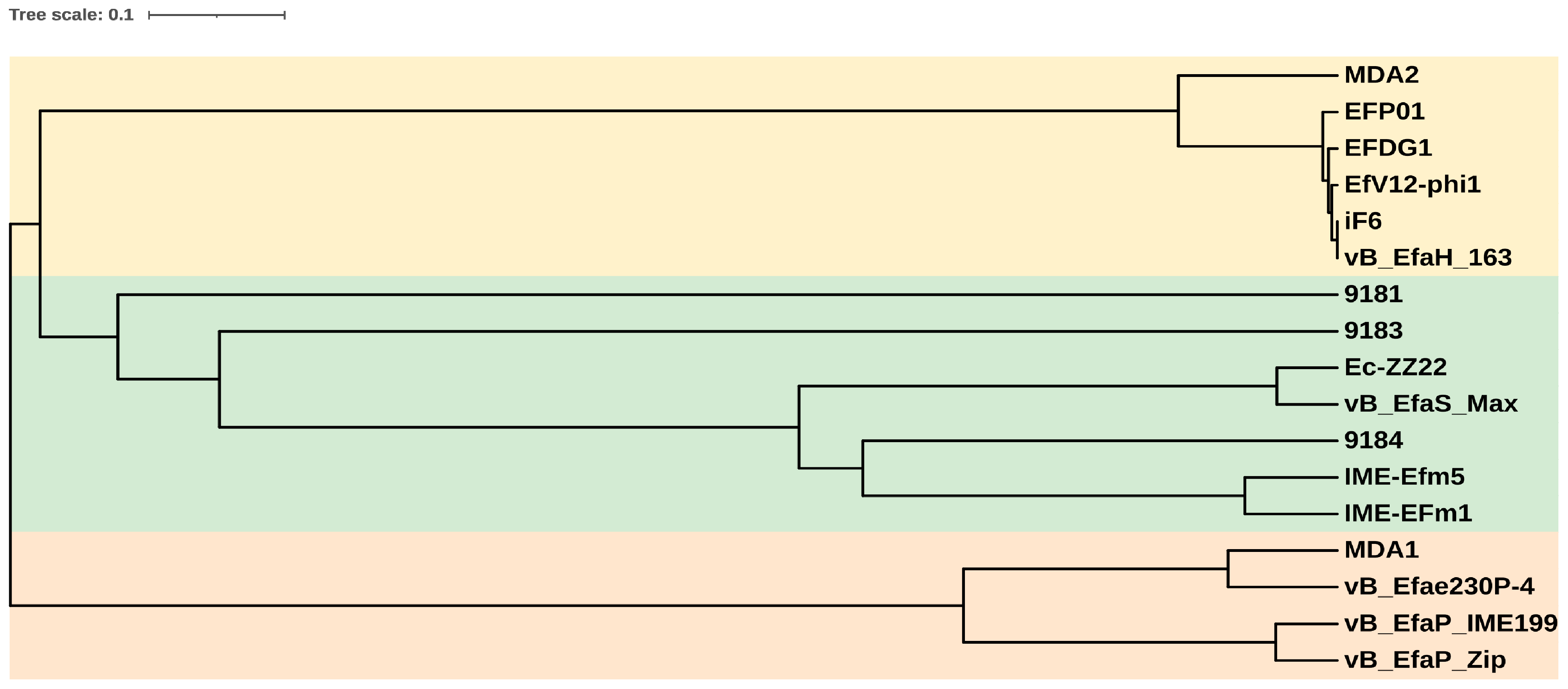
2.7. Phylogenetic Analysis
2.8. Technological Characterisation
2.9. Functional Characterisation
2.9.1. Biocontrol of the E. faecium VR-13 vanR Clinical Isolate by vB_EfaH_163 Infection in Broth
2.9.2. In Vivo Effectiveness of Phage Treatment in the Galleria mellonella Model
2.10. Statistical Analysis
3. Results
3.1. Phage vB_EfaH_163 Isolation
3.2. Microbiological Characterisation of Phage vB_EfaH_163
3.3. The vB_EfmH_163 Genome: Characterisation and Phylogenetic Analysis
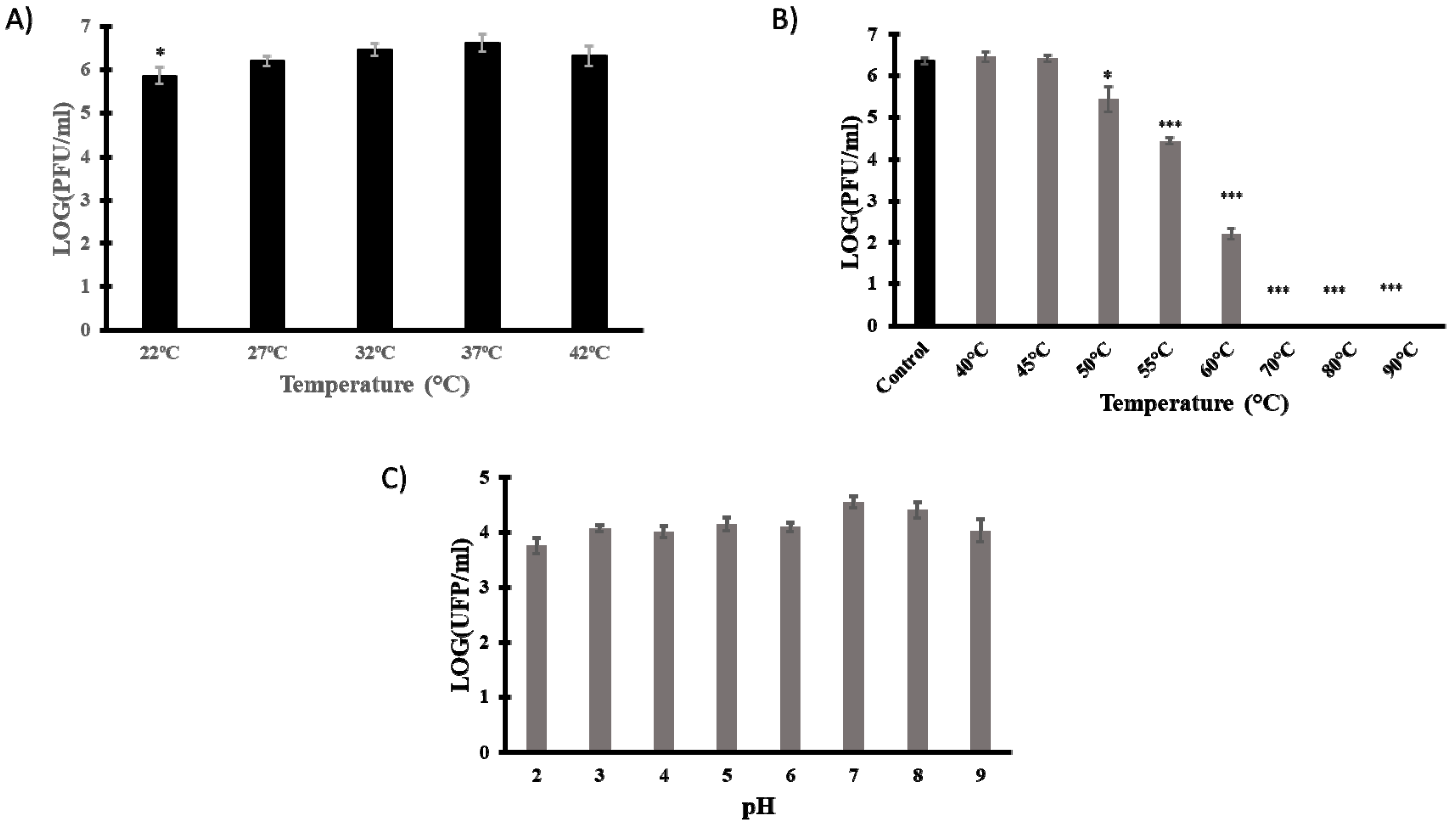
3.4. Technological Characterisation
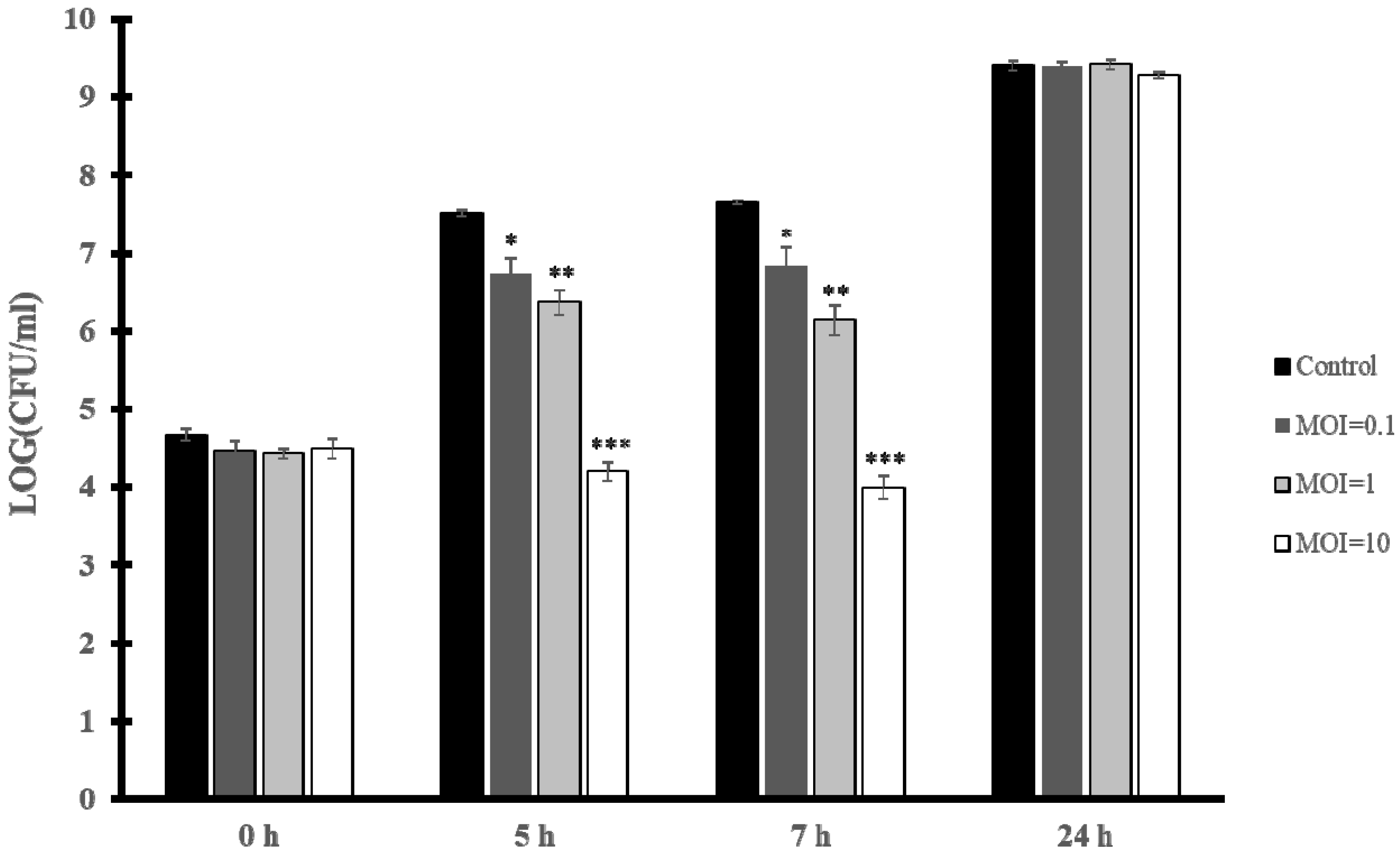
3.5. Biocontrol of E. faecium VR-13 by vB_EfaH_163
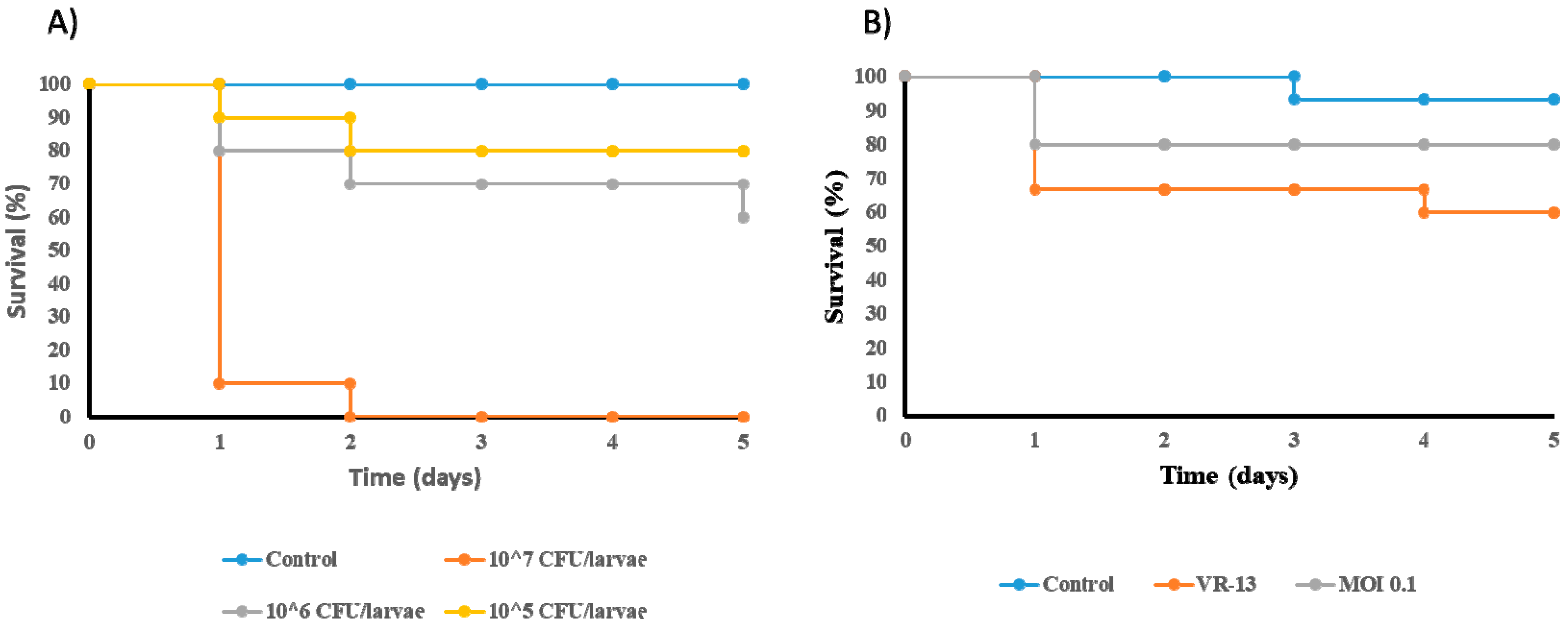
3.6. Reduction in the Mortality of Galleria mellonella Infected by E. faecium VR-13 due to Treatment with Phage vB_EfaH_163
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mestrovic, T.; Robles Aguilar, G.; Swetschinski, L.R.; Ikuta, K.S.; Gray, A.P.; Davis Weaver, N.; Han, C.; Wool, E.E.; Gershberg Hayoon, A.; Hay, S.I.; et al. The burden of bacterial antimicrobial resistance in the WHO European region in 2019: A cross-country systematic analysis. Lancet Public Health 2022, 2667, e897–e913. [Google Scholar] [CrossRef]
- Shankar, P.R. Book review: Tackling drug-resistant infections globally. Arch. Pharm. Pract. 2016, 7, 110. [Google Scholar] [CrossRef]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging strategies to combat ESKAPE pathogens in the era of antimicrobial resistance: A review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef] [PubMed]
- Melo, L.D.R.; Ferreira, R.; Costa, A.R.; Oliveira, H.; Azeredo, J. Efficacy and safety assessment of two enterococci phages in an in vitro biofilm wound model. Sci. Rep. 2019, 9, 6643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, A.; Bruun, N.E. Enterococcus faecalis infective endocarditis: Focus on clinical aspects. Expert Rev. Cardiovasc. Ther. 2013, 11, 1247–1257. [Google Scholar] [CrossRef]
- Zaheer, R.; Cook, S.R.; Barbieri, R.; Goji, N.; Cameron, A.; Petkau, A.; Polo, R.O.; Tymensen, L.; Stamm, C.; Song, J.; et al. Surveillance of Enterococcus spp. reveals distinct species and antimicrobial resistance diversity across a One-Health continuum. Sci. Rep. 2020, 10, 3937. [Google Scholar] [CrossRef] [Green Version]
- Stuart, C.; Schwartz, S.; Beeson, T.; Owatz, C. Enterococcus faecalis: Its role in root canal treatment failure and current concepts in retreatment. J. Endod. 2006, 32, 93–98. [Google Scholar] [CrossRef]
- Tebruegge, M.; Pantazidou, A.; Clifford, V.; Gonis, G.; Ritz, N.; Connell, T.; Curtis, N. The age-related risk of co-existing meningitis in children with urinary tract infection. PLoS ONE 2011, 6, e26576. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wang, Y.; Sun, L.; Wang, X. Vancomycin-resistant Enterococcus faecium pneumonia in a uremic patient on hemodialysis: A case report and review of the literature. BMC Infect. Dis. 2020, 20, 167. [Google Scholar] [CrossRef]
- Chuang, Y.-C.; Lin, H.-Y.; Chen, P.-Y.; Lin, C.-Y.; Wang, J.-T.; Chang, S.-C. Daptomycin versus linezolid for the treatment of vancomycin-resistant enterococcal bacteraemia: Implications of daptomycin dose. Clin. Microbiol. Infect. 2016, 22, 890.e1–890.e7. [Google Scholar] [CrossRef]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; Leon-Sampedro, R.; Del Campo, R.; Coque, T.M.; León-Sampedro, R.; Del Campo, R.; Coque, T.M. Antimicrobial resistance in Enterococcus spp. of animal origin. Microbiol. Spectr. 2018, 6, 185–227. [Google Scholar] [CrossRef] [PubMed]
- Palareti, G.; Legnani, C.; Cosmi, B.; Antonucci, E.; Erba, N.; Poli, D.; Testa, S.; Tosetto, A.; De Micheli, V.; Ghirarduzzi, A.; et al. Comparison between different D-Dimer cutoff values to assess the individual risk of recurrent venous thromboembolism: Analysis of results obtained in the DULCIS study. Int. J. Lab. Hematol. 2016, 38, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Lam, P.W.; Kozak, R.A.; Eshaghi, A.; Avaness, M.; Salt, N.; Patel, S.N.; Simor, A.E.; Leis, J.A. Nosocomial outbreak of vanD -carrying vancomycin-resistant Enterococcus faecium. Infect. Control Hosp. Epidemiol. 2018, 39, 1266–1268. [Google Scholar] [CrossRef] [PubMed]
- Egan, S.A.; Shore, A.C.; O’Connell, B.; Brennan, G.I.; Coleman, D.C. Linezolid resistance in Enterococcus faecium and Enterococcus faecalis from hospitalized patients in Ireland: High prevalence of the MDR genes optrA and poxtA in isolates with diverse genetic backgrounds. J. Antimicrob. Chemother. 2020, 75, 1704–1711. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Chatterjee, S.; Datta, S.; Prasad, R.; Dubey, D.; Prasad, R.K.; Vairale, M.G. Bacteriophages and its applications: An overview. Folia Microbiol. 2017, 62, 17–55. [Google Scholar] [CrossRef]
- Cisek, A.A.; Dąbrowska, I.; Gregorczyk, K.P.; Wyżewski, Z. Phage therapy in bacterial infections treatment: One hundred years after the discovery of bacteriophages. Curr. Microbiol. 2017, 74, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Elbreki, M.; Ross, R.P.; Hill, C.; O’Mahony, J.; McAuliffe, O.; Coffey, A. Bacteriophages and their derivatives as biotherapeutic agents in disease prevention and treatment. J. Viruses 2014, 2014, 382539. [Google Scholar] [CrossRef] [Green Version]
- Bolocan, A.S.; Upadrasta, A.; Bettio, P.H.A.; Clooney, A.G.; Draper, L.A.; Ross, R.P.; Hill, C. Evaluation of phage therapy in the context of Enterococcus faecalis and its associated diseases. Viruses 2019, 11, 366. [Google Scholar] [CrossRef] [Green Version]
- Fernández, L.; Gutiérrez, D.; García, P.; Rodríguez, A. The perfect bacteriophage for therapeutic applications—A quick guide. Antibiotics 2019, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, L.; Shlezinger, M.; Beyth, S.; Houri-Haddad, Y.; Coppenhagen-Glazer, S.; Beyth, N.; Hazan, R. Phage therapy against Enterococcus faecalis in dental root canals. J. Oral Microbiol. 2016, 8, 32157. [Google Scholar] [CrossRef]
- Khalifa, L.; Brosh, Y.; Gelman, D.; Coppenhagen-Glazer, S.; Beyth, S.; Poradosu-Cohen, R.; Que, Y.-A.; Beyth, N.; Hazan, R. Targeting Enterococcus faecalis biofilms with phage therapy. Appl. Environ. Microbiol. 2015, 81, 2696–2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifa, L.; Gelman, D.; Shlezinger, M.; Dessal, A.L.; Coppenhagen-Glazer, S.; Beyth, N.; Hazan, R. Defeating antibiotic- and phage-resistant Enterococcus faecalis using a phage cocktail in vitro and in a clot model. Front. Microbiol. 2018, 9, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Rio, B.; Sanchez-Llana, E.; Redruello, B.; Magadan, A.H.; Fernandez, M.; Martin, M.C.; Ladero, V.; Alvarez, M.A. Enterococcus faecalis bacteriophage 156 Is an effective biotechnological tool for reducing the presence of tyramine and putrescine in an experimental cheese model. Front. Microbiol. 2019, 10, 566. [Google Scholar] [CrossRef] [PubMed]
- Ladero, V.; Gomez-Sordo, C.; Sanchez-Llana, E.; del Rio, B.; Redruello, B.; Fernandez, M.; Martin, M.C.; Alvarez, M.A. Q69 (an E. faecalis-Infecting bacteriophage) as a biocontrol agent for reducing tyramine in dairy products. Front. Microbiol. 2016, 7, 445. [Google Scholar] [CrossRef] [PubMed]
- Biswas, B.; Adhya, S.; Washart, P.; Paul, B.; Trostel, A.N.; Powell, B.; Carlton, R.; Merril, C.R. Bacteriophage therapy rescues mice bacteremic from a clinical isolate of vancomycin-resistant Enterococcus faecium. Infect. Immun. 2002, 70, 1664. [Google Scholar] [CrossRef] [Green Version]
- Gelman, D.; Beyth, S.; Lerer, V.; Adler, K.; Poradosu-Cohen, R.; Coppenhagen-Glazer, S.; Hazan, R. Combined bacteriophages and antibiotics as an efficient therapy against VRE Enterococcus faecalis in a mouse model. Res. Microbiol. 2018, 169, 531–539. [Google Scholar] [CrossRef]
- Letkiewicz, S.; Międzybrodzki, R.; Fortuna, W.; Weber-Dąbrowska, B.; Górski, A. Eradication of Enterococcus faecalis by phage therapy in chronic bacterial prostatitis—Case report. Folia Microbiol. 2009, 54, 457–461. [Google Scholar] [CrossRef]
- Ladero, V.; Fernandez, M.; Calles-Enriquez, M.; Sanchez-Llana, E.; Canedo, E.; Martin, M.C.; Alvarez, M.A. Is the production of the biogenic amines tyramine and putrescine a species-level trait in enterococci? Food Microbiol. 2012, 30, 132–138. [Google Scholar] [CrossRef]
- Ladero, V.; Fernandez, M.; Alvarez, M.A. Isolation and identification of tyramine-producing enterococci from human fecal samples. Can. J. Microbiol. 2009, 55, 215–218. [Google Scholar] [CrossRef]
- Paulsen, I.T.; Banerjei, L.; Myers, G.S.; Nelson, K.E.; Seshadri, R.; Read, T.D.; Fouts, D.E.; Eisen, J.A.; Gill, S.R.; Heidelberg, J.F.; et al. Role of mobile DNA in the evolution of vancomycin-resistant Enterococcus faecalis. Science 2003, 299, 2071–2074. [Google Scholar] [CrossRef]
- Hevia, A.; Delgado, S.; Margolles, A.; Sánchez, B. Application of density gradient for the isolation of the fecal microbial stool component and the potential use thereof. Sci. Rep. 2015, 5, 16807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binetti, A.G.; del Rio, B.; Martin, M.C.; Alvarez, M.A. Detection and characterization of Streptococcus thermophilus bacteriophages by use of the antireceptor gene sequence. Appl. Environ. Microbiol. 2005, 71, 6096–6103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Rio, B.; Sánchez-Llana, E.; Martínez, N.; Fernández, M.; Ladero, V.; Alvarez, M.A. Isolation and characterization of Enterococcus faecalis-infecting bacteriophages from different cheese types. Front. Microbiol. 2021, 11, 592172. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial bioinformatics database and analysis resource center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Yukgehnaish, K.; Rajandas, H.; Parimannan, S.; Manickam, R.; Marimuthu, K.; Petersen, B.; Clokie, M.R.J.; Millard, A.; Sicheritz-Pontén, T. PhageLeads: Rapid assessment of phage therapeutic suitability using an ensemble machine learning approach. Viruses 2022, 14, 342. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2019, 48, D517–D525. [Google Scholar] [CrossRef]
- Garneau, J.R.; Depardieu, F.; Fortier, L.-C.; Bikard, D.; Monot, M. PhageTerm: A tool for fast and accurate determination of phage termini and packaging mechanism using next-generation sequencing data. Sci. Rep. 2017, 7, 8292. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Br. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Wandro, S.; Oliver, A.; Gallagher, T.; Weihe, C.; England, W.; Martiny, J.B.H.; Whiteson, K. Predictable molecular adaptation of coevolving Enterococcus faecium and lytic phage EfV12-phi1. Front. Microbiol. 2019, 9, 3192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Haddad, L.; Angelidakis, G.; Clark, J.R.; Mendoza, J.F.; Terwilliger, A.L.; Chaftari, C.P.; Duna, M.; Yusuf, S.T.; Harb, C.P.; Stibich, M.; et al. Genomic and functional characterization of vancomycin-resistant enterococci-specific bacteriophages in the Galleria mellonella wax moth larvae model. Pharmaceutics 2022, 14, 1591. [Google Scholar] [CrossRef]
- Canfield, G.S.; Chatterjee, A.; Espinosa, J.; Mangalea, M.R.; Sheriff, E.K.; Keidan, M.; McBride, S.W.; McCollister, B.D.; Hang, H.C.; Duerkop, B.A. Lytic bacteriophages facilitate antibiotic sensitization of Enterococcus faecium. Antimicrob. Agents Chemother. 2021, 65, e00143-21. [Google Scholar] [CrossRef]
- Li, J.; Shi, H.; Zhao, C.; Hao, Y.; He, Y.; Sun, Y. Complete genome sequence of the siphoviral bacteriophage ec-zz2, which is capable of lysing Enterococcus faecium. Genome Announc. 2016, 4, e01167-16. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, W.; Lv, Y.; Zheng, W.; Mi, Z.; Pei, G.; An, X.; Xu, X.; Han, C.; Liu, J.; et al. Characterization and complete genome sequence analysis of novel bacteriophage IME-EFm1 infecting Enterococcus faecium. J. Gen. Virol. 2014, 95, 2565–2575. [Google Scholar] [CrossRef] [Green Version]
- Gong, P.; Cheng, M.; Li, X.; Jiang, H.; Yu, C.; Kahaer, N.; Li, J.; Zhang, L.; Xia, F.; Hu, L.; et al. Characterization of Enterococcus faecium bacteriophage IME-EFm5 and its endolysin LysEFm5. Virology 2016, 492, 11–20. [Google Scholar] [CrossRef]
- Xing, S.; Zhang, X.; Sun, Q.; Wang, J.; Mi, Z.; Pei, G.; Huang, Y.; An, X.; Fu, K.; Zhou, L.; et al. Complete genome sequence of a novel, virulent Ahjdlikevirus bacteriophage that infects Enterococcus faecium. Arch. Virol. 2017, 162, 3843–3847. [Google Scholar] [CrossRef]
- Manohar, P.; Nachimuthu, R.; Lopes, B.S. The therapeutic potential of bacteriophages targeting gram-negative bacteria using Galleria mellonella infection model. BMC Microbiol. 2018, 18, 97. [Google Scholar] [CrossRef]
- Jeon, J.; Park, J.-H.; Yong, D. Efficacy of bacteriophage treatment against carbapenem-resistant Acinetobacter baumannii in Galleria mellonella larvae and a mouse model of acute pneumonia. BMC Microbiol. 2019, 19, 70. [Google Scholar] [CrossRef] [PubMed]
- Lagatolla, C.; Milic, J.; Imperi, F.; Cervoni, M.; Bressan, R.; Luzzati, R.; Di Bella, S. Synergistic activity of fosfomycin and chloramphenicol against vancomycin-resistant Enterococcus faecium (VREfm) isolates from bloodstream infections. Diagn. Microbiol. Infect. Dis. 2021, 99, 115241. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, H.; Morita, M. Phage DNA packaging. Genes Cells 2003, 2, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Barylski, J.; Kropinski, A.M.; Alikhan, N.-F.; Adriaenssens, E.M. ICTV report consortium ICTV virus taxonomy profile: Herelleviridae. J. Gen. Virol. 2020, 101, 362. [Google Scholar] [CrossRef]
- Turner, D.; Adriaenssens, E.M.; Tolstoy, I.; Kropinski, A.M. Phage annotation guide: Guidelines for assembly and high-quality annotation. PHAGE 2021, 2, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Chibani-Chennoufi, S.; Bruttin, A.; Dillmann, M.; Brüssow, H. Phage-host interaction: An ecological perspective. J. Bacteriol. 2004, 186, 3677–3686. [Google Scholar] [CrossRef] [Green Version]
- Kot, W.; Neve, H.; Heller, K.J.; Vogensen, F.K. Bacteriophages of leuconostoc, oenococcus, and weissella. Front. Microbiol. 2014, 5, 186. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, J.; Rashel, M.; Maeda, Y.; Takemura, I.; Sugihara, S.; Akechi, K.; Muraoka, A.; Wakiguchi, H.; Matsuzaki, S. Isolation and characterization of a novel Enterococcus faecalis bacteriophage φEF24C as a therapeutic candidate. FEMS Microbiol. Lett. 2008, 278, 200–206. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Fischer, D. Identification and investigation of ORFans in the viral world. BMC Genom. 2008, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Rubio, L.; Gutiérrez, D.; Donovan, D.M.; Martínez, B.; Rodríguez, A.; García, P. Phage lytic proteins: Biotechnological applications beyond clinical antimicrobials. Crit. Rev. Biotechnol. 2015, 36, 542–552. [Google Scholar] [CrossRef]
- Wang, R.; Xing, S.; Zhao, F.; Li, P.; Mi, Z.; Shi, T.; Liu, H.; Tong, Y. Characterization and genome analysis of novel phage vB_EfaP_IME195 infecting Enterococcus faecalis. Virus Genes 2018, 54, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Casey, E.; van Sinderen, D.; Mahony, J. In vitro characteristics of phages to guide ‘Real Life’ phage therapy suitability. Viruses 2018, 10, 163. [Google Scholar] [CrossRef] [Green Version]
- Ladero, V.; Calles-Enríquez, M.; Fernández, M.; Alvarez, M.A. Toxicological effects of dietary biogenic amines. Curr. Nutr. Food Sci. 2010, 6, 145–156. [Google Scholar] [CrossRef]
- McLaughlin, M.; Malczynski, M.; Qi, C.; Barajas, G.; Radetski, J.; Zembower, T.; Scheetz, M.H. Virulence of Vancomycin-resistant Enterococcus faecium according to linezolid resistance and clinical outbreak status. Antimicrob. Agents Chemother. 2013, 57, 3923–3927. [Google Scholar] [CrossRef] [Green Version]
- Skinner, K.; Sandoe, J.A.T.; Rajendran, R.; Ramage, G.; Lang, S. Efficacy of rifampicin combination therapy for the treatment of enterococcal infections assessed in vivo using a Galleria mellonella infection model. Int. J. Antimicrob. Agents 2017, 49, 507–511. [Google Scholar] [CrossRef] [Green Version]
- Tkhilaishvili, T.; Wang, L.; Tavanti, A.; Trampuz, A.; Di Luca, M. Antibacterial efficacy of two commercially available bacteriophage formulations, staphylococcal bacteriophage and pyo bacteriophage, against methicillin-resistant Staphylococcus aureus: Prevention and eradication of biofilm formation and control of a systemic infection of Galleria mellonella larvae. Front. Microbiol. 2020, 11, 110. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specie | Strain | Origin | vB_EfmH_163 Infection | Reference |
|---|---|---|---|---|
| E. faecium | LMA2 | Camel milk | + | MicroMol |
| E. faecium | LMA3 | Camel milk | + | MicroMol |
| E. faecium | LMA4 | Camel milk | + | MicroMol |
| E. faecium | LMA5 | Camel milk | + | MicroMol |
| E. faecium | LMA6 | Camel milk | + | MicroMol |
| E. faecium | LMA8 | Camel milk | + | MicroMol |
| E. faecium | LMA9 | Camel milk | − | MicroMol |
| E. faecium | LMA10 | Camel milk | + | MicroMol |
| E. faecium | LGMY-2 | Camel milk | + | MicroMol |
| E. faecium | LGMY-5 | Camel milk | + | MicroMol |
| E. faecium | LGMY-1 | Cow milk | + | MicroMol |
| E. faecium | LMGY-10 | Cow milk | − | MicroMol |
| E. faecium | LGMY-6 | Sheep milk | + | MicroMol |
| E. faecium | LGMY-11 | Goat milk | + | MicroMol |
| E. faecium | C39 | Cheese | − | [28] |
| E. faecium | AM | Cheese | − | [28] |
| E. faecium | 103 | Cheese | − | [28] |
| E. faecium | LGMY-12 | Date | + | MicroMol |
| E. faecium | LMGY-13 | Date | + | LGM |
| E. faecium | LGM11397 | Meat | − | LGM |
| E. faecium | LGM14205 | Meat | − | LGM |
| E. faecium | LGM20641 | Meat | − | LGM |
| E. faecium | HF11 | Human | − | [29] |
| E. faecium | HF14 | Human | + | [29] |
| E. faecium | HF24 | Human | + | [29] |
| E. faecium | HF52 | Human | − | [29] |
| E. faecium | HF56 | Human | + | [29] |
| E. faecium | VR-1 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-2 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-3 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-4 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-5 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-6 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-7 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-8 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-9 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-10 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-11 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-12 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-13 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-14 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-15 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-16 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-17 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-18 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-19 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-20 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-20b | Clinical | + | Bierzo Hospital |
| E. faecium | VR-22 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-23 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-24 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-25 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-26 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-27 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-28 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-29 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-30 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-31 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-32 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-33 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-34 | Clinical | + | Bierzo Hospital |
| E. faecium | VR-35 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-36 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-37 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-38 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-39 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-40 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-41 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-42 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-43 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-44 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-45 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-46 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-47 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-48 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-49 | Clinical | − | Bierzo Hospital |
| E. faecium | VR-50 | Clinical | − | Bierzo Hospital |
| E. faecalis | CECT481T | Type strain | − | CECT |
| E. faecalis | 18a | Cheese | − | [28] |
| E. faecalis | 23a | Cheese | − | [28] |
| E. faecalis | V63 | Cheese | − | [28] |
| E. faecalis | 63c | Cheese | − | [28] |
| E. faecalis | 52c | Cheese | − | [28] |
| E. faecalis | HFS56 | Human | + | [29] |
| E. faecalis | HFS57 | Human | − | [29] |
| E. faecalis | VR-5 | Clinical | − | Bierzo Hospital |
| E. faecalis | VR-11 | Clinical | − | Bierzo Hospital |
| E. faecalis | optra5 | Clinical | + | Bierzo Hospital |
| E. faecalis | V583 | Clinical | − | [30] |
| Phage | Accession Number | Host | Family | Genome Size | Origin | Reference |
|---|---|---|---|---|---|---|
| vB_EfmH_163 | CAJDKA010000002.1 | E. faecium E. faecalis | Herelleviridae | 150,836 | Human faecal samples | This work |
| EFDG1 | NC_029009 | E. faecium E faecalis | Herelleviridae | 147,589 | Sewage effluents | [21] |
| EfV12-phi1 | MH880817 | E. faecium | Herelleviridae | 152,770 | Sewage | [43] |
| EFP01 | NC_047796.1 | E. faecium | Herelleviridae | 155,053 | Sewage | - |
| iF6 | MT909815.1 | E. faecium | Herelleviridae | 156,592 | - | - |
| MDA2 | MW633168.1 | E. faecium | Herelleviridae | 140,226 | - | [44] |
| 9183 | MT939241.1 | E. faecium | Siphoviridae | 806,301 | Wastewater | [45] |
| 9181 | MT939240.1 | E. faecium | Siphoviridae | 71,854 | Wastewater | [45] |
| vB_EfaS_Max | MK360024 | E. faecium E. faecalis | Siphoviridae | 40,975 | Raw sewage from wastewater | [4] |
| 9184 | MT939242.1 | E. faecium | Siphoviridae | 44,108 | Wastewater | [45] |
| Ec-ZZ2 | NC_031260 | E. faecium | Siphoviridae | 41,170 | Sewage | [46] |
| IME-EFm1 | NC_024356 | E. faecium | Siphoviridae | 42,597 | Hospital sewage | [47] |
| IME-EFm5 | NC_028826 | E. faecium | Siphoviridae | 42,265 | Hospital sewage | [48] |
| vB_EfaP_Zip | MK360025 | E. faecium E. faecalis | Podoviridae | 18,742 | Raw sewage from wastewater | [4] |
| vB_Efae230p-4 | NC_025467 | E. faecium | Podoviridae | 17,972 | - | - |
| vB_EfaP_IME199 | KT945995 | E. faecium | Podoviridae | 18,838 | Sewage | [49] |
| MDA1 | MW623430.1 | E. faecium | Podoviridae | 18,058 | - | [44] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pradal, I.; Casado, A.; del Rio, B.; Rodriguez-Lucas, C.; Fernandez, M.; Alvarez, M.A.; Ladero, V. Enterococcus faecium Bacteriophage vB_EfaH_163, a New Member of the Herelleviridae Family, Reduces the Mortality Associated with an E. faecium vanR Clinical Isolate in a Galleria mellonella Animal Model. Viruses 2023, 15, 179. https://doi.org/10.3390/v15010179
Pradal I, Casado A, del Rio B, Rodriguez-Lucas C, Fernandez M, Alvarez MA, Ladero V. Enterococcus faecium Bacteriophage vB_EfaH_163, a New Member of the Herelleviridae Family, Reduces the Mortality Associated with an E. faecium vanR Clinical Isolate in a Galleria mellonella Animal Model. Viruses. 2023; 15(1):179. https://doi.org/10.3390/v15010179
Chicago/Turabian StylePradal, Inés, Angel Casado, Beatriz del Rio, Carlos Rodriguez-Lucas, Maria Fernandez, Miguel A. Alvarez, and Victor Ladero. 2023. "Enterococcus faecium Bacteriophage vB_EfaH_163, a New Member of the Herelleviridae Family, Reduces the Mortality Associated with an E. faecium vanR Clinical Isolate in a Galleria mellonella Animal Model" Viruses 15, no. 1: 179. https://doi.org/10.3390/v15010179